
{kind=link}
{kind=link}
{kind=link}
{kind=link}
科尔沁沙地沙漠化恢复过程中冷蒿种群的扩散对策研究
引用本文
孙英杰, 李衍青, 赵爱芬, 黄玉清, 周爱萍, 罗艾滢, 徐广平, 姚月锋. 科尔沁沙地沙漠化恢复过程中冷蒿种群的扩散对策研究. 23(1): 3-09
SUN Ying-jie, LI Yan-qing, ZHAO Ai-fen, HUANG Yu-qing, ZHOU Ai-ping, LUO Ai-ying, XU Guang-ping, YAO Yue-feng. Research on the spread strategy ofArtemisia frigida populations during the desertification recovery process in Horqin sandy land . Acta Prataculturae Sinica, 23(1): 3-09 
Permissions
SUN Ying-jie, LI Yan-qing, ZHAO Ai-fen, HUANG Yu-qing, ZHOU Ai-ping, LUO Ai-ying, XU Guang-ping, YAO Yue-feng. Research on the spread strategy of
Copyright©2014, 草业学报编辑部
本文属于开放获取期刊。
科尔沁沙地沙漠化恢复过程中冷蒿种群的扩散对策研究
作者简介:孙英杰(1984-),女,山东济宁人,研习员,硕士。E-mail:syj525123@163.com
摘要
在科尔沁沙地冷蒿种群有2种明显形态差异的个体即直立型和匍匐型2种生长表现型植株。为深刻理解冷蒿种群在沙漠化植被恢复过程中的生态学意义,对不同生长型个体的地上和地下部分的形态特征和繁殖特性,及冷蒿种群的扩散现况进行了初步研究。结果表明,2种表现型冷蒿在枝高、枝条分化、花序数、地下根系等个体形态上均表现出差异性;直立型冷蒿地上部分的高度及平均单株地上生物总量均优于匍匐型冷蒿,匍匐型冷蒿生物量主要集中于靠近基部的0~10 cm处;通过对匍匐型冷蒿的克隆能力调查发现,被沙埋后大多数冷蒿产生分株数在10株以下,分株扩散距离主要在10~20 cm以内;冷蒿枝条被沙埋后其生根数量及长度与当季的降雨量有极大关系,而枝条被沙埋后产生分株对冷蒿种群扩散起着重要的作用;直立型冷蒿主要以种子方式进行繁殖,且种群扩散方向与当地风向有关。
关键词:
科尔沁沙地; 冷蒿; 个体形态; 沙埋; 扩散对策
中图分类号:S812.8
文献标志码:A
文章编号:1004-5759(2014)01-0003-09
Research on the spread strategy ofArtemisia frigida populations during the desertification recovery process in Horqin sandy land
Abstract
The erect and the creeping types of
Keyword:
Horqin sandy land; Artemisia frigida ; individual morphology; sand burial; diffusion strategy
引言
科尔沁沙地属于我国半干旱半湿润的农牧交错带脆弱区域,由于人类的过度放牧、开垦和樵采等,使得该地区成为我国沙漠化比较严重的地区之一。近年来,随着退耕还林还草等一系列防治工程的实施,科尔沁沙地生态环境整体上呈现出逐渐恢复的趋势。在退化生态系统恢复过程中,植被恢复是退化生态系统重建的重要部分,因此,对沙地沙漠化恢复过程中植物的适应生理生态方面研究显得尤为重要。
