


{kind=link}
{kind=link}
{kind=link}
{kind=link}
松嫩草地两种广布豆科植物分布特征及其与土壤因子关系
引用本文
李强, 周道玮, 宋彦涛. 松嫩草地两种广布豆科植物分布特征及其与土壤因子关系. 23(1): 31-40
LI Qiang, ZHOU Dao-wei, SONG Yan-tao. The distribution features of two widespread legumes and their relationships with soil factors in Songnen grassland. Acta Prataculturae Sinica, 23(1): 31-40
Permissions
LI Qiang, ZHOU Dao-wei, SONG Yan-tao. The distribution features of two widespread legumes and their relationships with soil factors in Songnen grassland. Acta Prataculturae Sinica, 23(1): 31-40
Copyright©2014, 草业学报编辑部
本文属于开放获取期刊。
松嫩草地两种广布豆科植物分布特征及其与土壤因子关系
作者简介:李强(1984-),男,吉林农安人,在读博士。E-mail:liqiang@neigae.ac.cn
摘要
通过野外调查研究了松嫩草地中五脉山黧豆、野豌豆这两种广泛分布的豆科植物的地上生物量分布特征与土壤因子的关系。结果表明,五脉山黧豆地上生物量分布的最大值为286.25 g/m2,最小为51.73 g/m2,野豌豆的最大地上生物量分布为494.77 g/m2,最小为61.19 g/m2。两种豆科植物种群聚集水平主要在100~200 g/m2和100~300 g/m2。两种豆科植物地上生物量和群落高度、地上生物量、物种丰富度、多样性指数、均匀性指数之间均有显著的正相关关系,而与群落密度有显著负相关关系。两种豆科植物地上生物量与土壤有机碳、总氮含量呈现显著正相关关系,而与土壤pH、电导率呈显著负相关关系,与土壤总磷含量无显著相关关系。低豆科地上生物量下,群落特征和土壤因子随豆科植物地上生物量增加变化更大,而高豆科植物地上生物量下,群落特征和土壤因子随豆科地上生物量增加变化幅度较小。与野豌豆相比,群落特征和土壤因子随五脉山黧豆生物量增加的变化幅度更大。通过上述研究结果我们得出:51.73和61.19 g/m2是五脉山黧豆和野豌豆在群落中维持种群延续的最小单位面积地上生物量;两种豆科植物的生态性状可能对群落的结构和功能有促进作用,且单位质量的五脉山黧豆较野豌豆对群落结构的影响可能更强;土壤盐碱程度不会或至少不会单独构成对两种豆科植物分布的限制;磷可能是豆科植物分布的限制因子之一。这个研究为在本区内保护和发展这两种豆科植物提供了重要参考。
关键词:
豆科植物; 地上生物量; 群落特征; 土壤; 盐碱化
中图分类号:S812.2
文献标志码:A
文章编号:1004-5759(2014)01-0031-10
