

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
青海湖地区紫花针茅草原群落特征
引用本文
岳鹏鹏, 卢学峰, 叶润蓉, 周玉碧, 杨仕兵, 彭敏. 青海湖地区紫花针茅草原群落特征. 23(4): 10-10
YUE Peng-peng, LU Xue-feng, YE Run-rong, ZHOU Yu-bi, YANG Shi-bing, PENG Min. Community characteristics of Stipa purpurea stepp in the Qinghai Lake region . Acta Prataculturae Sinica, 23(4): 10-10
Permissions
YUE Peng-peng, LU Xue-feng, YE Run-rong, ZHOU Yu-bi, YANG Shi-bing, PENG Min. Community characteristics of Stipa purpurea stepp in the Qinghai Lake region . Acta Prataculturae Sinica, 23(4): 10-10
Copyright©2014, 草业学报编辑部
本文属于开放获取期刊
青海湖地区紫花针茅草原群落特征
作者简介:岳鹏鹏(1981-),女,山东德州人,讲师,博士。E-mail:yue_pengpeng@163.com
摘要
选择青海湖地区典型紫花针茅草原作为研究对象,以大量的样方调查为基础,采用等级聚类、TWINSPAN (two-way indicator species analysis)、DCA (detrended correspondence analysis)、CCA (canonical correspondence analysis)、物种多样性,相关分析等多元数量分析方法,系统研究了紫花针茅草原的群落特征、物种多样性、群落与环境因子的关系。结果表明,所调查的紫花针茅草原是物种组成相对较简单、菊科和禾本科植物最多、北温带分布的属为主体的草原群落。青海湖地区的紫花针茅草原群落可分为4个群丛类型:紫花针茅+多枝黄芪群丛,紫花针茅+矮嵩草群丛,紫花针茅+西北针茅群丛和紫花针茅+川青早熟禾群丛。影响青海湖地区紫花针茅草原群丛分布的主要环境因子首先是以年均降雨量和年均温为代表的气候因子,其次是土壤水分、有机质和全氮磷钾等土壤因子。该区紫花针茅草原群丛均匀度指数和Simpson指数的变化趋势基本一致,紫花针茅+川青早熟禾群丛是各群丛中最高的。物种丰富度受气候和土壤因子的影响大,但Simpson指数和均匀度指数与所调查的所有环境因子都没有显著相关性。本研究揭示了紫花针茅草原群落的内在规律,对维护青海湖地区生态系统平衡具有重要意义。
关键词:
紫花针茅; 数量分类和排序; 物种多样性; 青藏高原
中图分类号:S812
文献标志码:A
文章编号:1004-5759(2014)04-0010-10
Community characteristics of Stipa purpurea stepp in the Qinghai Lake region
Abstract
The classification, association and distribution characteristics and relationships between the distributions and environmental factors of Stipa purpurea steppe in the Qinghai Lake region were studied. A combined transect and quadrat method was used to collect field data. Multivariate statistical methods, including cluster analysis, TWINSPAN (two-way indicator species analysis), DCA (detrended correspondence analysis), CCA (canonical correspondence analysis) and correlation analysis, were used to calculate the data on the basis of investigating a large number of vegetation communities in this region. The species composition of S. purpurea in the study area was relatively simple. The dominant families of seed plants were Gramineae and Compositae, and the dominant aerial types of genera were North temperate types. The community in the area could be classified into 4 associations: Ass. S. purpurea+Astragalus polycladus, Ass. S. purpurea+Kobresia humilis, Ass. S. purpurea+Poa indattenuata, Ass. S. purpurea+Sinochasea krylovii. The major environmental factors influencing the distribution of steppe types in the area were first climate factors (mainly average annual precipitation and temperature), then soil factors (mainly soil moisture, organic matter and N, P, K). Changes in evenness index and Simpson index of S. purpurea stepp associations were basically the same, and the indexes of Ass. S. purpurea+Poa indattenuata were the highest. Richness index was largely affected by climate and soil factors, but the Simpson index and evenness index of all environmental factors investigated were not significantly correlated with area. The present study provides the ecological parameters of S. purpurea steppe in the Qinghai Lake region and would be a basis for maintaining the ecosystem equilibrium of ecologically fragile zones.