冷蒿( Artemisia frigida)属菊科小灌木, 广泛分布于半干旱草原和沙地[ 1, 2]。是科尔沁沙地耐性较强的一种重要固沙植物种,它具有较强的补偿性生长能力,其数量及繁殖方式随着演替退化的各个阶段变化而变化, 这一特性对植物群落的结构与功能具有重要影响[ 3]。因此,冷蒿作为沙地植物群落演替后期的优势种及草原指示种[ 4],在科尔沁沙地退化生态系统恢复过程中尤其是群落演替后期发挥着重要的作用。冷蒿具有旺盛的营养繁殖能力和一定的有性繁殖能力。沙埋、动物踩食、刈割等因素都会对其繁殖特性产生影响[ 5, 6, 7, 8]。
国内外对冷蒿进行了大量的研究,这些研究主要关注于冷蒿个体形态[ 9]、冷蒿的形态学特性[ 10]、饲用价值、生态型、根系分布等生态学特性[ 11],冷蒿挥发性油的分离和鉴定[ 12], 冷蒿的抗性生理与其他生理特性[ 13, 14, 15, 16],冷蒿繁殖对放牧的响应[ 5, 6, 17, 18]、生物量资源分配[ 7, 19, 20],冷蒿草原土壤种子库[ 21]、冷蒿群落特性[ 22, 23, 24]等方面。而关于科尔沁沙地冷蒿种群扩散现象的研究少见报道。笔者在研究科尔沁沙地植被恢复的野外调查时发现,冷蒿有2种差异明显的生长表现型,分别为枝条密集、木质化程度高且直立生长的冷蒿和枝条较柔软且匍匐生长的冷蒿。本研究分析了这2种生长型不同的个体地上与地下部分的形态特征及繁殖特点,进而了解直立型冷蒿和匍匐型冷蒿在种群扩散上的地位及作用。旨在期望能为科尔沁沙地的植被恢复和土地荒漠化治理中植物筛选方面提供理论依据或可行性措施。
1 材料与方法
1.1 研究区概况
研究样地位于内蒙古奈曼旗境内(120°55' E,42°41' N),平均海拔360 m。该区域属温带半干旱大陆性气候,干旱且多大风[ 25],年均气温6.5℃,1月份平均气温-12.7℃,7月份平均气温23.7℃,年均降水量362.3 mm,主要集中在6,7,8月,年均蒸发量1 935.4 mm,雨热同季。8级以上大风年均21次,年均风速达3.2~4.0 m/s,其中10~15 d沙尘暴天气主要出现在春季。
植被种类主要以灌木、半灌木为主,由菊科、禾本科、豆科、藜科、蒺藜科、蓼科植物组成。近百年来由于人为的干扰,使该区的原生植被受到破坏,沙漠化日益严重,植被中的乔木层已基本消失,草本层退化,灌木层发育强烈。目前植被以灌木、多年生和一年生草本植物组成的沙生植物为主,主要植物种包括冷蒿、小叶锦鸡儿( Caragana microphylla)、杠柳( Periploca sepium)、黄柳( Salix gordejevii)、差巴嘎蒿( Artemisia halodendron)、黄蒿( A. scoparia)、猪毛菜( Salsola collina)、糙隐子草( Cleistogenes squarrosa)、狗尾草( Setaria viridis)等。该区域土壤贫瘠,主要为风沙土、沙质栗钙土、沙质草甸土和沼泽土[ 26]。
1.2 研究方法
1.2.1 样地选择与实验设计
1) 调查样地:中国科学院奈曼沙漠化研究站1994年建立的禁牧的放牧围栏(样地A)、与放牧围栏相邻的东南方向的固定沙丘(样地B)。
样地A处冷蒿种群基本为直立生长,个体界限清楚,在封育围栏内成斑块状分布,约200株左右,主要集中生长在相对低洼的区域。斑块中心少有其他植物生长而靠近斑块边缘则有较多的地锦( Euphorbia humifusa)、糙隐子草、灰绿藜( Chenopodium glaucum)等植物分布。
