{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
紫茎泽兰潜在分布对气候变化响应的研究
引用本文
王翀, 林慧龙, 何兰, 曹坳程. 紫茎泽兰潜在分布对气候变化响应的研究. 23(4): 20-30
WANG Chong, LIN Hui-long, HE Lan, CAO Ao-cheng. Research on responses of Eupatorium adenophorum’s potential distribution to climate change . Acta Prataculturae Sinica, 23(4): 20-30
Permissions
WANG Chong, LIN Hui-long, HE Lan, CAO Ao-cheng. Research on responses of Eupatorium adenophorum’s potential distribution to climate change . Acta Prataculturae Sinica, 23(4): 20-30
Copyright©2014, 草业学报编辑部
本文属于开放获取期刊。
紫茎泽兰潜在分布对气候变化响应的研究
作者简介:王翀(1987-),女,甘肃白银人,在读博士。E-mail:anne_cat2008@163.com
摘要
紫茎泽兰的生态适应性强,是我国外来入侵物种中危害最为严重的恶性杂草。研究紫茎泽兰的适生性特征及对全球气候变化的响应规律是制定防控策略的重要基础,为此,本研究采用了将生态位因子分析(ENFA)与最大熵模型(MaxEnt)嵌套的方法,首先通过ENFA对环境因子进行降维,利用降维后的环境因子以及当前及A1b情景的未来气候数据,根据最大熵模型(MaxEnt)预测紫茎泽兰的潜在分布,并使用ROC曲线分析对预测结果进行评价。结果显示,当前气候情景下,紫茎泽兰的分布区以云南、贵州、广西等省为主;未来A1b情景下,易入侵等级(入侵概率为0.6~1.0)的区域面积将会由当前的12.82 km2增加至2080s的21.30 km2,中心点将由当前位置向西南方向移动61 km;而其中高入侵概率等级(入侵概率为0.8~1.0)的区域面积将由当前的0.42 km2增加至2080s的0.91 km2, 中心点将由当前位置向东南方向移动178.66 km。根据当前及未来A1b气候情景下紫茎泽兰潜在分布情况,并根据不同入侵等级区域采取相应的防除治理措施,将对紫茎泽兰的综合治理具有重要指导意义。
关键词:
物种入侵; 紫茎泽兰; 生态位模型; 生态位因子分析(ENFA); 最大熵模型(MaxEnt)
中图分类号:S812.29
文献标志码:A
文章编号:1004-5759(2014)04-0020-11
Research on responses of Eupatorium adenophorum’s potential distribution to climate change
Abstract