The distribution features of two widespread legumes and their relationships with soil factors in Songnen grassland
Abstract
Through field investigations, distribution features of aboveground biomass of
Keyword:
legume; aboveground biomass; community characteristics; soil; salinization
引言
豆科植物在草地植物资源中具有重要地位[ 1]。豆科植物具有低纤维、高蛋白含量特点,饲喂价值高,一般是优良的牧草[ 2],许多豆科植物还能为绿肥、蜜源、药材所用[ 3]。更重要的,豆科植物根瘤特有的固氮特性能将大气中的N2转化为植物可利用的形态,不仅能减少自身生长对土壤氮素的依赖,其固定的氮还能通过根部的分泌、死亡根的分解、动物粪便等途径转移给邻居植物利用[ 4, 5]。有研究报道在豆科与禾本科混播草地中,豆科植物固氮最多可达到草地地上部氮积累总量的46%,这对维持草地系统的生产力起到了重要作用[ 6]。豆科植物还具有高的枯落物质量,能够通过枯落物的快速分解加速养分循环,改善土壤养分状况[ 7]。土壤状况的改善可能提供更多的生态位给其他植物,对保护草地的物种多样性和增加资源互补性是有利的[ 8]。发达的根系也是豆科植物的普遍特征,许多豆科植物的根系分布可深至5 m,这不仅能够增加群体的水分利用和养分吸收效率,还能提高植被抗风、固土的作用[ 9]。鉴于豆科植物在草原系统中的重要价值,保护和利用豆科植物资源对发展草原经济和生态有重要意义。
保护和发展草原豆科植物资源需要对豆科植物的分布和建植规律有充分的认知,如了解豆科物种的地理分布、豆科物种分布与环境因子关系、豆科分布区的植被特征等都是有效利用豆科植物资源的前提。国外早有学者在局域内对天然草地豆科植物分布特征开展了研究,如Ehrman和Cocks[ 10]研究了西亚一年生豆科饲草分布的地理、生态特征,Beale等[ 11]研究了非洲西北部草地一年生豆科植物的分布及其与气候、土壤因子关系。国内对天然草原中豆科植物分布规律的研究较少,且多集中于豆科资源调查[ 9, 12, 13],也有一些研究针对区域自然条件从抗逆生理角度研究了一些豆科物种对干旱、盐碱等环境的适应能力,间接地预测了这些豆科物种在该区域内分布的可能性和成功性[ 14, 15, 16]。与国外比较,国内对豆科植物分布规律的研究,尤其是天然草地豆科植物分布规律的研究相对薄弱。
松嫩草地位于欧亚草原区东缘,地理和水文条件决定了松嫩草地独特的土壤特性——盐碱化[ 17]。受土壤理化性质限制,以羊草( Leymus chinensis)占绝对优势的盐碱化草甸是松嫩草地的主要植被。在羊草草甸中广泛分布着许多杂类草群落,这些杂类草群落以大小不一的斑块状镶嵌在草甸中,相邻的杂类草斑块包裹羊草群落形成杂类草区。豆科植物就生长在这些杂类草区中。这些豆科植物不仅改善了松嫩草地牧草种类单一的局面,且对草地的饲草产量和质量、生态稳定性有极大贡献[ 18]。保护和发展这些豆科物种要求我们了解这些豆科植物的分布特性,然而,很少有学者在该区域针对这个问题开展研究。