样地B处于典型恢复过程中的半固定沙丘。土壤有风蚀,地表形成浅风蚀坑。属于冷蒿+杠柳群落。该样地与玉米地、人工杨树林、放牧围栏毗邻;土壤为沙土质土壤;小半灌木为冷蒿、差巴嘎蒿;草本植物以黄蒿、雾冰藜( Bassia dasyphylla)、灰绿藜为主;以及灌木植物——杠柳。冷蒿为匍匐生长,呈现大面积斑块状分布,从群落中心到群落边缘,冷蒿密度逐渐变小并呈现向外分散生长的态势。
2) 野外沙埋试验地:微气象站东面的固定沙丘(样地C)。此区域冷蒿植株由微气象站的固定沙丘顶部的冷蒿种群扩散而来。该样地内冷蒿为优势植物种,差巴嘎蒿逐渐退出仅余少量几株。此处冷蒿个体以直立型和匍匐型2种表现型生长,且生长旺盛,种群密度较高。
1.2.2 调查时间与方法
1) 野外调查: 2009年6月29日,在样地A处的冷蒿斑块内,以种群中最原始的冷蒿植株(调查时植株已经死亡,但根系仍在土壤中,大部分枯枝完好保留)为原点,向该株冷蒿周围种群边缘的10个冷蒿植株做样线,用经纬仪测量每条样线间的角度,并记录下每条样线上冷蒿个体在样线上的投影长度,以代表其盖度[ 27]。
2009年6月30日,在样地B中选择30株匍匐型冷蒿作为研究对象。调查每株冷蒿产生的分株的数量和分株与基株间的距离。
2009年7月24日,在样地B处,选择30株直立型冷蒿和30株匍匐型冷蒿标记,记录每株冷蒿样株的平均高度、最高枝高度、最低枝高度、丛辐、营养枝数量、生殖枝数量及花序数。然后,分别将标记的冷蒿的地上部分从茎基部剪掉,装入袋中标记后带回实验室。用分层法[ 20, 28, 29]从冷蒿茎的底端向最顶端将植株分为0~10 cm、10~20 cm、20~30 cm、30~40 cm、40~50 cm、50 cm以上6层,把每层生物量放入编号信封中,然后放入80℃的烘箱中烘24 h,24 h后取出称量,记录下每株冷蒿的各层生物量干重。
选择典型的直立型冷蒿和匍匐型冷蒿个体,将其地下部分小心挖出,去掉根系部分的沙土和地上部分,用扫描仪扫描2株冷蒿的根系。
2) 野外沙埋实验:2009年分别在6和7月选择22株352个枝条做沙埋处理实验,以确定不同沙埋时间对冷蒿生长和分株的影响。6月2日,在样地C中选择自然状态下生长良好的直立型冷蒿和匍匐型冷蒿各4株作为实验对象。从每株冷蒿中各选取6枝健康且生长态势一致的枝条做沙埋处理。7月2日,试验地点与6月2日相同,方法类似,在样地中选择生长良好的直立型和匍匐型冷蒿各7株,每株沙埋处理10个枝条。
2009年8月22日用小铁锹挖出2次处理的所有枝条,从枝条基部与母株分离并小心去掉枝条上的沙土标记装袋。小心去掉枝条附着的沙土及其他植物的细根。分别测量出每次沙埋处理的每个枝条生根枝条数,不定根的数量,不定根的长度。
1.3 数据分析与处理
1.3.1 封育围栏内冷蒿种群扩散图的绘制
1994年在因过度放牧引起的沙漠化区域进行了围栏封育,样地本底调查时冷蒿种群按照Drude多度等级应为“个别种”(Un),1998年调查时即形成冷蒿小斑块,所发现植株均为直立型生长。种群扩散调查图以样地内最大的1株冷蒿为中心,向各个方向辐射,每条线间隔30°,图上以黑点代表冷蒿植株在样线上的位置,2009年则以1998年调查地内最原始的冷蒿植株O为中心,根据经纬仪测量出来的各个样线的角度画图,图上每小格代表实际距离1 m。
冷蒿丛辐=各样线上冷蒿的投影总长度(m)/冷蒿株数
1.3.2 匍匐型冷蒿调查及2次沙埋处理枝条的处理
利用Microsoft Excel 2003对30株匍匐型冷蒿分株调查结果进行统计分析,并计算出每株冷蒿母株产生的平均分株数量,以及其分株扩散的平均最远和最近距离。
2 结果与分析
2.1 冷蒿个体地上部分的形态及生物量特征