Eupatorium adenophorum is one of the most invasive alien species in China. It has a strong ecological adaptability, and it can seize the niche very quickly thus the native species would be posed a threat to biodiversity by being crowded out. In order to study the potential distribution of E. adenophorum and its response to climate change, also to find out the environmental factors which influence the distribution of E. adenophorum, the ecological niche factor analysis (ENFA) and the maximum entropy model were combined in this study. Firstly the environmental factors dimension was reduced by ENFA; secondly together with current and future climate data the study use MaxEnt model to predict the distribution of E. adenophorum in the future. The results showed that in the A1b scenario, the main distribution areas were Yunnan, Guizhou, and Guangxi province. From now to 2080s, the area of L45 grade (the invasion probability is 0.6-1.0) was increased from 12.82 km2 of nowadays to 21.30 km2 in 2080s, and the center moved 46.62 km northwest. While the area of L5 grade (the invasion probability is 0.8-1.0) was increased from 0.42 km2 of nowadays to 0.91 km2 in 2080s, and the center moved 178.66 km northeast. According to the research of the potential distribution of E. adenophorum in the current and future, the area of different invasion grade could be controlled by appropriate measures, and it will provide an important guidance for the comprehensive management.
Keyword:
invasive species; Eupatorium adenophorum ; niche model; ecological niche factor analysis (ENFA); maximum entropy model (MaxEnt)
引言
紫茎泽兰(Eupatorium adenophorum)是菊科(Compositae)泽兰属(Eupatorium)的多年生丛生型半灌木状草本植物, 原产于中美洲的墨西哥和哥斯达黎加等国, 19 世纪曾作为观赏植物引种到欧洲, 后来引种到澳洲和亚洲, 是一种抗逆性强、生态适应广、传播速度快且群体自然繁殖与演替能力极强的植物。20世纪40 年代从中缅边境传入西双版纳后, 以平均20 km/a的速度向东、向北快速蔓延, 目前其入侵面积超过3000万hm2。紫茎泽兰是我国外来入侵物种中危害最为严重的植物之一, 据估计, 它对中国畜牧业和草原生态系统服务功能造成的损失分别为9.89 和26.25 亿元/h
关于生态位的定义虽然为数不少, 但最具代表性的当推Grinnell[6, 7, 8]、Elton[9]和Hutchinson[10]三人, 后人分别称他们对生态位的定义为“ 空间生态位” 、“ 功能生态位” 和“ 多维超体积生态位” 。其中Hutchinson(1957)[10]认为, 生物在环境中受着多个资源因子的供应和限制, 每个因子对该物种都有一定的适合度阈值, 在所有这些阈值所限定的区域内, 任何一点所构成的环境资源组合状态上, 该物种均可以生存繁衍, 所有这些状态组合点共同构成了该物种在该环境中的多维超体积生态位。多维超体积概念为现代生态位理论研究奠定了基础[11]。
生态位模型是根据生态位理论进行的建模。根据是否需要物种分布信息可将生态位模型分为两类, 即关联模型和机理模型, 其中关联模型需要物种分布信息, 而机理模型(代表模型有CLIMEX、SDM等)则不需要。根据物种分布数据的不同, 又可将关联模型分为组判别模型和框架模型。组判别模型需要知道“ 存在” 数据和“ 不存在” 数据, 这类模型的目的就是要找出一个能将这两个类别正确分开的判别规则, 即建立一个合适的分类器, GARP和MaxEnt即为这类模型; 而框架模型只需要知道物种“ 存在” 的分布数据, 这类模型主要是从这些物种分布信息中概括出该物种所需的生态位需求, 主要代表有BIOCLIM、DOMAIN、ENFA等[12, 13]。
最大熵模型(MaxEnt)以最大熵理论为基础, 是一个物种密度估计和物种分布预测模型。该模型的最大特点在于能够预测物种在某个地点存在的概率, 而非是否存在。许多学者的研究已经证明, MaxEnt不仅能够很好的预测物种分布, 并且模型运行速度快[12, 13, 14, 15]。在使用MaxEnt模型进行较大空间尺度的物种潜在分布预测时, 由于选取的环境因子数目较多, 会大大降低MaxEnt模型的运行效率, 此时就需要对环境因子进行筛选(或降维)[12]。为了实现这一目的, 可选取生态位因子分析(ENFA)和主成分分析(PCA)两种方法。两种方法的原理相似, 但PCA得出的只是反映环境变量本身变异程度的因子, 并不考虑物种分布信息, 而ENFA考虑了物种分布信息, 使其选取的环境因子更具有生态学意义, 所以本研究选取ENFA对环境变量进行降维。