Keyword:
Stipa purpurea ; quantitive classification and ordination; species diversity; Qinghai-Tibet Plateau
引言
亚洲大陆中部素有“ 世界屋脊” 之称的青藏高原, 是一个独特的地理单元, 其环境气候的特殊性、高寒生态类群的特有性和生态系统的脆弱性在我国乃至世界都有典型的代表意义。青海湖地区因其重要的地理位置、独特的自然环境以及与此相适应的生物多样性格局受到学术界的普遍关注。近年来, 由于气候变化和人类活动的综合影响, 青海湖流域已经出现了青海湖水位下降、土地沙漠化趋势加剧、草场植被破坏日益严重、渔业资源锐减、野生动植物和鸟类栖息地环境恶化等一系列的生态环境问题。
紫花针茅(Stipa purpurea)草原是青藏高原高寒草原中分布面积最大、最重要的群落类型[1, 2, 3]。它不仅在高山构成一定宽度的垂直带, 而且在高原开阔面上构成高寒草原水平地带景观, 对景观和生态系统多样性的保护维持意义重大。另外, 紫花针茅草原在防风固沙、水土保持等维护自然环境稳定性等方面起着重要作用。同时还为草地畜牧业的发展提供了不可缺少的物质基础。由于紫花针茅草原分布区域辽阔, 各地环境有异, 其分布的海拔和群落特征也有明显不同。刘洋等[4]研究了青海巴滩地区紫花针茅草原群落中出现频率较高的14个物种之间的交互联结性, 结果表明紫花针茅草原群落总体种间关联性为负相关, 种对间没有显著相关性, 群落尚处在演替初期阶段, 或群落受到外界严重干扰。但对沱沱河地区紫花针茅群落12个主要种的种间联结性进行分析时, 发现该区紫花针茅群落优势种多物种间显著正关联, 存在稳定共存的种间关系, 建群种与其他物种间联结性不显著, 在群落中能够稳定存在并占据优势生态位, 群落处于植被演替过程的一个相对稳定的阶段[5]。该两项研究中不同地区的紫花针茅草原表现了不同的特征。马世震等[6]、全晓毅等[7]研究证明退化草地植被优势物种发生了变化, 紫花针茅、早熟禾(Poa alpigena)等植物种生长势减弱, 物候期推迟, 群落优势植物逐步演替为以菊科、豆科等杂类草植物为优势种的群落, 物种多样性指数下降。祁彪等[8]对退化高寒干旱草地植物群落进行调查研究时, 发现在长期的放牧利用过程中, 草地群落的优势物种由原始的赖草(Leymus secalinus)、早熟禾逐渐演变为紫花针茅, 其伴生种也发生了明显的相互更替。Liu等[9]研究了青藏高原腹地紫花针茅种群的遗传多样性, 发现紫花针茅居群内的遗传多样性相对较低, 居群间的遗传差异较大, 说明遗传分化主要是发生在居群之间; 遗传距离和地理距离有较高的一致性; 其遗传多样性随着经纬度的增加而增加, 与海拔没有明显关系, 意味着高山植物的遗传多样性可能受湿度和温度的影响。综上所述, 紫花针茅的研究虽不断增多, 但主要是限于局部地区的单项研究, 对大尺度范围内紫花针茅草原群落的群落特征、内部分类、空间分布、物种多样性和生态环境变化特征以及它们之间的相关关系的综合性系统研究较少, 群落自身的生态特点、内部的分布规律等还存在很多未知问题。
本文以青海湖北岸分布的大片紫花针茅草原为研究对象, 在东西向系统取样的基础上, 采用数量分析方法, 研究紫花针茅草原经向分布群丛的特征、多样性变化以及与环境因子的关系, 旨在揭示紫花针茅草原群落的内在规律, 为维护生态脆弱区生态系统平衡和资源的可持续利用提供数据基础。
1 材料与方法
1.1 研究区域概况