尽管陶岩等[ 19]研究了该区域5种豆科植物总体的空间分布格局,但该研究并未在群落水平上具体揭示每种豆科植物分布规律,包括其与杂草区群落特征、土壤理化因子的对应关系。而这些信息恰是在天然草地中保护、引种利用豆科植物的关键背景。基于此,本研究通过野外调查实验针对本区内分布最广、数量最多,且产量和饲喂价值均较高的两种豆科植物——五脉山黧豆( Lathyrus quinquenervius)和野豌豆( Vicia cracca)进行这些信息的揭示,以为本区域这两种豆科植物资源利用提供关键理论支持。
1 材料与方法
1.1 研究区自然概况
研究地点位于中国科学院长岭草地农牧生态研究站(44°33' N, 123°31' E, 海拔145 m),地处松嫩草地腹地。该区属于温带半湿润半干旱温带大陆性季风气候,春季干旱多风且降水稀少,地表蒸发剧烈;夏季降水集中,冬季降雪较少。年均温4.9℃,无霜期140~160 d。年降水量300~500 mm,集中在6-9月,占总降水量的70%。年蒸发量1600.2 mm,约是降水量的3.5倍。地带性土壤为黑钙土。主要植被类型为羊草草甸。以多年生根茎禾草羊草为优势的草甸中,镶嵌分布着一定数量的杂草群落聚集区,包括扁蓿豆( Medicago ruthenica)、兴安胡枝子( Lespedeza davurica)、细叶胡枝子( Lespedeza hedysaroides)、五脉山黧豆、野豌豆等在内的豆科植物以零散或聚集的方式生长在这些杂草群落区内。一种豆科植物往往集中分布于特定的杂草群落区(如 图1中A、B、C)。其中,五脉山黧豆和野豌豆是数量最多、分布最广的两种豆科植物。
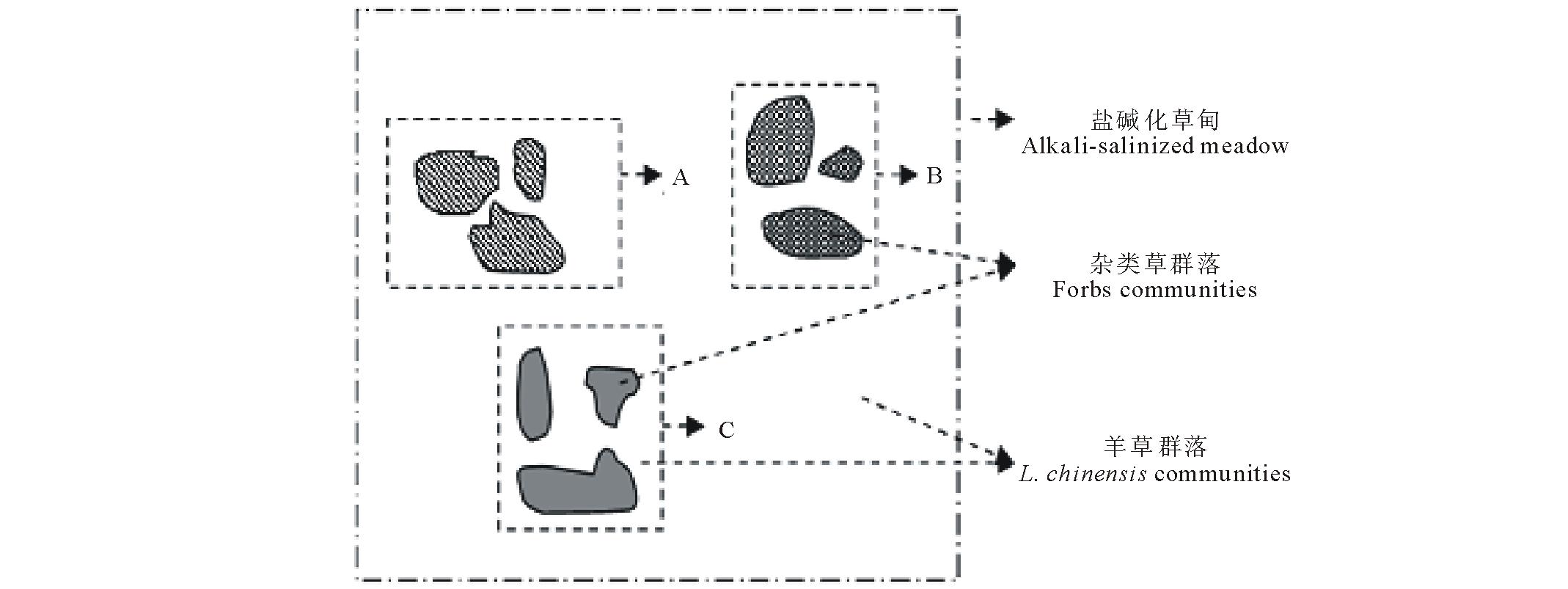 | 图1 盐碱化草甸杂类草群落分布特征Fig.1 The distribution feature of forb communities in alkaline meadow |
1.2 研究方法
2009年8月在1个1000 hm2的天然羊草草甸中选取五脉山黧豆和野豌豆分布的典型杂草群落区,根据两类杂草群落斑块格局特点,各分别设置一个50 m×100 m的样带。每个样带均分为100个5 m×5 m的小区,每个小区中央设置一个1 m×1 m的样方进行植物材料和土壤取样。查数每种植物密度后,齐地面收获地上生物量,目标豆科植物单独收获。取样过程中未发现有目标以外其他种豆科植物生长在样方内。地上植物材料在70℃下烘干48 h后称取干重。地上部分取样的同时,采用直径3 cm的土钻钻取0~20 cm深度土壤样品,对每层土壤,每个样方内随机钻取3钻混合为一份样品,土壤样品带回室内阴干,待测土壤理化性质。
1.3 土壤样品分析
每份风干土壤样品的一部分过2 mm筛后,实验室内使用PHS-3C pH仪和DDS-307电导率仪(上海,中国)测定土壤pH和电导率(水土质量比5∶1),另一部分过0.25 mm筛,采用重铬酸钾氧化法测定土壤有机质,土壤全氮用凯氏定氮仪(Kjeltec 2003 Analyzer Unit, FOSS TECATOR, Höganäs, Sweden)测定, 土壤全磷用钼锑抗比色法测定[ 20]。
1.4 数据处理