直立型冷蒿与匍匐型冷蒿的地上形态差异主要表现在个体高度、枝条数、丛辐等方面( 表1)。从总体上来看,直立型冷蒿的高度、枝条数、每株花序数均大于匍匐型冷蒿。但直立型冷蒿的平均丛辐(61.1 cm×51.0 cm)小于匍匐型冷蒿(63.5 cm×57.8 cm),两者之间无较大差异。由于调查样地冷蒿种群大小的限制,30株冷蒿个体间生长差异的变异性较大。
| 表1 2种表现型冷蒿个体地上生长形态比较 Table 1 The comparison of growth form aboveground of two phenotypes of A. frigida |
由 t-检验结果可知,直立型和匍匐型冷蒿的最低枝高度、平均枝高、营养枝条数和平均每株每枝的花序数均存在显著差异( P<0.05),直立型冷蒿植株的生殖枝一般多于营养枝。但最高枝高度及生殖枝条数均无显著差异。虽然花序数与种子产量有很大关系,但种子形成又受到天气条件、传粉方式等因素的影响,所以花序数不能用于预测冷蒿的种子产量。
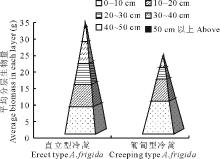
通过对冷蒿个体调查结果分析可知枝条密集生长的直立型有较高的地上生物量,直立型冷蒿植株有76.67%的单株生物量大于20 g,而匍匐型冷蒿中有50%。从构建的生物量金字塔( 图1)可以看出,在地上部分的高度及平均单株地上生物总量上,直立型冷蒿均优于匍匐型冷蒿。在0~10 cm即靠近植株基部,匍匐型冷蒿的生物量大于直立型冷蒿,而且基部的生物量大于10~50 cm以上部分的任一层。直立型冷蒿10~50 cm以上这5层的生物量大于匍匐型冷蒿,其株高达50 cm以上,但匍匐型冷蒿却在50 cm以内。以上结果可能与冷蒿的枝条木质化及匍匐型冷蒿生长得较低矮直接相关。
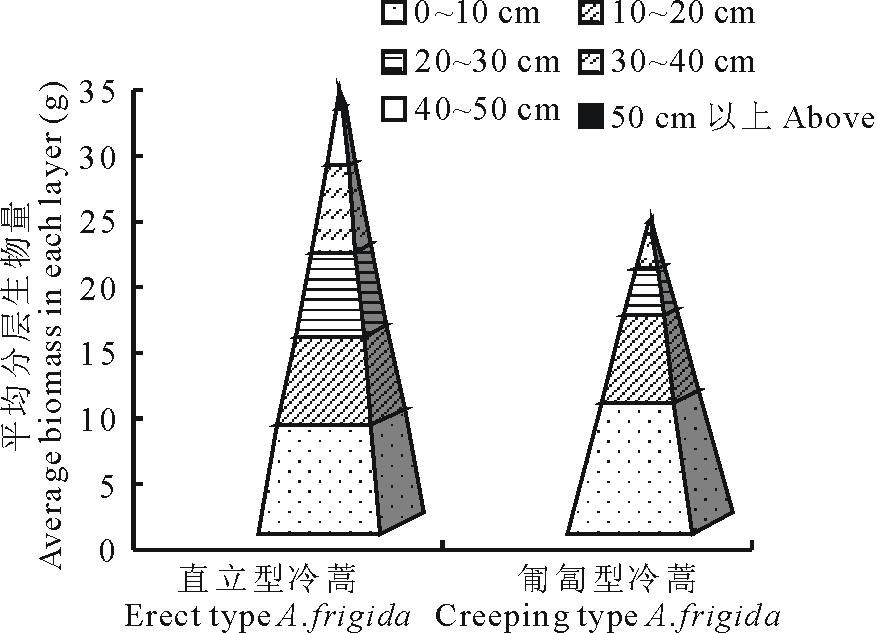 | 图1 2种表现型冷蒿的地上生物量比较Fig.1 The comparison of aboveground biomass of two phenotypes of A. frigida |
2.2 冷蒿地下根系形态生长特性
调查发现直立型冷蒿是由种子萌发的有性生殖产生,因此植株的根系类似于直根系( 图2左图),这种形态有利于冷蒿植株从更深的土壤层次中吸取水分以满足整个植物的生长发育所需。而匍匐型冷蒿的根系都有与其他植株相连的老枝条,形态如 图2右图所示,o点为冷蒿基株的基部,a~e分别为基株产生的分株处。随着时间的推移,各分株与基株脱离形成一个新的冷蒿植株个体。冷蒿枝条被沙埋后,在适宜环境下会生长出不定根进而在基株上形成不同大小的分株,从而形成了丛辐较大的匍匐型冷蒿个体。因此,匍匐型冷蒿个体比直立型冷蒿对地上空间利用的更充分,而直立型冷蒿个体则在土壤纵向的资源利用上占优势。
 | 图2 直立型冷蒿根系(左)和匍匐型冷蒿(右)根系形态图Fig.2 The morphology of erect type’ roots (left) and creeping type’ roots (right) of A. frigida |
2.3 匍匐型冷蒿的分株能力
在适宜的水分条件下,冷蒿被沙埋的枝条发展成匍匐茎并向上生长出不定芽,向下产生不定根。匍匐茎是冷蒿为其克隆生长增繁地上枝条、地下不定根的营养器官。冷蒿通过这种分枝方式实现其在生境内获取必需资源的策略。随着时间的推移,不定芽会慢慢长成枝条,不定根则汲取土壤中的养分并向土壤深处活动,形成冷蒿个体。分株最终脱离基株,成为单个的冷蒿植株,完成了冷蒿数量上的扩张。冷蒿对环境的适应特性是通过其生长特征(生物量、分株数) 和形态特征(匍匐茎、节间距和不定根间距) 来表现的,生长型主要包括分株高、枝条长、枝条数量等[ 30]。