本文拟利用生态位原理, 将生态位因子分析(ENFA)与最大熵模型(MaxEnt)进行嵌套, 找出影响紫茎泽兰分布的10个主要环境因子, 并结合地形数据和当前及未来A1b情景(即中等碳排放水平[16])下的气象数据, 预测紫茎泽兰在中国的潜在分布情况, 为不同入侵等级的区域制定相应的防控策略提供必要条件。
1 材料与方法
1.1 数据来源
1.1.1 紫茎泽兰分布数据
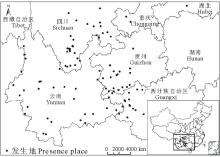
本研究共收集全国紫茎泽兰发生地数据106个(图1), 其中部分来自于全球生物多样性信息交换所GBIF数据库(http://data.gbif.org/welcome.htm), 其他数据来源于已发表的文献及出版物[17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32]。根据收集到的数据, 紫茎泽兰目前主要分布在云南、贵州、广西、四川等省, 西藏、湖北、台湾等地区也有零星分布。
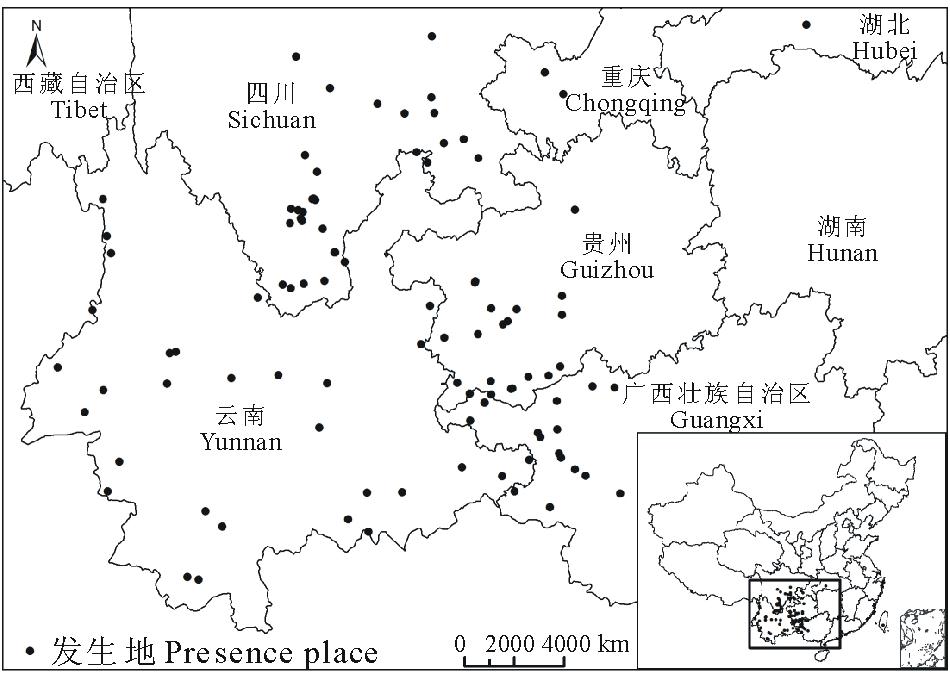 | 图1 中国紫茎泽兰发生地示意图Fig.1 The schematic of E. adenophorum occurred in China |
1.1.2 环境因子数据
为研究影响紫茎泽兰分布的环境因子, 本研究收集了包括高程、生物气候数据、月均温和月降水等共计46个环境因子数据, 其中44个环境因子数据依据WorldClim(http://www.worldclim.org)提供的当前(1961-1990年)和加拿大气候模型与分析中心(Canadian Centre for Climate Modelling and Analysis, CCCMA)发布的A1b情景下的未来(2010-2099年)月降水、温度数据计算得到(表1)。坡度和坡向数据根据高程数据计算得到。
| 表1 环境变量及其代表的意义 Table 1 Content information of bioclimatics data |
1.1.3 行政区划数据
从国家基础地理信息系统(http://nfgis.nsdi.gov.cn/)下载获得1∶ 400 万的中国地图作为分析底图。
1.1.4 相关软件
训练样本和测试样本抽取使用DIVA-GIS 软件进行, 而根据样本数据和环境因子变量预测物种潜在分布则使用MaxEnt软件进行。该软件可在线免费获取(http://www.cs.princeton.edu/~schapire/MaxEnt/), 版本为3.3.3。生态位因子分析(ENFA)是Biomapper软件的主要模块, Biomapper软件可以从http://www2.unil.ch/biomapper/下载, 版本为4.0; 本研究在进行生态位因子分析前还需要对环境因子变量进行格式转换, 可使用ArcGIS的一个模块完成(从http://www.pierssen.com/idrisi/grid.htm下载); GIS相关分析和地图制作使用ESRI公司的ArcGIS 9.3软件。