采用物种丰富度指数( R)、Shannon-Wiener 多样性指数( H)和均匀性指数( E)表示每个样方的群落结构特征,计算公式如下:
物种丰富度指数( R): R=S
Shannon-Wiener 多样性指数 ( H): H=-
均匀性指数 ( E): E=H/ln S
式中, S代表样方中的物种数目, Pi 表示第 i个物种个体数目占总个体数目的比例。
在每个豆科分布区内,根据调查样方内目标豆科植物的地上生物量,将全部样方进行分组。无目标豆科植物的样方为一组,考虑到野豌豆和五脉山黧豆个体和群体地上生物量的差异,从>0始,五脉山黧豆地上生物量每增加50 g以内(含50 g)的样方为一组,野豌豆地上生物量每增加100 g以内(含100 g)的样方为一组。统计每组内的样方数及在总样方中的比例,在每组内,统计目标豆科地上生物量的最大值、最小值、均值。不同组别群落特征和土壤理化性质的差异通过单因素方差分析后多重比较(Duncan)实现,各组豆科地上生物量和群落特征、土壤因子均值间的关系采用一般线性回归分析,显著性水平均为0.05。所有统计采用SPSS软件完成(SPSS 16.0 for windows, SPSS Inc., Chicago, IL, USA)。采用Sigma Plot 10.0(Systat software, Inc.,San Jose, CA, USA)完成作图。
2 结果与分析
2.1 豆科植物生物量分布特征
选定的五脉山黧豆分布区样带内,包含目标豆科植物的样方数为75个,占据了总调查样方数的3/4( 表1)。在含有五脉山黧豆的群落中,五脉山黧豆的最大地上生物量分布为286.25 g/m2,最小地上生物量为51.73 g/m2,未见有其地上生物量低于50 g/m2的样方。全部的100个样方按照规定方案分为6组(各组内豆科生物量均值见 表1)。其中,五脉山黧豆地上生物量在100~150 g/m2和150~200 g/m2组的样方数分别为24和21个,明显高于其他五脉山黧豆分布组。野豌豆的地上生物量分布与五脉山黧豆相似。分布有山野豌豆的样方数为75个,同样占据了总调查样方数的3/4( 表1)。在含有野豌豆的群落中,野豌豆的最大地上生物量分布为494.77 g/m2,最小为61.19 g/m2。所有的100个样方同样被划分为6组(各组内豆科地上生物量均值见 表1),其中,野豌豆地上生物量在100~200 g/m2和200~300 g/m2组的样方数分别为27和17个,明显高于其他野豌豆分布组。
| 表1 豆科植物地上生物量在杂草群落区中的分布特征 Table 1 The distribution feature of aboveground biomass of legumes in forb community area |
2.2 豆科植物生物量与群落特征关系