通过对30株匍匐型冷蒿的调查发现,产生的分株数多数在10株以下,最少的产生3株,最多的可达16株,扩散的距离为4~35 cm,大部分分株与基株距离在10~20 cm之间。这与匍匐型冷蒿的枝条长度相关。这种繁殖特性是在沙漠化地区其不断被沙埋、被放牧牲畜啃食等环境干扰下种群能够生存下去的重要因素。
2.4 不同沙埋时间对冷蒿枝条生根的影响
被沙埋的冷蒿枝条产生不定根与否以及数量的多少与沙埋时间有关,分析认为与生长期间的气温、降水量有很大关系。实验地区6,7,8月月平均气温分别为20.28,23.44,23.25℃。降水主要集中在4-7月,虽然8月份降水较少,但从整体上来看,适度的气温和降水量都是冷蒿枝条生根的有利条件。
由 表2可知,在不同沙埋时间处理下,冷蒿生根枝条数在每株冷蒿处理枝条中所占比例、每株的平均最长根长度及每株生根数总量上均表现为匍匐型冷蒿>直立型冷蒿,平均每株冷蒿的生根数量在0~5 cm根长度中占大部分。而且第1次处理下2种表现型冷蒿根的长度在5~10 cm与10 cm以上所占比例大致相近,第2次处理下却表现出根长为5~10 cm的根数多于10 cm以上的趋势。可以推测出在适宜环境下,处理时间越长越利于冷蒿枝条生根。此外,同一处理时间下,2种表现型冷蒿的平均最长根长度及各级根长间的数量均接近,这说明同一条件下2种表现型冷蒿的枝条生根能力无差异。
| 表2 2种表现型冷蒿枝条的沙埋处理结果比较 Table 2 The comparison of sand buried branches of two phenotypes of A. frigida |
2.5 封育围栏内冷蒿种群扩散特征
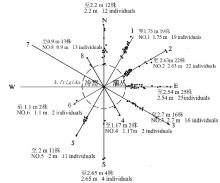
自1994年沙地围栏封育后,在无人为干扰及牲畜采食、践踏的自然条件下,冷蒿种群从最初的仅几株发展到1998年的119株以上,以选定的测量植株为基点,扩散最远的冷蒿植株与基株灌丛间距离达到2.7 m( 图3),位于东偏南30°方向。并且冷蒿在各条样线上成斑块状聚集分布。基株的西方无冷蒿生长,大部分冷蒿主要集中在东北方向、东方而且与基株间的距离也较远,西南方向有少数冷蒿分布但西方和西偏北30°方向的冷蒿却更少。
 | 图3 1998年恢复样地的冷蒿种群扩散图Fig.3 The diffusion of A. frigida population in restoration plot in 1998 |
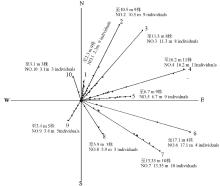
2009年的调查结果显示:在十多年的无干扰条件下,冷蒿种群无论在数量上还是规模上都远远超过了1998年。在冷蒿基株周围,扩散最远的冷蒿植株距离基株17.1 m,位于东偏南52°( 图4)。从整体上来看,冷蒿种群的扩散方向及分布情况与1998年调查结果类似,冷蒿植株主要分布在东北方向、东方,其次是东南方,西北方向和西南方向、西方冷蒿较少甚至没有冷蒿生长。而且在扩散距离上也是东北,东方和东南方的距离最远,西南、西北方向距离较近。
实地调查中发现,该恢复样地中的冷蒿植株均为直立型冷蒿。许多研究发现[ 10, 11, 19],直立型冷蒿枝条中生殖枝占绝大部分,营养枝较少,主要以产生种子的方式繁殖新个体。 图3和 图4中出现的冷蒿种群的扩散方向和距离的趋势类似。这种结果可能与冷蒿种子较小,种子散布时受风的影响较大,尤其是在科尔沁沙地这种风沙环境有关。科尔沁沙地冬春季节受蒙古冷高压控制,盛行偏西或偏北风[ 31], 所以冷蒿秋季成熟的种子顺着主风向方向散布的量最多,距离最远,而向相应的逆主风向方向散布的种子量少,距离最近。已有研究[ 8]表明,种子散布的数量和距离,能够反映种群数量动态的潜在能力或种群扩展空间生态位的潜在趋势。