青海湖地区整个地势自南向北渐低, 湖岸发育着多级阶地, 湖盆为宽阔平坦的冲击洪积扇平原, 湖东地区则为风沙地貌。该区气候寒冷干燥, 全年以西北风和西风为主, 光照充足, 太阳辐射强烈, 具有明显的高原大陆气候特征。以刚察县为例, 年平均气温-0.6~-5.7℃, 气温日较差13.3~16.5℃, 相对极端最低温度可达-31.0℃, 极端最高温25℃, 年均降水量324.5~522.3 mm, 且多集中在6-9月, 年蒸发量1273.7~1847.8 mm[10]。本区主要的土壤类型为高山草甸土、山地草甸土、黑钙土、风沙土和盐渍土等, 其植被类型主要为紫花针茅草原。
本研究区域范围为N 36° 15'~38° 20', E 97° 50'~101° 20', 主要包括天峻县和刚察县, 各样地概况见表1。
| 表1 样地概况 Table 1 General conditions of plots |
1.2 研究方法
1.2.1 样地设置和数据采集
选择青海湖地区北部连续分布的紫花针茅草原设立研究区(表1), 在研究区紫花针茅分布的典型地段由西向东依次设置5个样地(样地间隔≥ 30 km), 各样地分别设置5~6条样带(表1), 相邻样带至少间隔500 m, 样带方向随机设定。每条样带上设置10个1 m× 1 m的样方, 第一个样方在样带0~9 m间随机选定, 后续样方沿样带以10 m间隔依次等距设定。记录每个样方中植物的种类组成、分盖度和频度、群落总盖度以及物候等群落参数。同时用GPS测定记录样方的经纬度和海拔等环境因子。各样带上取其10个样方相关参数的算术平均值, 作为后续计算分析的基础数据。研究区在每一个样带中用TSC-1土壤水分快速测试仪测定土壤水分, 并用土钻随机取0~15 cm的土样5个, 充分混匀, 取适量放入铝盒并用保鲜纸盖严封, 详细编号后带回供室内分析测定土壤理化性质。所有野外调查工作于2005年的夏季8月份生物量高峰期完成, 在研究区5个样地上共调查样带27条, 1 m× 1 m样方270个。
各样带的多年平均气温、降雨量、湿润度等气候因子由中国科学院地理科学与资源研究所人地系统主题数据库获得, 采用的是中国1 km2气象拟合数据提取值(数据来源:人地系统主题数据库http://www.data.ac.cn)。
1.2.2 土样测定
土样带回实验室后, 采用重铬酸钾容量法测定有机质含量; 重铬酸钾— 硫酸消化法测定土壤全氮; 碱解扩散法测定速效氮; 重铬酸钾— 硫酸消化法测定土壤磷; 碳酸氢钠法测定速效磷; 氢氧化钠熔融— 火焰光度计法测定土壤全钾; 醋酸铵— 火焰光度计法测定速效钾; 电位计法测定土壤pH值[10, 11]。
1.2.3 群落数量分析方法
共调查270个样方隶属于27条样带, 记录种子植物58种。建立了两个矩阵:1)由27条样带58种植物的重要值组成的植被矩阵; 2)由27条样带15个环境参数组成的环境属性矩阵, 应用PCORD 5.0程序进行数量分类和排序。
重要值的计算由于草本植物个体数统计较为困难, 为使计算的各种指数更具代表性, 便于不同群丛间的比较, 本文采用各物种的重要值代替个体数进行计算, 其公式如下:重要值(Ⅳ )=(相对频度+相对盖度)/2
2)数量分类方法
本文结合使用双向指示种分析(two-way indicator species ananlysis, TWINSPAN)方法和等级聚合方法对青海高原紫花针茅草原群落进行数量分类。
3)排序方法