两类杂草群落区中,豆科植物地上生物量和群落高度、植物密度、群落地上生物量之间均有显著的线性相关关系( P<0.05;相关系数及方程见 图2)。其中,随着豆科植物地上生物量的增加,群落高度和群落地上生物量持续增加( 图2A、 图2C),而群落的植物密度逐渐降低( 图2B)。五脉山黧豆地上生物量与群落密度、群落地上生物量关系斜率绝对值较野豌豆更大,表明二者随五脉山黧豆地上生物量增加的变化速率更大。按豆科植物地上生物量由低到高,不同五脉山黧豆地上生物量组对应的群落高度均值范围为37.8~57.8 cm;植物密度均值范围为896~507 株/m2; 群落地上生物量均值范围为335.03~593.25 g/m2。 不同野豌豆地上生物量组对应的群落高度均值范围为43.2~75.9 cm;植物密度均值范围为902~526株/m2;群落地上生物量均值范围为381.95~733.35 g/m2( 图2)。在相邻低豆科地上生物量组别间,群落高度、植物密度、群落地上生物量差异明显,尤其是无豆科植物组和相邻组间,几乎都呈现显著性差异,而在高豆科地上生物量组间,这些指标的变化相对较小( 图2)。
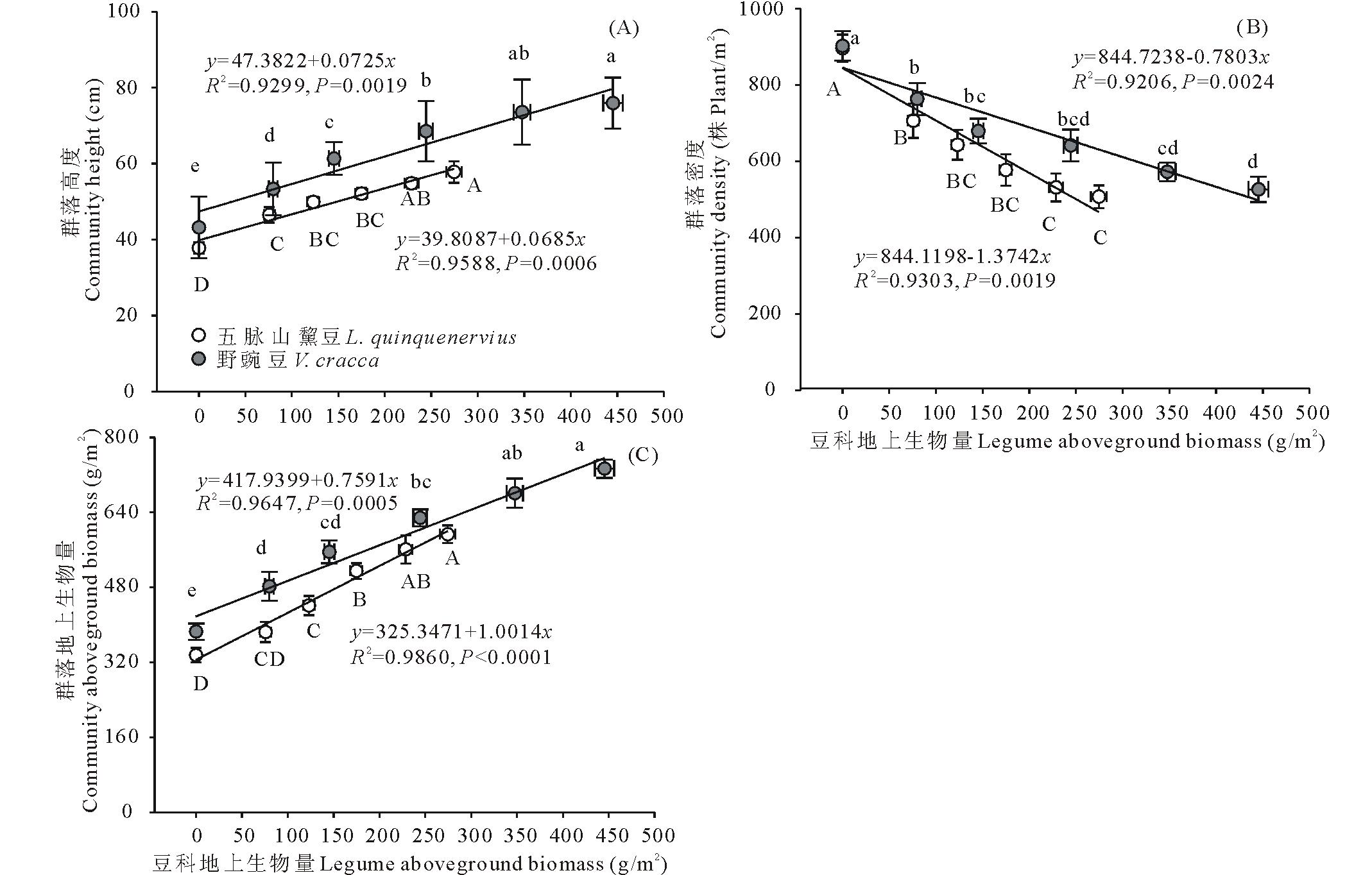 | 图2 豆科植物地上生物量与群落高度(A)、密度(B)、总地上生物量(C)的关系Fig.2 The relationships between legumes aboveground biomass and height(A), density(B), aboveground biomass(C) of community |
两类杂草群落区中,群落物种丰富度、多样性、均匀性和豆科植物地上生物量之间均有显著的线性正相关关系( P<0.05;相关系数及方程见 图3),并且三者均随群落中豆科地上生物量的递增而持续增加。五脉山黧豆地上生物量与三者关系斜率较野豌豆更大,表明3个变量随五脉山黧豆地上生物量增加的变化更快。同样的,按豆科植物地上生物量由低到高,不同五脉山黧豆地上生物量组对应的群落物种丰富度指数均值范围为5.08~11.27;Shannon-Wiener指数均值范围为0.88~2.36;均匀性指数均值范围为0.52~0.98。不同野豌豆地上生物量组对应的群落物种丰富度指数均值范围为4.63~9.89;Shannon-Wiener指数均值范围为0.68~2.20;均匀性指数均值范围为0.45~0.96( 图3)。与群落高度等相似,在相邻低豆科地上生物量组别间,群落物种丰富度、多样性、均匀性差异明显,尤其是无豆科植物组和相邻组间,均呈现显著性差异( P<0.05),而在高豆科地上生物量组间,这些指标的变化相对较小( 图3)。
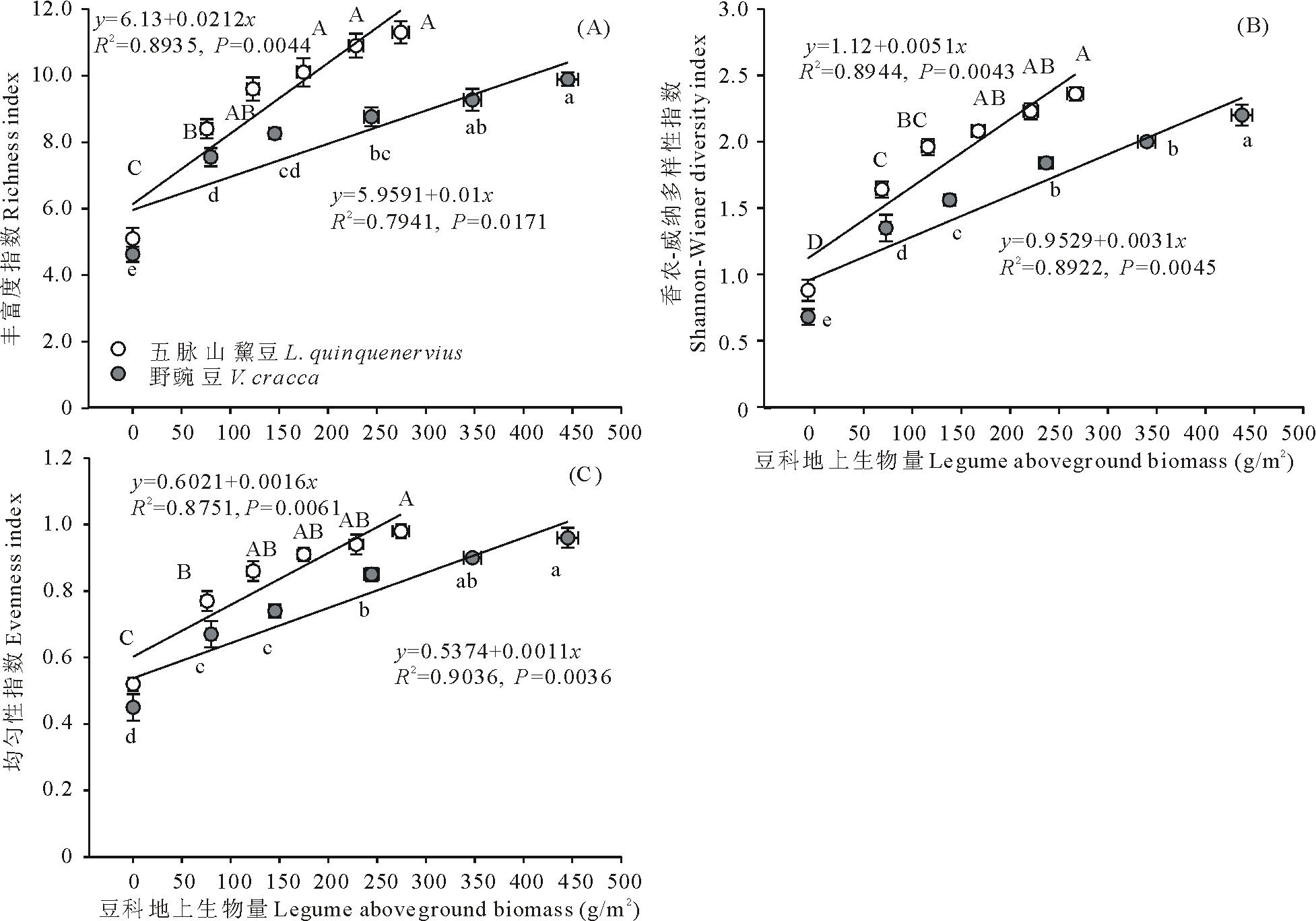 | 图3 豆科植物地上生物量与群落物种丰富度(A)、多样性(B)、均匀性(C)的关系Fig.3 The relationships between legumes aboveground biomass and species richness(A), diversity(B), evenness(C) of community |
2.3 豆科植物生物量与土壤因子的关系
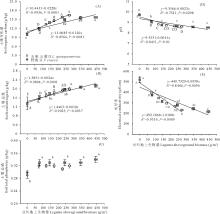