1.2 研究方法
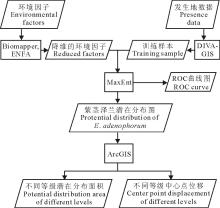
1.2.1 训练样本和测试样本选取 本研究的数据处理流程见图2。通过DIVA-GIS软件选取全部样点的75%作为训练样本, 其余样点和随机选取10倍于物种分布点的背景点共同作为测试样本。训练样本数据用于预测物种分布情况, 测试样本数据用于检验预测结果的可靠性。
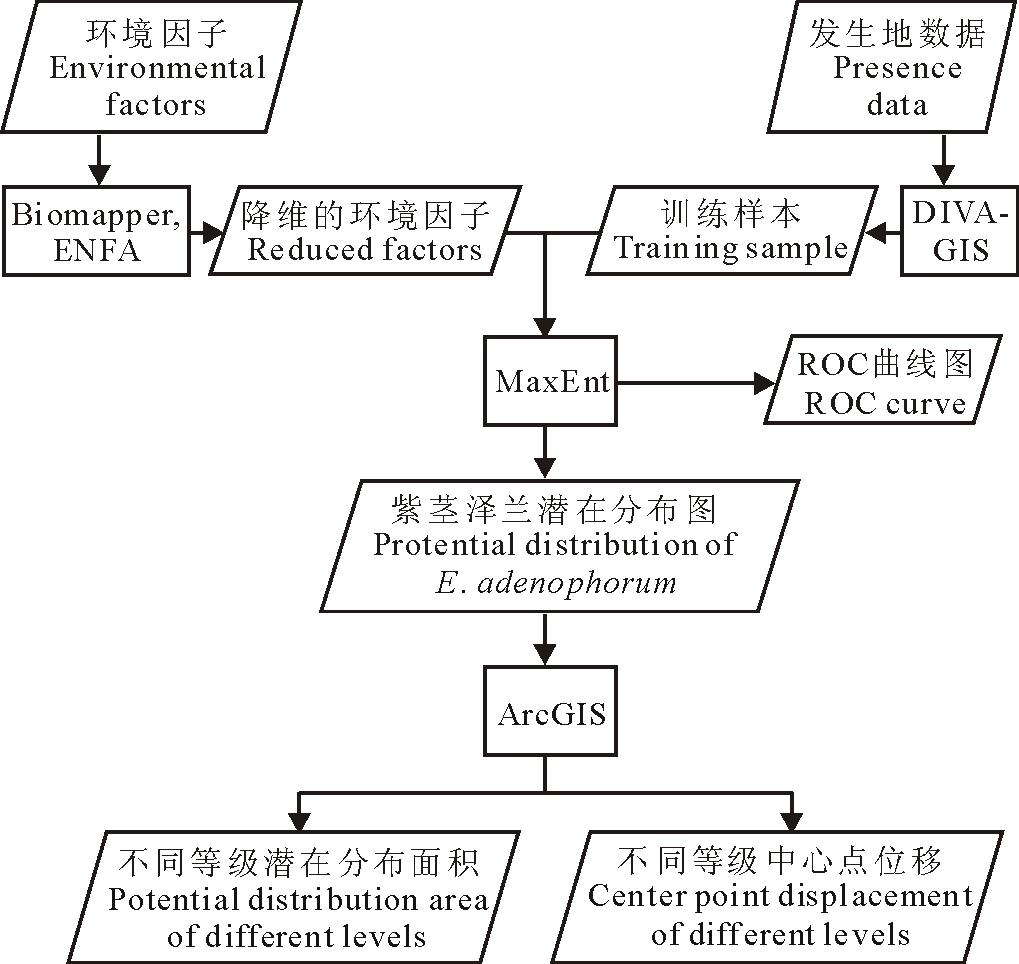 | 图2 数据处理流程图Fig.2 The flowchart of data processing |
1.2.2 环境变量降维
由于本研究中选取了46个环境变量因子, 如果全部参与计算会导致计算量大、耗时长, 因此需要对环境变量做降维处理。
在本研究中, 利用当前和未来气候数据对紫茎泽兰入侵概率的模拟均以迄今为止紫茎泽兰的发生地作为存在点。在此前提下, 利用ENFA对环境变量进行降维, 并最终选取得分矩阵中排名前十的环境因子进行物种分布预测的计算。
1.2.3 物种潜在分布预测及模拟结果评价
将训练样本数据和10个环境变量作为输入数据, 采用MaxEnt模型生成物种分布预测图、受试者工作特征曲线图(receiver operating characteristic curve, 即ROC曲线图)。ROC曲线图以假阳性率为横坐标, 灵敏度为纵坐标, ROC曲线的曲线下面积(AUC)可用于评价模型的预测能力[14, 33]。AUC 取值范围为0~1, 值越大代表模型的预测越准确。一般认为, AUC 值为0.5~0.7 时诊断价值较低, 为0.7~0.9时诊断价值中等, 大于0.9 时诊断价值较高。本研究将使用ROC曲线分析对MaxEnt模型的预测结果进行评价。最后根据物种潜在分布情况, 将紫茎泽兰的入侵概率分为5个等级, 各入侵等级分别代表不发生入侵(L1)、低入侵概率等级(L2)、中等入侵概率等级(L3)、较高入侵概率等级(L4)、极高入侵概率等级(L5)。分别统计各等级在未来不同时期的面积变化情况以及各等级中心点的位移。计算中心点位移的具体方法参见Hart[34]和Yue等[35]的研究。2 结果与分析2.1 环境变量的降维结果
根据ENFA降维后筛选出的10个环境因子可知, 影响紫茎泽兰分布的主要限制因子为冬季温度, 其次为暖季(主要为8月和10月)月降水。地形因素对紫茎泽兰分布的影响不大(表2)。
| 表2 不同时期的主要环境因子 Table 2 Main environmental factors in different periods |
2.2 紫茎泽兰潜在分布预测结果评价
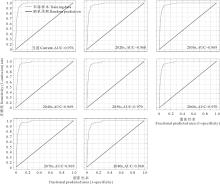
由图3可知, 通过MaxEnt模型得到的紫茎泽兰物种分布预测结果的AUC值都大于0.9, 预测结果很好, 对紫茎泽兰潜在分布区的预测较为可信。