采用除趋势对应分析(detrended correspondence analysis, DCA)和典范对应分析(canonical correspondence analysis, CCA)对紫花针茅草原群落进行排序和与环境因子的关系研究。
4)α 多样性计算公式
α 多样性用丰富度指数、物种多样性指数、均匀度指数3类多样性指数分析讨论, 公式如下:
物种丰富度(richness index): R=S
均匀度指数(eveness index): E=-∑ (Piln Pi)/ln S
群落物种多样性指数(diversity index):
Shannon-wiener 指数: H'=-∑ (Piln Pi)
Simpson 指数: D=∑ Ni(Ni-1)/[N(N-1)]
式中, S为群落中的总种数; N为所在样方的各个种重要值之和; Ni为种i的重要值; 相对重要值Pi=Ni/N。
2 结果与分析
2.1 种子植物区系组成
所调查研究区内紫花针茅草原群落共出现种子植物58种, 隶属45属, 21科(表2)。区内紫花针茅草原植物种同样以菊科(12种)和禾本科(10种)为主, 二者分别占了总种数的20.69%和17.24%。其次, 豆科、莎草科、玄参科、蔷薇科和毛茛科各4种, 分别占总数的6.90%。其他还有龙胆科、伞形科、百合科等14科, 各仅有1~2种, 共占总种数的27.59%。
| 表2 研究区紫花针茅草原群落种子植物组成情况 Table 2 The vascular species composition of S. purpurea steppe |
按吴征镒[12]关于中国种子植物属划分的15个分布区类型, 研究区紫花针茅草原群落中的45个植物属可划分为8个类型(表3)。其中, 北温带分布属的数量占首位(27个), 世界分布属数量次之(6个), 中国特有属1个, 其他5种分布类型共21属, 共占总属数的24.44%。
| 表3 研究区紫花针茅草原群落种子植物属的分布区类型 Table 3 The generic a real-types of seed plants in S. purpurea steppe |
2.2 紫花针茅草原群落数量分类
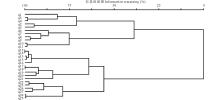
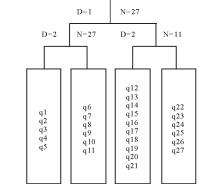
对植被矩阵采用可变类平均法和Euclidean Distance距离公式进行聚类, 结果如图1所示, 在信息保留量55%处, 27条样带可归入4个组, 代表对应的4个群丛类型。用TWINSPAN对该矩阵进行等级分划, 二级分划的结果和等级聚类的结果一致, 27条样带也可以分为4个组(图2)。因此将研究区的紫花针茅草原群落划分为4个组, 代表4个对应的有生态意义的群丛类型。
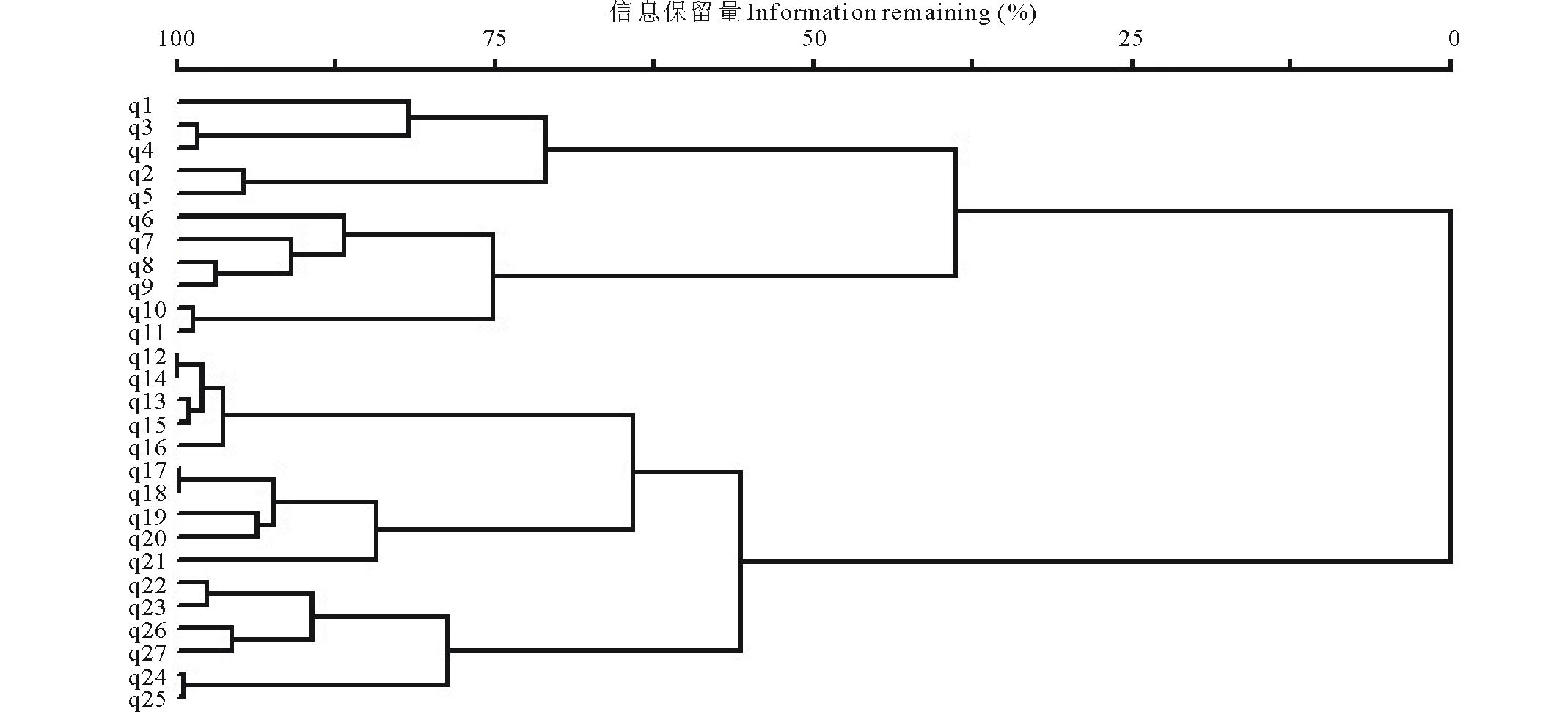 | 图1 研究区聚类分析结果Fig.1 The results of cluster analysis of S. purpurea steppe |
紫花针茅+多枝黄芪(Astragalus polycladus)群丛(Ⅰ ):该群丛类型包含样带q1~q5, 分布在天峻县快尔玛地区。群落总盖度为70%~76%, 平均总盖度73.40%, 紫花针茅相对盖度为54.24%。主要伴生种有伊凡苔草(Carex ivanovae)、弱小火绒草(Leontopodium humilum)、 小早熟禾(Poa caliopsis)、 多裂委陵菜(Potentilla multifida)、高山嵩草(Carex parva)、大花嵩草(Kobresia macrantha)等。