两类杂草群落区中,群落豆科植物地上生物量和土壤有机碳含量、总氮含量、土壤pH、土壤电导率之间均有显著的线性相关关系( P<0.05;相关系数及方程见 图4)。其中,土壤有机碳含量和总氮含量随群落豆科植物地上生物量的增加而增加( 图4A、 图4B),而土壤pH和电导率随群落豆科植物地上生物量增加而降低( 图4D、 图4E)。土壤总磷含量与群落豆科植物生物量间未发现有显著相关关系( P>0.05; 图4C)。除土壤总磷外,五脉山黧豆地上生物量与其他土壤因子关系斜率绝对值较野豌豆更大,表明这些因子随五脉山黧豆地上生物量增加的变化更快。按豆科植物地上生物量由低到高,不同五脉山黧豆地上生物量组对应的土壤有机碳含量、土壤总氮含量、土壤pH、土壤电导率均值范围为10.33~16.44 g/kg、1.36~1.99 g/kg、9.77~8.56、524~232 μS/cm。不同野豌豆地上生物量组对应的土壤有机碳含量、土壤总氮含量、土壤pH、土壤电导率均值范围为10.08~15.73 g/kg、1.31~2.14 g/kg、9.71~8.51、516~216 μS/cm( 图4)。相邻低豆科地上生物量组别间,土壤有机碳含量、总氮含量、土壤pH和电导率差异明显,尤其是无豆科植物组和相邻组间,均呈现显著性差异( P<0.05),而在高豆科生物量组间,这些土壤因子的变化相对较小( 图4)。对土壤总磷含量,仅发现野豌豆分布区0豆科生物量组与其他组间的显著性差异( P<0.05; 图4C)。五脉山黧豆分布区中,0豆科生物量组的土壤总磷含量均值为0.30 g/kg,其他组维持在0.32 g/kg左右;野豌豆分布区中,0豆科生物量组的土壤总磷含量均值为0.29 g/kg,而其他组维持在0.33 g/kg左右。
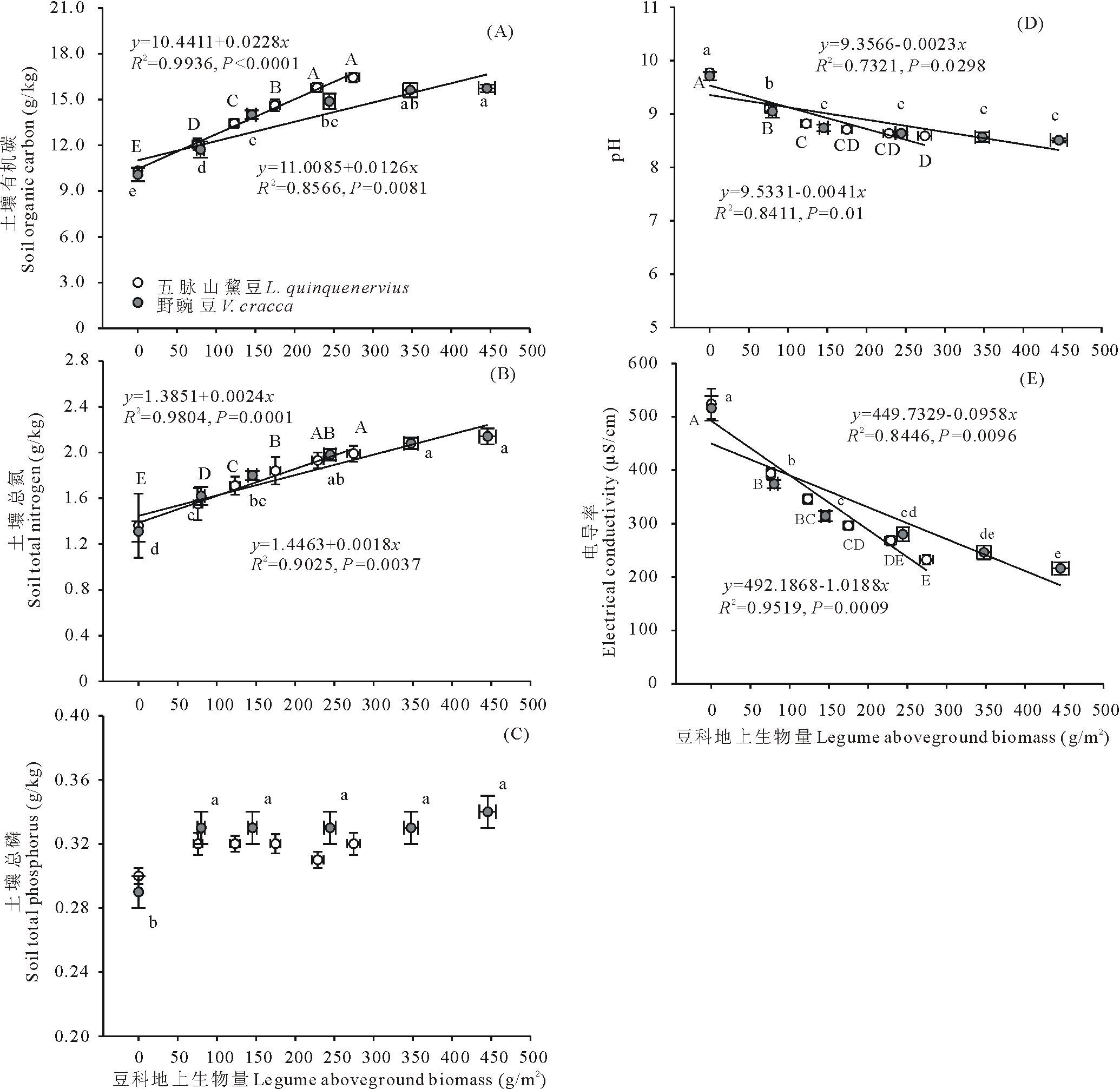 | 图4 豆科植物地上生物量与土壤因子的关系Fig.4 The relationships between legumes aboveground biomass and soil factors |
3 讨论
3.1 五脉山黧豆和野豌豆的分布特点
陶岩等[ 19]的研究中已表明五脉山黧豆和野豌豆在松嫩草地中表现聚集分布的空间格局,但该研究未具体给出两种豆科植物种群的聚集程度。而本文通过研究得出两种豆科植物在单位面积上的分布数量和各数量级所占面积比例。结果表明:五脉山黧豆和野豌豆种群聚集程度主要在100~200 g/m2和100~300 g/m2 水平。依据两种豆科植物在杂草区中被发现的最大单位面积地上生物量,这两个范围都处于两种豆科植物的中等聚集程度上。在两种豆科植物各自的调查样带内,含有目标豆科植物的样方均为75个,这可能意味着在相同大小的杂草区域内,两种豆科植物的分布面积相近。种群大小能够影响物种在群体中占据的生态位,进而影响物种的生存、繁殖以及环境变化下的适应性[ 21]。我们的研究结果发现, 五脉山黧豆和野豌豆的最小单位面积聚集量都高于50 g/m2 ,分别为51.73和61.19 g/m2,这可能是本区域内两种豆科植物在群落中维持种群延续的最小单位面积地上生物量。
3.2 两种豆科植物生物量与群落特征关系