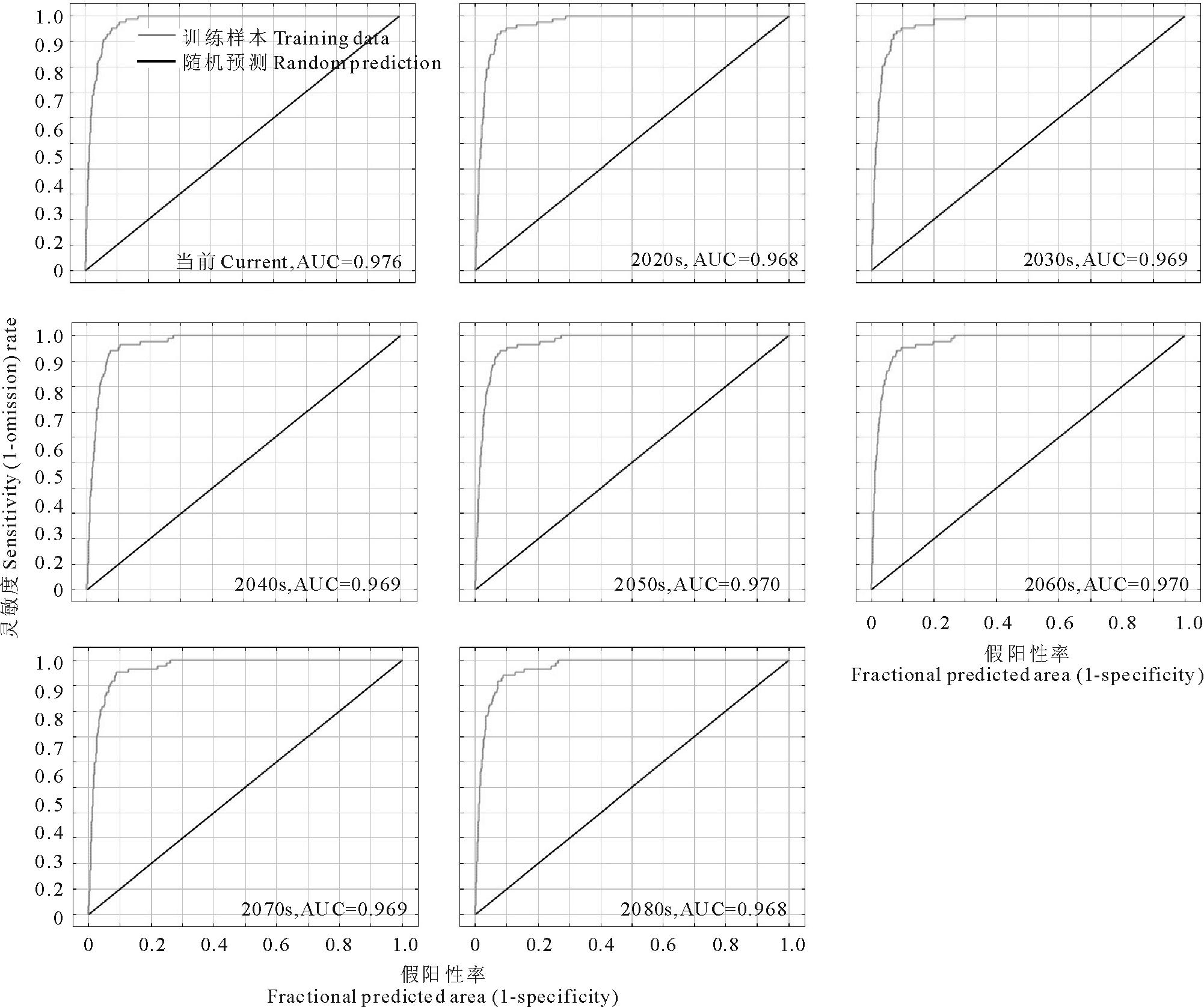 | 图3 ROC曲线分析及AUC值Fig.3 ROC curve and AUC value |
2.3 紫茎泽兰潜在分布
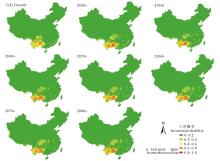
通过MaxEnt模型对紫茎泽兰潜在分布的模拟可知(图4), A1b情景下未来紫茎泽兰的入侵区仍主要集中在云南、四川、广西、贵州等省, 但存在向江西、湖北、湖南、安徽等省扩散的趋势。
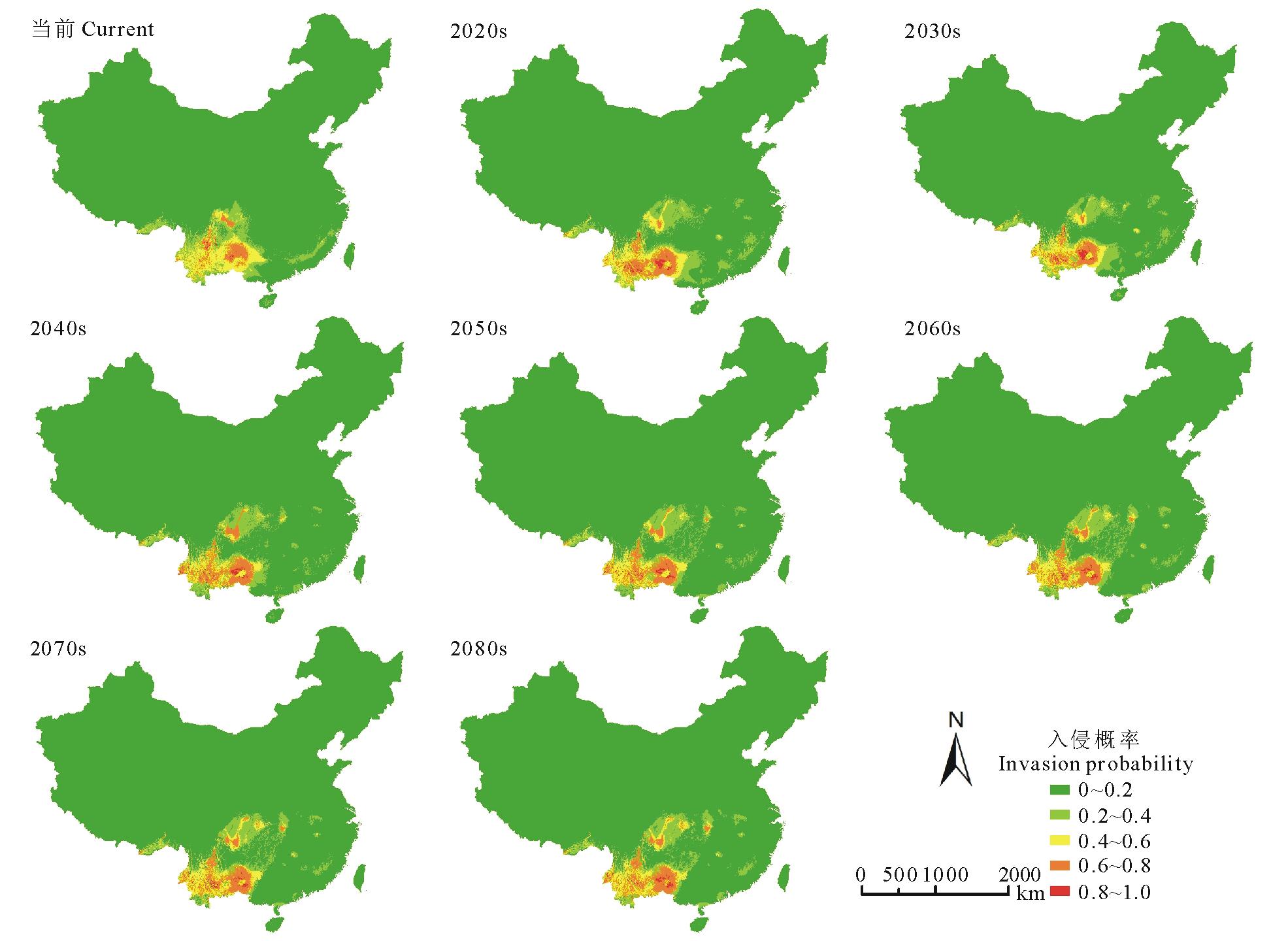 | 图4 当前及未来情境下紫茎泽兰的潜在分布区Fig.4 Potential distribution of E. adenophorum in the current and future in China |
2.4 不同时期不同入侵等级的面积变化
将入侵概率分为5个等级, 分别表示为L1~L5。其中L1和L2(即入侵概率分别为0~0.2, 0.2~0.4)为不发生入侵或入侵概率低, 在此不做讨论, L3~L5等级对应的入侵概率为0.4~0.6, 0.6~0.8, 0.8~1.0。将L3~L5等级合并为可入侵等级L345, L4和L5合并为易入侵等级L45。本研究中将根据Hart[34]和Yue等[35]的研究方法, 结合GIS技术手段分别研究不同入侵等级的面积变化和中心点位移情况。
2.4.1 L3面积变化及中心点位移
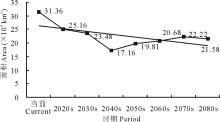
就L3等级而言, 在当前及未来A1b情景下, 其区域面积呈现出先减少后增加的变化趋势。总体上由当前的33.36 km2减少至2080s的21.58 km2(图5)。
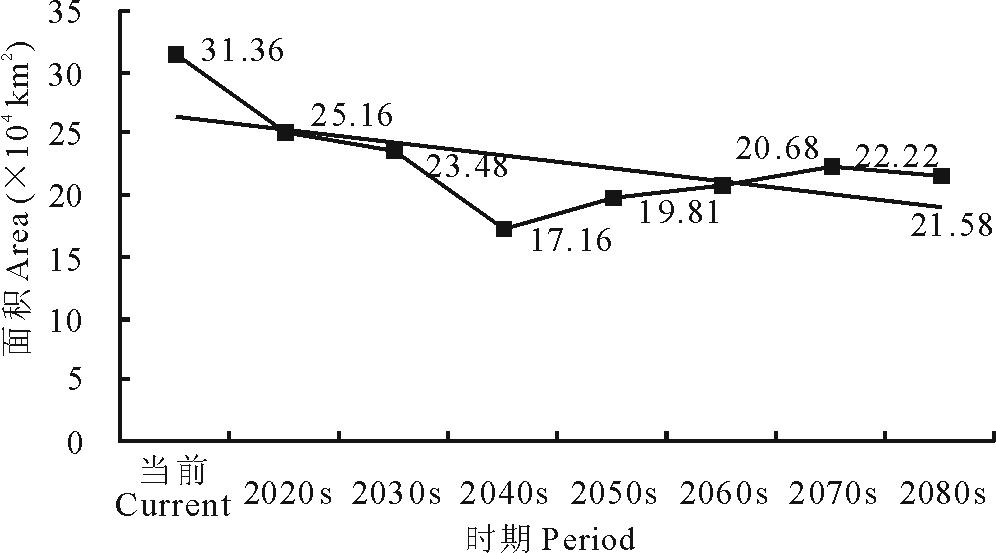 | 图5 不同时期L3面积变化Fig.5 Areas of L3 in different periods |
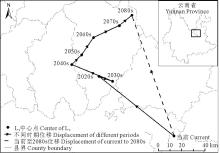
当前至2080s, L3区域的中心点由宜良县经嵩明县、寻甸彝族自治区、富民县、武定县, 最后移动至禄劝彝族苗族自治县, 总体上向西北方向移动了109.62 km, 并在2020s~2030s期间出现了往复(表3, 图6)。
| 表3 不同时期L3中心点位移 Table 3 Center displacement of L3 in different period |
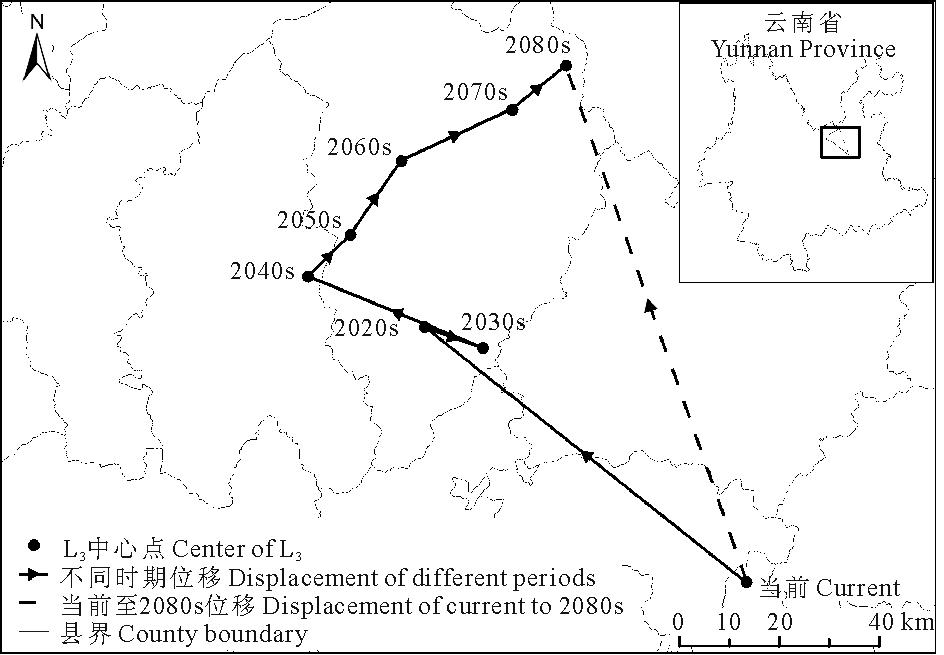 | 图6 不同时期L3中心点位移Fig.6 Center displacement of L3 in different periods |
2.4.2 L4面积变化及中心点位移
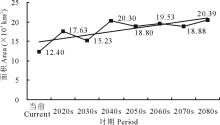
就L4等级而言, 在当前及未来A1b情景下, 其区域面积呈现出波动增加的变化趋势(图7)。总体上由当前的12.40 km2增加至2080s的20.39 km2。