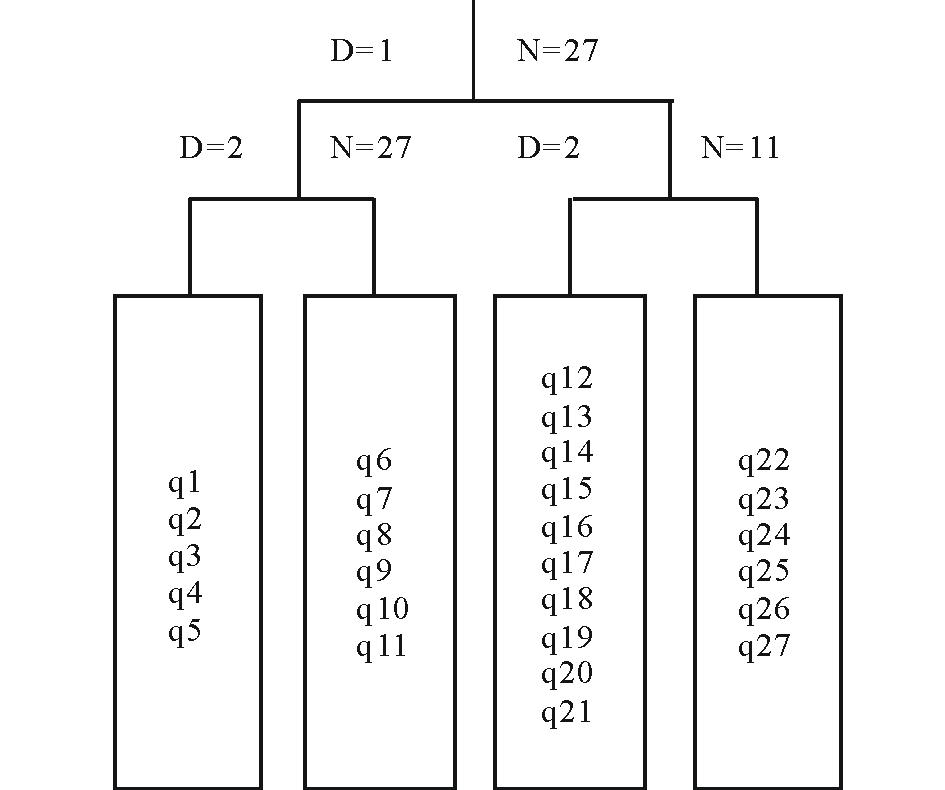 | 图2 TWINSPAN分类结果树状图Fig.2 The result dendrogram of TWINSPAN of S. purpurea steppe N代表样方数, D代表分类次序数, q1~q27指样号, Ⅰ ~Ⅳ 表示群丛类型。N indicates the nuber of transects. D means the division order, q1~q27 mean transect numbers. Ⅰ ~Ⅳ indicate the ecological associations. |
紫花针茅+矮嵩草(Kobresia humilis)群丛(Ⅱ ):该群丛类型包含样带q6~q11, 分布在天峻县县城以南。群落总盖度为67%~84%, 平均总盖度77.25%, 紫花针茅的相对盖度为45.11%。主要伴生种有钉柱委陵菜(Potentilla saundersiana)、多枝黄芪、伊凡苔草、小早熟禾、短穗兔儿草(Lagotis brachystachya)、多裂委陵菜、川青早熟禾(Poa indattenuata)等。
| 表4 各排序轴特征值及各排序轴与环境因子的相关系数 Table 4 The eigenvalues of ordination axes and the correlation coefficients between ordination axes and environmental factors |
紫花针茅+川青早熟禾群丛(Ⅲ ):该群丛类型包含样带q12~q21, 分布在刚察县宁秀和三角羊场附近, 包括2、3两个样地。群落总盖度为57%~79%, 平均总盖度68.65%, 紫花针茅相对盖度为44.70%。主要伴生种有西北针茅(Stipa krylovii)、大花嵩草、多枝黄芪、伊凡苔草、三辐柴胡(Bupleurum triradiatum)、沙蒿(Artemisia desertorum)、猪毛蒿(Artemisia scoparia)、赖草、楔叶山莓草(Sibalddia cuneata)、冰草(Agropyron cristatum)、阿尔泰狗哇花(Heteropappus altaicus)等。紫花针茅+西北针茅群丛(Ⅳ ):该群丛类型包含样带q22~q27, 分布在刚察县样地5。群落总盖度为61%~90%, 平均总盖度78.28%, 紫花针茅相对盖度为63.70%。主要伴生种有川青早熟禾、多枝黄芪、异叶青兰(Dracocephalum heterophyllum)、沙蒿、三辐柴胡、鳞叶龙胆(Gentiana squarrosa)、大花嵩草等。
数量分类划分的4个群丛, 均是紫花针茅占据绝对优势地位的群丛, 但各群丛结构还是存在差异。从上面各群丛的描述可以看出群落总盖度和紫花针茅的相对盖度不同。从4个群丛主要种的重要值变化情况(图3)来看, 样地5群丛Ⅳ 中紫花针茅重要值在4个群丛中最大, 包含样地3和样地4的群丛Ⅲ 中紫花针茅重要值在4个群丛中最小, 但这两个群丛其主要种的重要值比例比较接近。在聚类分析中群丛Ⅳ 和Ⅲ 聚到了一起后才与天峻地区的两个群丛聚合。群丛Ⅱ 中矮嵩草和钉柱委陵菜的重要值比其他群落明显高出很多, 说明该群落有一定的湿润草甸性质。