豆科植物地上生物量和群落特征的关系信息能够为我们调研和保护豆科植物资源、认识豆科植物功能提供直观的依据。我们的研究结果发现, 两种豆科植物的地上生物量与群落高度、群落地上生物量间有显著的正相关关系。含野豌豆的群落具有更高的群落高度和更大的群落地上生物量。分析其原因可能包括, 1)与两种豆科植物的性状有关。两种豆科植物一般较禾本科植物叶片繁密、多枝、个体地上生物量更大。一方面,豆科植物在群落中数量越多可能增加取样效应(sampling effect)对群落地上生物量的影响[ 22, 23]。另一方面,豆科植物在群落中存在能够增加群落冠层的密闭度和光资源的截获,而其他植物可能通过增加个体高度的方式实现对光资源的竞争,间接增加群体高度和地上生物量[ 24, 25]。尤其野豌豆,不仅分枝更多、个体地上生物量更大,且具有蔓生的特点,能够依附在其他植物的茎上不断延伸个体,其主茎长度可达1 m以上,这增加了对光的截获强度,促使了其他植物群体高度和地上生物量的增加。2)两种豆科植物存在的群落中往往伴生有牛鞭草( Hemarthria sibirica)、箭头唐松草( Thalictrum simples)等植物,这些植物本身具有较高的高度,也能通过光竞争间接促进群体生长。3)豆科植物地上生物量和土壤养分呈正相关关系,和土壤盐碱程度呈负相关关系( 图 4),高的土壤肥力和低的盐碱胁迫可能直接促进豆科和邻居植物的生长。研究结果显示, 豆科植物地上生物量和群落植物密度之间有显著负相关关系。这主要是因为在豆科植物数量增加情况下,群落的物种组成发生了明显变化,原初密度较大的禾草群体不断被个体更大的豆科植物和杂草物种取代,导致总体密度的下降。
伴随豆科植物地上生物量不断增加,群落中禾草的绝对优势被打破,加之与豆科植物量协同增加的土壤养分,这些因素可能在群落内为更多物种提供生态位[ 8],导致群落物种丰富度、多样性指数、均匀性指数的增加。在结果中我们也发现:群落密度、物种丰富度、多样性和均匀性随五脉山黧豆生物量增加较随野豌豆地上生物量增加的变化速率更大,且相同两种豆科地上生物量下,上述变量在含五脉山黧豆的群落中具有更高值。这表明伴随五脉山黧豆数量的改变,群落物种替换和结构变化的速率更大。而且,从土壤数据中发现:在低豆科地上生物量下,五脉山黧豆生境的土壤条件并未优于野豌豆。尽管当前的研究还无法断定其背后的机理,但我们推定单位质量的五脉山黧豆较野豌豆对其他植物建植的促进作用(facilitation effect)更强。
众所周知土壤本底条件可能直接影响群落特征,包括豆科植物在群落中的数量[ 10, 11],而豆科植物也可通过对土壤的改善作用或资源的修饰作用间接地影响植物的建植和生长[ 6, 7, 8, 24, 25]。尽管我们对两种豆科植物地上生物量与杂草群落特征的关系给予了解释,但据当前野外研究的结果,我们仍无法判定土壤条件和豆科植物二者对杂草群落特征的决定作用,这需要进一步的因子控制实验来验证。
3.3 两种豆科植物生物量与土壤因子关系
我们的研究结果发现, 土壤有机碳、总氮、土壤pH、电导率与豆科地上生物量存在显著相关关系。但土壤条件和豆科植物数量之间相互的决定关系却很难确定。因为自然生态系统中,土壤和植物存在相互作用,即土壤条件能够影响植物的定居,而定居的植物又将通过生命活动和物质循环途径反馈作用于土壤[ 26],尤其豆科植物,其对土壤条件可能有显著的改善作用。在盐碱化草地中,土壤盐碱程度是制约植物建植的首要因子[ 27, 28],土壤的高度盐碱化能通过抑制植物种子的萌发和幼苗的生长影响植物的建植[ 29]。