 | 图4 2009年恢复样地冷蒿种群扩散图Fig.4 The diffusion of A. frigida population in restoration plot in 2009 |
从 表3中可知,从数量上来说,1~4,6,10号样线上的冷蒿分布较多,5,7,8,9号样线上相对较少均在5株以下,即北偏东、南偏东52°方向冷蒿数量比南偏东、南偏西和北偏西方向多,占种群的大多数。从冷蒿在每条样线上所占的总长度来看,变化趋势与数量相似,冷蒿的丛辐大小却表现出相反的趋势,南偏东52°的平均个体最大,丛辐为1.37 m,其次是南偏西和北偏西方向的冷蒿个体,北偏东方向冷蒿丛辐较小,在0.26~0.37 m之间。这可能与样线上冷蒿个体的数量多有很大关系。
| 表3 各样线上冷蒿个体的分布 Table 3 The distribution of A. frigida on each line transect |
3 结论与讨论
在长期的自然选择下,冷蒿进化出了许多适应干旱环境的生理生态的特征,其中之一便是形成了2种表现型即直立型冷蒿和匍匐型冷蒿。不同表现型冷蒿繁殖方式上也存在差异。直立生长的冷蒿为有性繁殖,丛生枝条密集,地上地下部分均为明显的独立个体;匍匐生长的多为克隆繁殖,枝条较稀疏,地下部分常有与其他植株连结的老枝条。在沙漠化地区围栏封育开始时冷蒿种群扩散以种子繁殖为主,随着植被的恢复物种增多,土壤水分减少,种间竞争强度更大,会出现克隆繁殖。冷蒿2种繁殖方式的结合不仅能提高和维持种群的扩散能力,其聚集群体更利于提高种群的生存和竞争力,使得它在植被更新、群落恢复过程中起着非常重要的作用[ 32, 33]。
从植株个体高度上来看,直立型冷蒿高于匍匐型冷蒿,并且前者的枝条总数、每株花序数均多于后者。但直立型冷蒿的平均丛辐小于匍匐型冷蒿,冷蒿个体间的生长差异的波动性较大。从冷蒿的单株地上生物总量结果中可知,直立型冷蒿的总量大于匍匐型冷蒿,且波动范围较广。在地上生物量的分配上,直立型冷蒿靠近基部的生物量小于匍匐型冷蒿,但基部以上的部分均大于后者。这与直立型冷蒿个体高于匍匐型冷蒿,后者的枝条较低矮有很大关系。
在干旱半干旱的沙地,季节性的降水对植株种子萌发及枝条生根具有较大的影响[ 34]。通过沙埋实验发现沙埋时间的长短及降水对冷蒿生根数量和最长不定根的长度影响较大,但同一沙埋时间处理下2种表现型冷蒿的不定根数量和最长不定根的长度无差异。直立型冷蒿是由1粒种子经过萌发发育而成,其根系分布较深,能充分利用土壤较深处的水分和养分。而匍匐型冷蒿由基株和分株构成,其分株能扩散到距离基株35 cm处,而且基株能产生多达16个分株,这就使得其地下根系构成的网络状结构在土壤的横向资源利用上远优于直立型冷蒿。
围栏封育样地的冷蒿种群以最原始植株为点逐渐向东北方与偏东方向扩散分布。同时植株在其扩散方向上呈斑块状聚集分布。造成这一现象可能是与冷蒿种子较小并能随着风向传播到较远距离,以及科尔沁沙地冬春季盛行偏西或偏北风有关。这一结论与杨允菲和祝玲[ 35]的研究即种子散布的数量及方向与地区的季风气候有很大的关系相符合。
与其他地区相比,科尔沁气候更干旱,土壤含沙量高,在其沙漠化恢复过程中,黄蒿、冷蒿等菊科植物和虫实( Corispermum dilutum)等藜科植物则会大量出现在植被演替后期[ 36, 37]。而冷蒿独特且有效的繁殖方式加快了冷蒿整个种群的扩散速度,这样就更利于科尔沁沙地退化生态系统恢复进程。因此,一方面可以结合2种表现型冷蒿的不同繁殖方式进行植被恢复,另一方面可以通过适当沙埋的方式促使冷蒿植株枝条生根产生分株以扩大其生长范围从而获得更多的资源,在进一步促进沙丘固定的同时,能为更多物种的入侵定居打下坚实的基础。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|