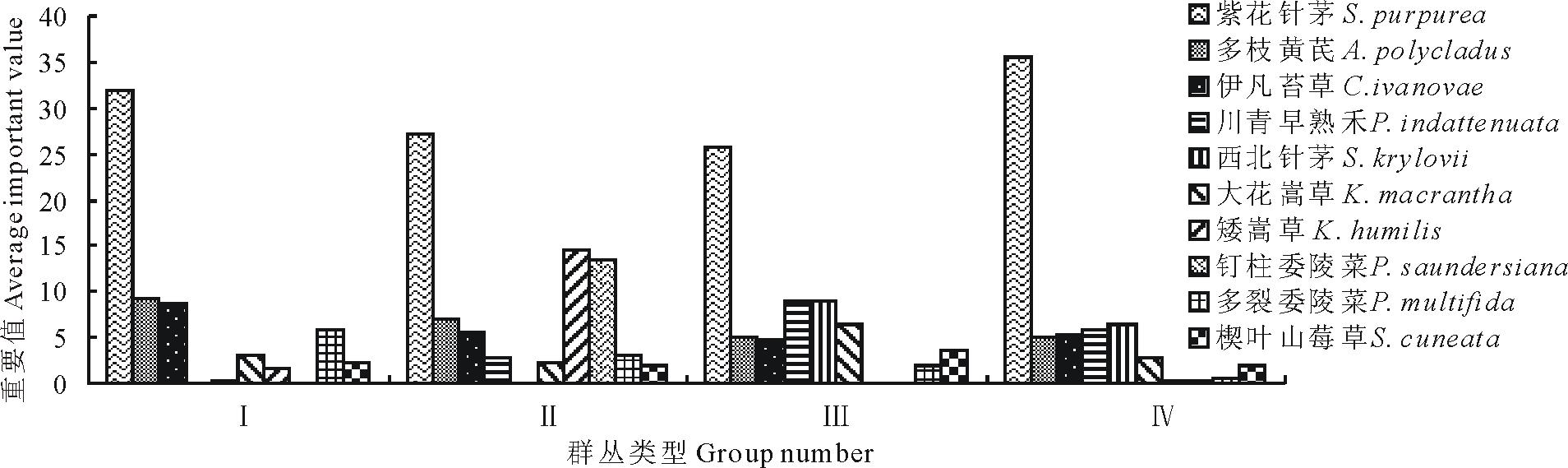 | 图3 研究区内紫花针茅草原群丛主要种重要值的变化Fig.3 Changes of important value of the S. purpurea steppe dominant species |
2.3 紫花针茅草原群丛分布与环境因子的关系
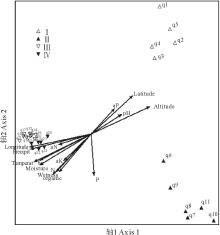
样带及环境因子的DCA和CCA排序结果反映了一致的规律。两种排序方法第1轴和第2轴的特征值都远高于第3轴(表4), 反映了绝大部分的变量信息。排序第1轴和经度、年均降雨量、海拔、年均温相关系数高, 主要反映了空间和气候因子的变化。第2轴和年均湿润度、土壤有机质, 纬度、土壤全氮磷含量、土壤水分相关系数较高, 主要反映了水分和土壤因子的变化。土壤速效磷、速效钾等与各排序轴相关性都很小。数量分类的4个群丛在DCA和CCA排序二维空间均被分开(图4, 5), 进一步证明了数量分类的合理性。排序第1轴很好的指示了群丛Ⅰ 、Ⅱ 和群丛Ⅲ 、Ⅳ 的区别。沿着第一排序轴从左向右, 经度逐渐变小, 降雨量减少, 海拔升高, 温度降低, 群丛类型从东部刚察县低海拔典型草原群丛Ⅲ 和Ⅳ 过渡到西部天峻县高海拔草甸化草原群丛Ⅰ 和Ⅱ 。沿着DCA第2排序轴从下到上, 湿润度逐渐减小, 土壤养分减少, 纬度增大, 群丛类型从分布在较湿润肥沃环境的茂密群丛Ⅱ 过渡到分布在较干旱贫瘠环境的稀疏群丛Ⅰ 。群丛Ⅲ 、Ⅳ 的区分相对不明显, 但仍然可以看出群丛Ⅲ 的湿润度和土壤营养元素低于群丛Ⅳ 。
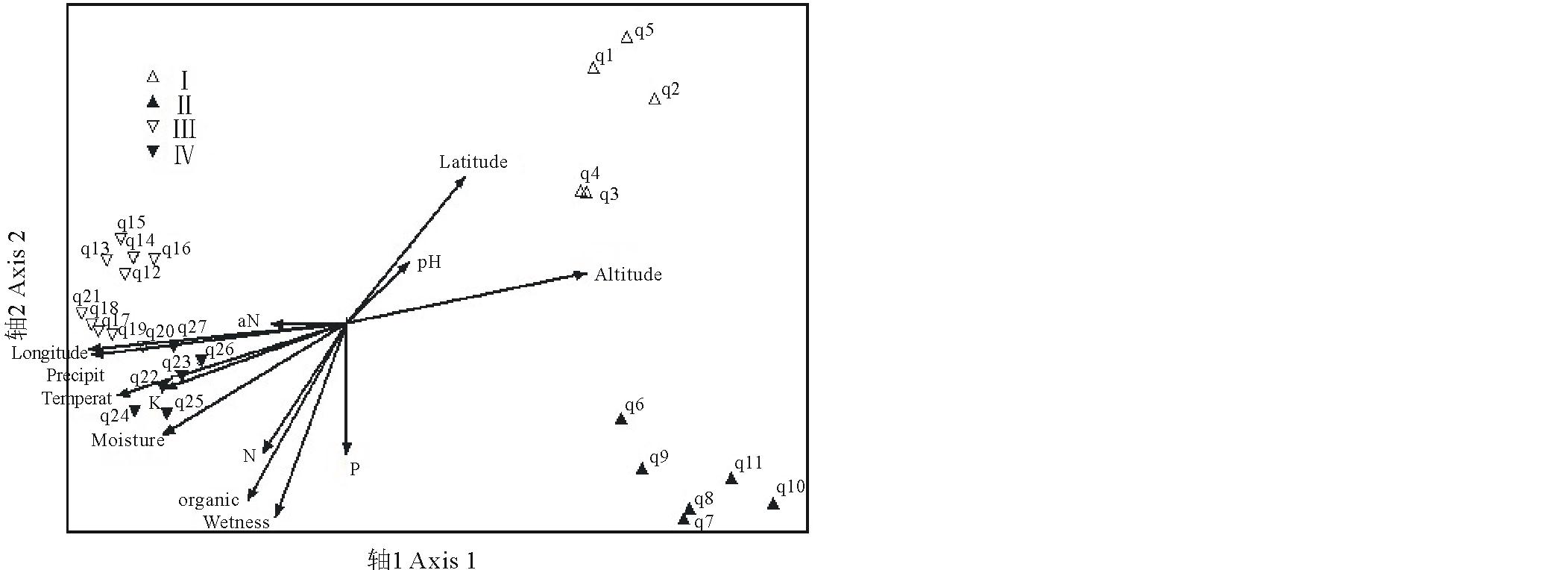 | 图4 研究区紫花针茅草原样带及环境因子的DCA排序图Fig.4 The DCA ordination diagram of transects and environmental factors of S. purpurea steppe Altitude, 海拔; Longitude, 经度; Latitude, 纬度; Temperat, 年均温度Average annual temperature; Wetness, 年均湿润度Degree of wetness; Precipit, 年均降雨量Average annual precipitation; pH, 土壤pH值 Soil pH; N, 土壤全氮含量 Soil N; aN, 土壤速效氮含量 Soil available N; P, 土壤全磷含量 Soil P; aP, 土壤速效磷含量 Soil available P; K, 土壤全钾含量 Soil K; aK, 土壤速效钾含量 Soil available K; organic, 土壤有机质含量 Soil organic content; Moisture, 土壤水分 Soil moisture; q1~q27指样号, Ⅰ ~Ⅳ 表示群丛类型。q1~q27 mean trinsect numbers. Ⅰ ~Ⅳ indicate the ecological association. 下同 The same below. |
2.4 α 多样性
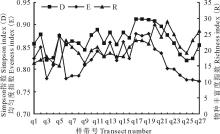