然而,杨光[ 30]利用两种豆科植物在碱性盐胁迫下的发芽研究发现,即使土壤pH达到11.5以上,两种豆科植物种子的发芽率仍能达到83%以上,而在包括裸碱斑在内的整个松嫩草地,土壤的盐碱程度都要低于pH=11.5的水平[ 31]。因此,松嫩草地的土壤盐碱化状况可能不会、或至少不会单独构成两种豆科植物建植的限制因素。但考虑自然界中环境因子的协同变化,如在研究本区域内,表土盐碱化程度往往匹配着土壤干旱、养分贫瘠程度,因此并不排除盐碱胁迫协同其他因子共同限制两种豆科植物分布。豆科植物本身具有固氮的特性,因此土壤氮不可能成为限制豆科植物分布的因素,也未见有相关的报道。对于土壤磷水平,李强等[ 32]和宋彦涛[ 33]在本区域内的研究报道: 未退化羊草草地的土壤0~20 cm层总磷含量均低于0.20 g/kg,这个数值要低于本研究中杂草区的最低土壤磷含量,因此土壤磷可能是这两类豆科植物分布的限制因子之一。但对任意豆科植物分布区,在含豆科植物的各组样方中,土壤总磷含量并未表现显著差异,这可能意味着土壤磷对两种豆科植物的分布限制有个阈值,而超过这个阈值,土壤磷即不构成对两种豆科植物生长数量的限制。而根据我们的研究结果,如果磷确是两种豆科植物分布限制因子,这个阈值应该是有五脉山黧豆和野豌豆分布的最小土壤磷水平,分别是0.29和0.27 g/kg。综上,对本研究结果中土壤有机碳、总氮、土壤pH、电导率随两种豆科植物地上生物量增加的变化,可能有两个解释:1)尽管这些变量不会或至少不会单独构成限制两种豆科植物分布的因素,但它们附和了其他限制因子的变化而变化,譬如土壤水分或其他因子。但同时,两种豆科植物本身一定程度上也在改善土壤条件。因此,这些土壤因子在不同豆科植物数量的群落中的差异是土壤条件自身的变化和豆科植物改良土壤的共同作用结果;2)本区域内任一土壤因素均不构成对两种豆科植物分布的限制,两种豆科植物聚集程度的变化完全取决于其种子散布的随机性[ 34],而土壤性质的变化则源于不同数量豆科植物改良土壤作用的差异。尽管土壤性质与两种豆科植物生物量之间关系的机理还有待更深入的研究,但本文直观地给出了两者的对应关系,这对我们保护和发展这两种豆科植物提供了重要参考。4 结论 野外调查结果表明, 五脉山黧豆和野豌豆种群聚集程度主要在100~200 g/m2和100~300 g/m2 水平,五脉山黧豆和野豌豆的最小单位面积聚集量分别为51.73和61.19 g/m2,这可能是两种豆科植物在群落中维持个体生存和种群延续的最小单位面积地上生物量。群落高度、地上生物量、物种多样性随两种豆科植物地上生物量的增加而增加,这可能与两种豆科植物的性状和促进效应有关。五脉山黧豆存在下,群落物种替换和结构变化的速率更大。结合土壤数据,我们推定单位质量的五脉山黧豆较野豌豆对其他植物建植的促进作用更强。土壤有机碳、总氮、土壤pH、电导率与两种豆科植物地上生物量存在显著相关关系。而参考以往研究,我们得出这些土壤因子不会或至少不会单独构成对两种豆科植物分布的限制。磷可能是豆科植物分布的限制因子之一。因此,本研究中涉及的土壤因子随豆科植物数量的变化可能是协同了其他豆科植物分布限制因子的变化而出现的,或由豆科植物对土壤的改良作用决定的。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|