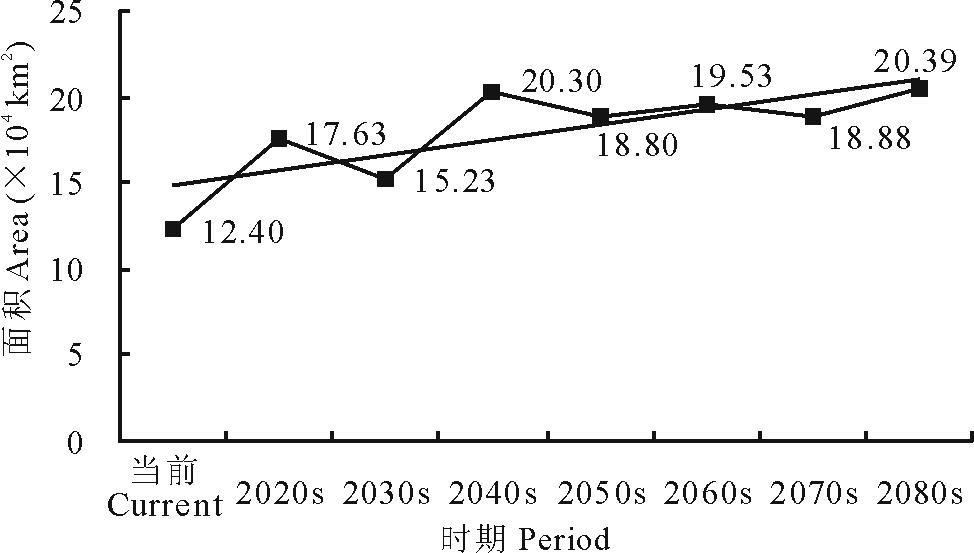 | 图7 不同时期L4面积变化Fig.7 Areas of L4 in different periods |
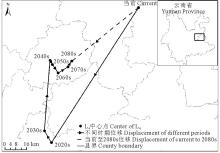
当前至2080s, L4区域的中心点由沾益县经曲靖市、陆良县、石林彝族自治县、宜良县至马龙县, 总体上由当前位置向西南方向移动了63.03 km(表4, 图8)。
| 表4 不同时期L4中心点位移 Table 4 Center displacement of L4 in different periods |
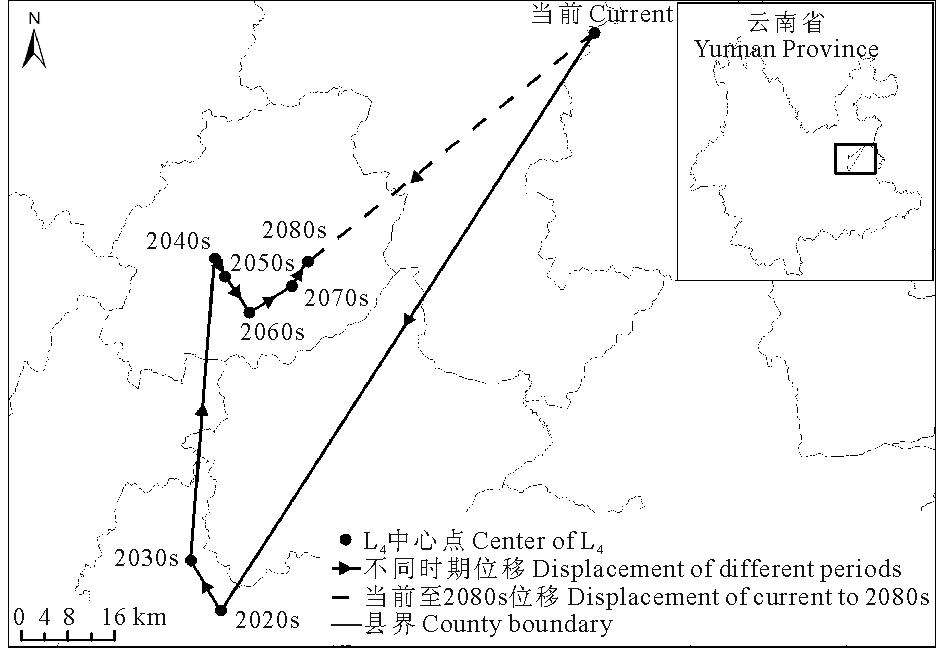 | 图8 不同时期L4中心点位移Fig.8 Center displacement of L4 in different periods |
2.4.3 L5面积变化及中心点位移
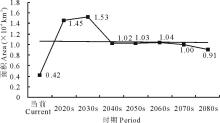
就L5等级而言, 在当前及未来A1b情景下, 其区域面积呈现出先增加后减少的变化趋势。总体上由当前的0.42 km2增加至2080s的0.91 km2(图9)。
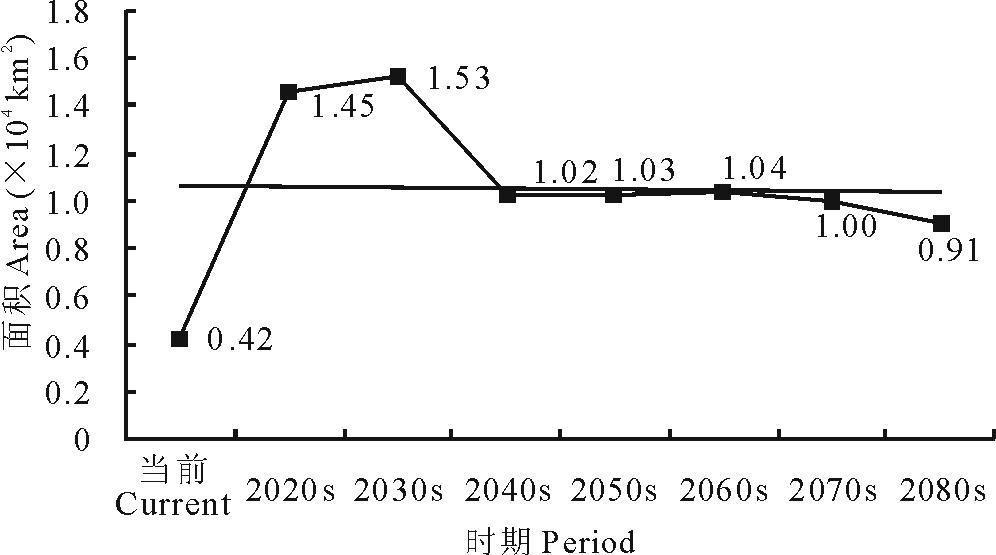 | 图9 不同时期L5面积变化Fig.9 Areas of L5 in different periods |
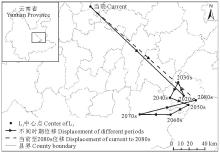
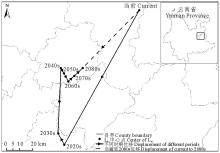
当前至2080s, L5区域的中心点在劝禄彝族自治县、曲嵩明县、宜良县、石林彝族自治县、泸西县、弥勒县内移动, 期间出现多次往复, 总体上由当前位置向东南方向移动了178.66 km(表5, 图10)。
| 表5 不同时期L5中心点位移 Table 5 Center displacement of L5 in different periods |
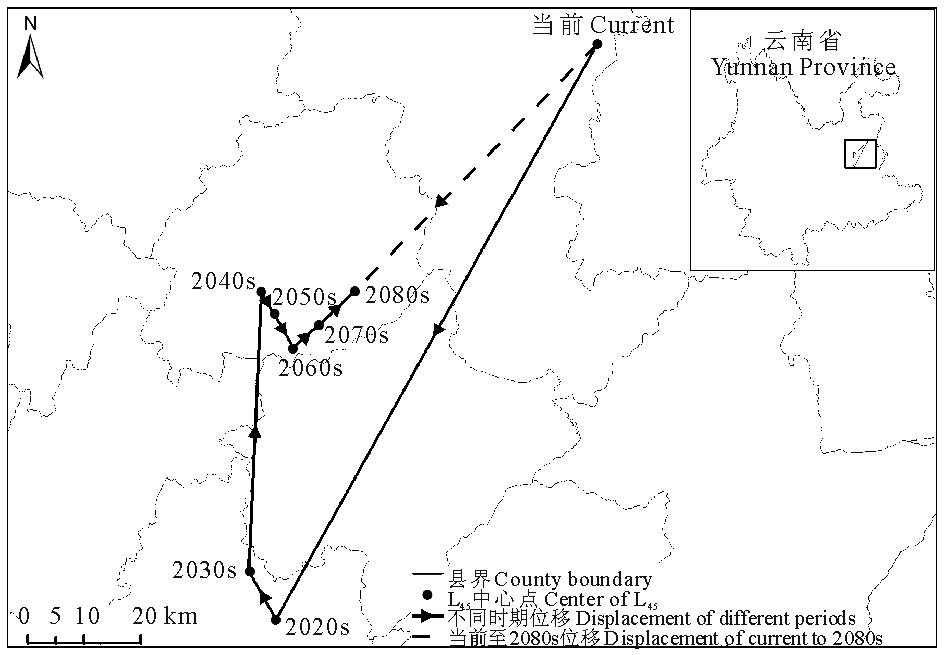
2.4.4 L345面积变化及中心点位移
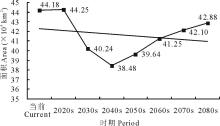
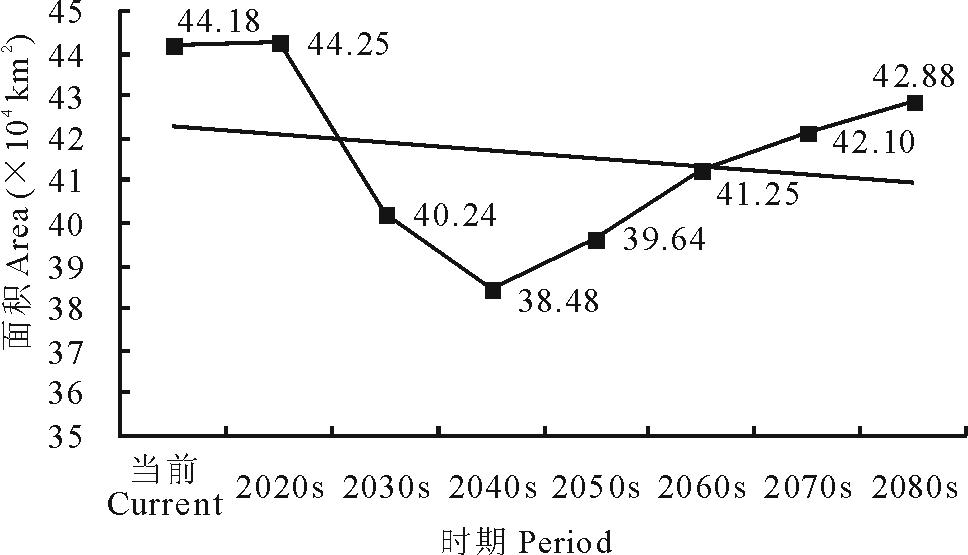
当前至2080s, 可入侵等级L345的面积呈现先减少后增加的变化趋势, 总体上由当前的44.18 km2减少至2080s的42.88 km2(图11)。
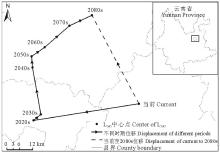
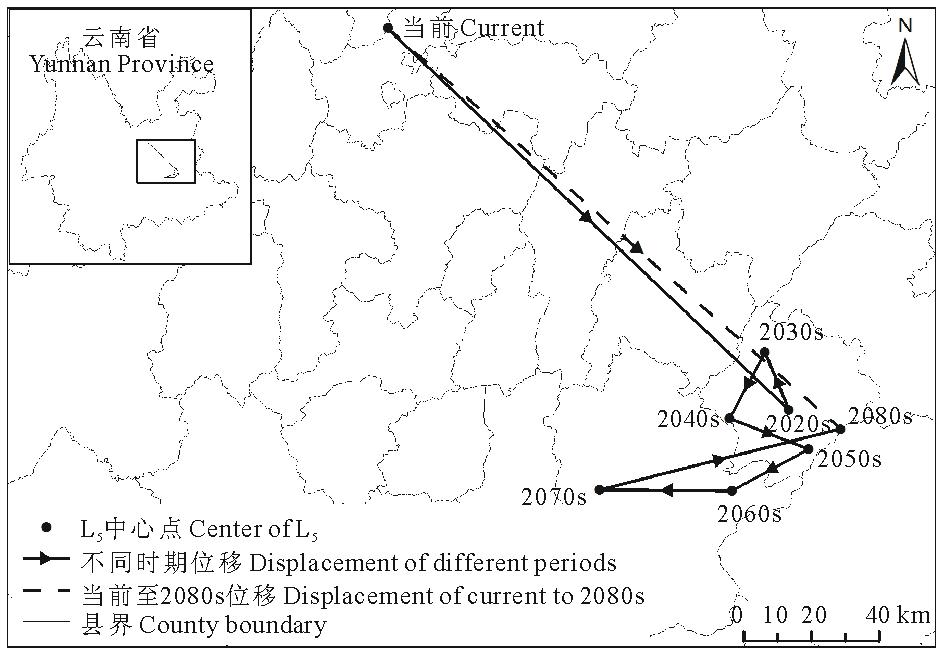
当前至2080s, L345的中心点经马龙县、嵩明县至寻甸彝族苗族自治县, 总体上向西北方向移动了46.62 km(表6, 图12)。
| 表6 不同时期L345中心点位移 Table 6 Center displacement of L345 in different periods |
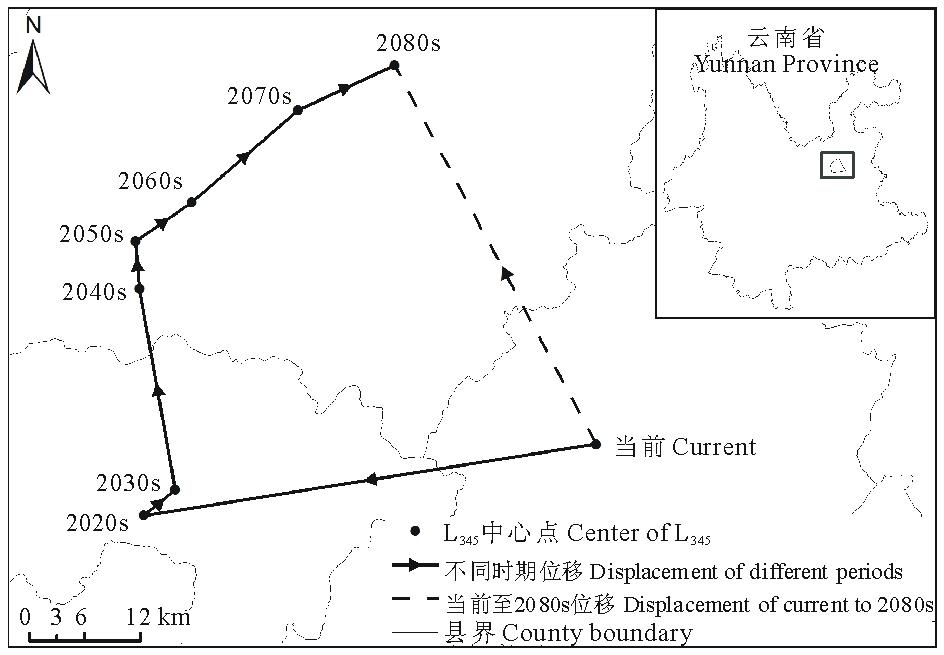