图6显示了研究区紫花针茅草原27条样带的Simpson指数(D)、均匀度指数(E)和物种丰富度指数(R)的变化曲线。总体来看, Simpson指数(D)和均匀度指数(E)的变化规律较相似, 物种丰富度指数和前两者相比波动较大。各群丛多样性平均值比较来看(表5), 均匀度指数变化趋势为群落Ⅲ > 群丛Ⅰ > 群丛Ⅱ > 群丛Ⅳ , Simpson指数的变化趋势为群落Ⅲ > 群丛Ⅱ > 群丛Ⅰ > 群丛Ⅳ 。群丛Ⅲ 均匀度指数和Simpson指数在各群丛中都是最高的。
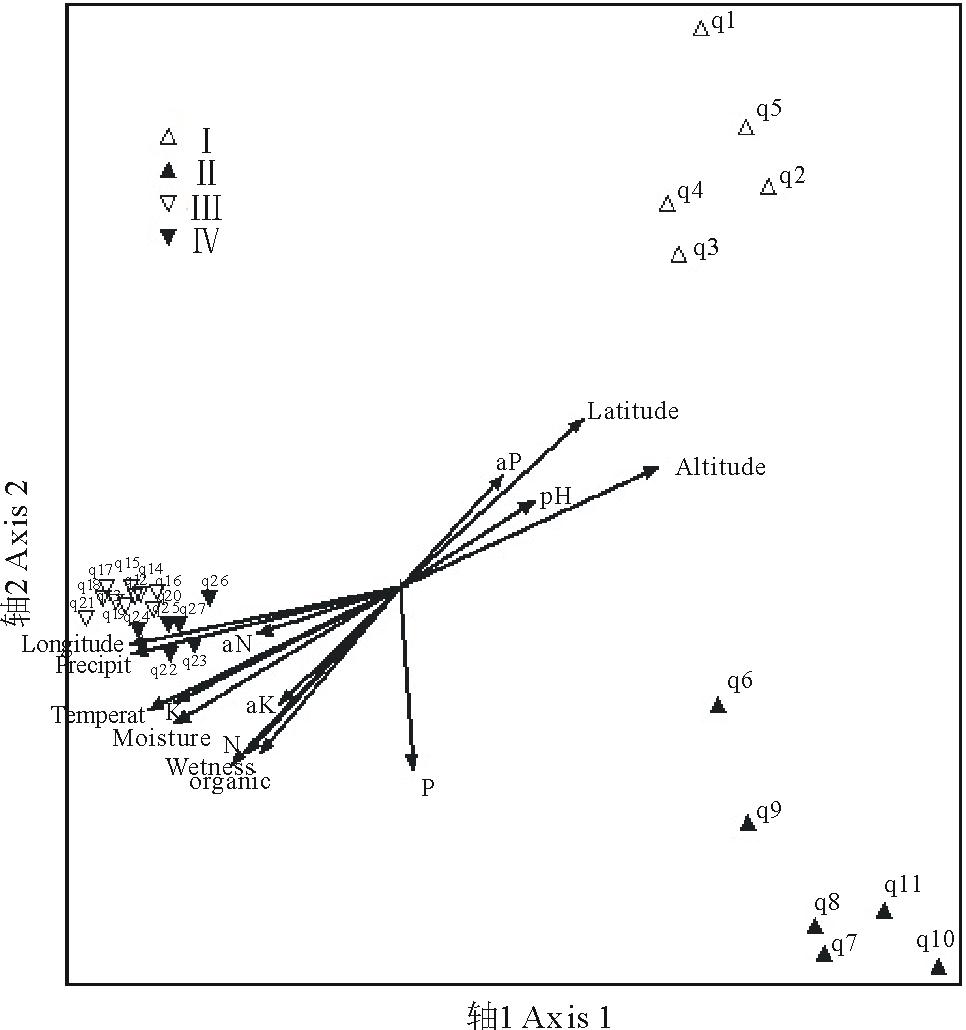 | 图5 研究区紫花针茅草原样带及环境因子的CCA排序图Fig.5 The CCA ordination diagram of transects and environmental factors of S. purpurea steppe |
通过SPSS计算, 得出了3个α 多样性指数和各环境参数间的相关系数(表5)。结果显示, 物种丰富度指数和海拔、经纬度、均温、平均降雨量、湿润度以及大部分的土壤因子均达到了显著相关, Simpson指数和均匀度指数却与所调查的所有环境因子都没有显著相关性。
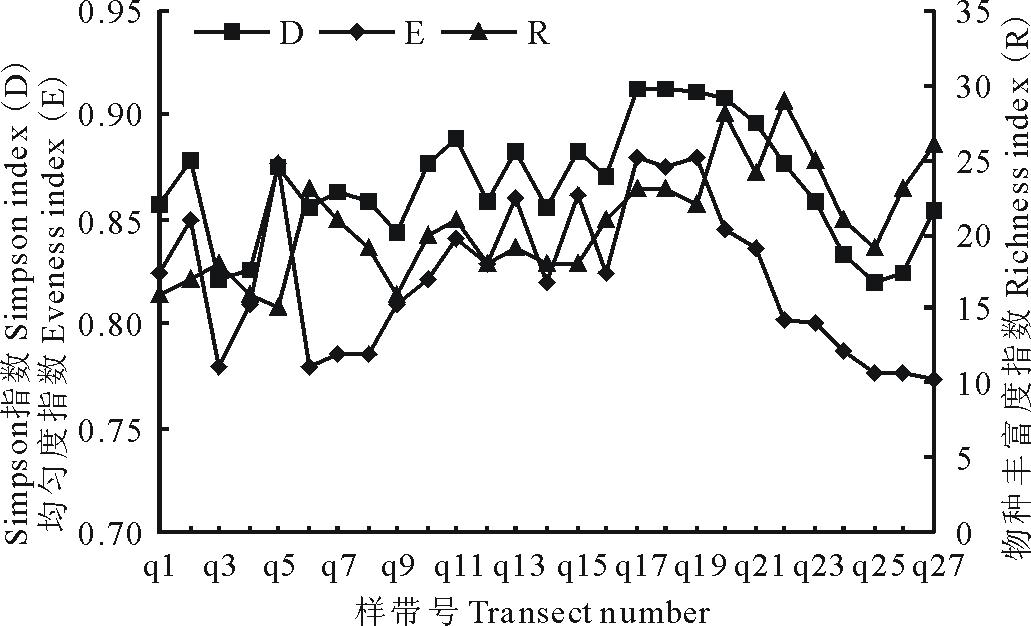 | 图6 研究区紫花针茅草原群落丰富度指数、均匀度指数和Simpson指数变化曲线Fig.6 The curves of richness, evenness and Simpson indices of S. purpurea steppe |
3 讨论
排序结果显示影响群丛分布的主要环境因子首先是空间和气候因子。鉴于青海湖北岸紫花针茅草原连续分布无显著地理隔离, 空间因子最终反映了温度和水分的变化, 例如随着海拔升高, 年均降雨量和年均温均下降等, 空间因子和气候因子在该研究区具有同维性。故最终影响紫花针茅草原群丛分布格局的主要环境因子归结为以年均降雨量和年均温为代表的气候因子。土壤因子对群丛分布的影响排在其次, 以土壤水分、有机质和全氮磷钾为代表。多枝黄芪和弱小火绒草分布于寒旱、高海拔区, 以多枝黄芪为次优势种的群丛Ⅰ 位于排序图右上端, 干旱、寒冷、海拔高, 和群丛分布区样地1的实际情况相符。群丛Ⅱ 和群丛Ⅰ 海拔都较高, 但群丛Ⅱ 以矮嵩草为次优势种, 伴生种中钉柱委陵菜和短穗兔儿草等湿润区分布的物种重要值大, 位于排序轴的右下端, 和分布区样地2较样地1降雨量大、湿润的实际情况相符。群丛Ⅲ 和群丛Ⅳ 都分布于刚察县, 气候环境相似, 伴生种相似, 在排序图中距离也较近。群丛Ⅲ 以川青早熟禾为次优势种, 总盖度低于群丛Ⅳ , 和排序图中左上端较干旱、土壤有机质少的位置相符合。本文中青海湖北岸紫花针茅草原群落27条样带通过两种数量分类方法划分为4个群丛类型, 各群丛类型在DCA和CCA排序图中也得到了较好的区分, 环境因子对群丛分布格局解释充分。
| 表5 α 多样性指数和环境因子间的相关分析 Table 5 The correlation analysis among α diversity indexes and environmental factors |
表6研究表明, 生物多样性具有重要的生态系统功能[13, 14, 15], 其中α 多样性, 或者说物种多样性的研究尤为引人注目, 它是一个群落结构和功能复杂性的度量。本文在实地调查的基础上初步分析了青海湖地区紫花针茅草原群落α 多样性。群丛Ⅲ 紫花针茅重要值相对最小, 主要伴生种的重要值相对最均匀, 故该群丛均匀度指数和Simpson指数最大。群落α 多样性分布格局不同于群落分布格局, 影响两者的环境因子可能不同。肖绪培等[16]研究发现土壤水分是影响植被生长的关键生态因子, 但它对物种丰富度影响较小。李愈哲等[17]研究发现长时间的围封显著增加群落的地上地下生产力, 但对群落物种组成和物种多样性影响不大。王建兵等[18]研究也表明草甸退化引起植物物种的变化。本文中物种丰富度指数和海拔、经纬度、年均温、年均降雨量、湿润度以及大部分的土壤因子均达到了显著相关, 说明此研究区物种丰富度可能受空间、气候和土壤因子的影响很大, 而这些影响恰好在此处具有良好的同维性。不同于群丛分布和环境因子的高相关系数, Simpson指数和均匀度指数与本文所调查的所有环境因子都没有显著相关性, 原因可能是这两个反映物种组成和比例的多样性指数受多环境因子协同作用的调节, 和单一环境因子的相关性反而较小。此外, 青海湖地区紫花针茅草原群落物种多样性分布特点与江河源区紫花针茅草原群落不尽相同[19], 可能是研究区之间存在明显的地形地势差异、各区主要限制因子不同所致。这也反映了多元环境因子相互作用的复杂性, 群落特征在不同的环境下表现各自不同特点, 而二者之间的关系和作用机理更是复杂多变, 需要更深入和系统的研究, 以揭示青藏高原高寒草原的演替与环境因子的相互作用。
面对众多的数量分类方法, 为了使数据分析更加客观合理, 研究者有时使用两种或几种方法对同一组数据进行分类, 比较结果; 有时使用同一类方法, 而选用不同数据类型来验证矫正[20]。本文选择了TWINSPAN和等级聚类两种数量分类方法, 得到较好的相互验证和矫正作用, 使紫花针茅草原群落分类更符合实际生态学意义。在现有的数量排序方法中, DCA与高斯的群落模型最为吻合, 是植被分析中最为有效的一种方法, 也是20世纪80年代最常用的排序方法。一般情况下, 如果同分类方法结合使用, DCA的效果要好于CCA[21]。但CCA的应用突出反映了物种与环境的相关性, 有些学者的研究发现, DCA和CCA方法的结合使用有时比单独使用的效果好[22, 23]。本文中DCA排序的结果和CCA排序结果基本一致, 一方面显示了排序的合理性, 客观反映了群丛分布和环境的关系; 另一方面又和数量分类的相互验证效果良好, 与同类研究结果一致。在紫花针茅草原群落数量分类和排序中, TWINSPAN、等级聚类和DCA、CCA排序均较适用, 综合验证效果好。植物群落的各种参数都是某一方面特征的反映, 它们是相关相补的。只有有效运用合理的数量分析方法, 客观反映紫花针茅草原群落的共性和各区域不同群丛的差异, 才能有针对性地提出保护和管理对策, 为江河源区生态环境保护提供科学根据。
4 结论
1)研究区内所调查的东西向连续分布紫花针茅草原群落出现种子植物58种, 隶属45属, 21科。其物种组成简单, 北温带分布的属数量最多。
2)紫花针茅草原可细分为4个群丛类型:紫花针茅+多枝黄芪群丛(Ⅰ ), 紫花针茅+矮嵩草群丛(Ⅱ ), 紫花针茅+西北针茅群丛(Ⅲ )和紫花针茅+川青早熟禾群丛(Ⅳ )。影响紫花针茅草原群丛类型分布的主要环境因子首先是以年均降雨量和年均温为代表的气候因子, 其次是土壤水分、有机质和全氮磷钾等土壤因子。数量分类和排序的相互验证性良好。
3)研究区东西向分布的紫花针茅草原群落均匀度指数和Simpson指数的变化趋势基本一致, 紫花针茅+川青早熟禾群丛均匀度指数和Simpson指数是最高的。物种丰富度受气候和土壤因子的影响大, 但Simpson指数和均匀度指数与所调查的所有环境因子都没有显著相关性。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|