2.4.5 L45面积变化及中心点位移
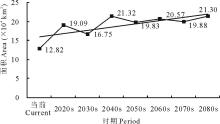
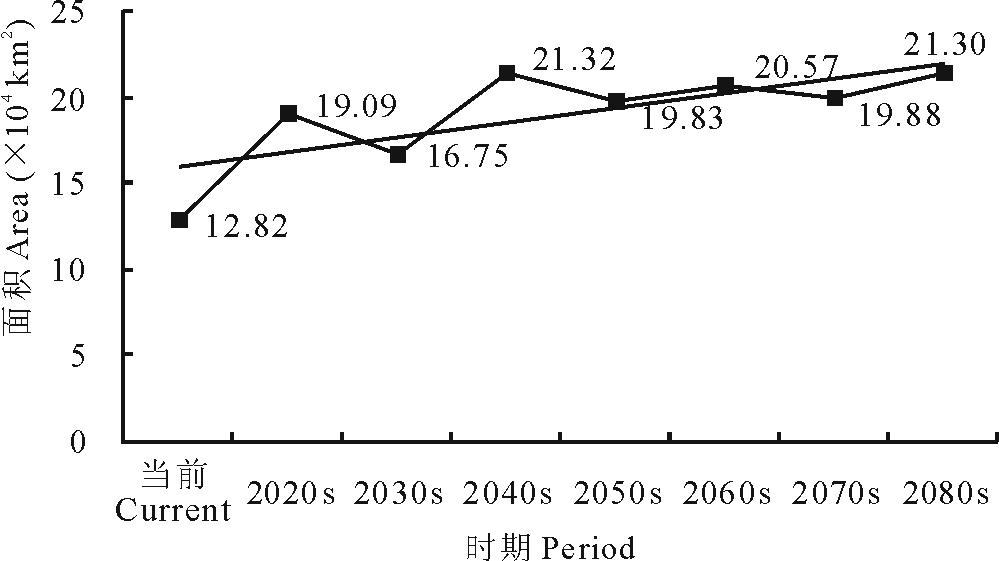
当前至2080s, 易入侵等级L45的面积呈现增加的趋势, 由当前的12.82 km2增加至2080s的21.30 km2(图13)。可见紫茎泽兰易入侵等级的区域范围在未来将会扩大。L45的中心点将由当前位置向西南方向移动61.38 km(图13)。
从以上结果可知, 当前至未来不同时期, 紫茎泽兰的主要入侵区依然集中在云南及周边各省。可入侵区域L345的面积虽然有所下降, 但是易入侵区域L45的面积增加了近1倍。可入侵等级L345的中心点向西北方向移动, 而易入侵等级L45的中心点则向西南方向移动(表7, 图14)。
| 表7 不同时期L45中心点位移 Table 7 Center displacement of L45 in different periods |
3 讨论
紫茎泽兰作为一种危害大, 入侵能力强的物种, 已经引起广泛关注[36, 37, 38, 39]。但目前相关研究多集中在对其入侵机理、生存习性以及防除治理等方面, 研究尺度也多为某个小范围的发生区域。比如王俊峰和冯玉龙[40]、李渊博等[41]、韩利红和冯玉龙[42]认为紫茎泽兰能够分泌化感物质使其能够成功入侵, 而于兴军等[43]认为化感物质并不是紫茎泽兰入侵的主要原因, 并认为入侵种改变了土壤细菌群落很可能造成紫茎泽兰能够成功入侵; 刘晓燕等[44]也认为紫茎泽兰在资源争夺中的优势使其成功入侵。然而利用生态位模型在更大的尺度上研究紫茎泽兰适生区, 并利用未来气候情景数据研究其入侵趋势, 此类研究还较为少见。
MaxEnt模型较其他生态位模型而言, 在预测物种分布方面获得了较好的效果。王运生[12]通过比较BIOCLIM、MaxEnt等6种生态位模型, 研究了胜红蓟(Ageratum conyzoides)的潜在分布, 发现MaxEnt模型较其他模型预测效果更好, 运行效率更高; 张海娟等[14]使用MaxEnt和GARP模型研究了薇甘菊(Mikania micrantha)在中国的适生区; 曹向锋等[13]利用MaxEnt等5种模型研究了黄顶菊(Flaveria bidentis)在中国的潜在适生区, 以上研究成果均证明MaxEnt模型对入侵物种的适生地预测模拟结果较好, 并且更为稳定。但总体而言, 目前利用生态位模型对动物类适生区[44]、病菌扩散趋势等[45, 46, 47, 48]研究较多, 而研究入侵植物适生区的相关研究较少。
本研究使用未来气候A1b情景数据, 通过ENFA筛选出影响紫茎泽兰分布的环境因子, 并对未来气候情景下紫茎泽兰在我国的入侵情况进行预测。然而, 该研究以当前紫茎泽兰的发生点为存在点, 当使用未来气候情境数据进行预测时, 这种假设可能会忽略未来紫茎泽兰实际存在点的影响, 假设是否充分可行还有待论证。此外, 影响紫茎泽兰分布的环境因素可能不仅仅是温度和降水, 土壤条件、人类活动、光照等都可能对紫茎泽兰的分布造成影响, 但是由于数据的限制, 本文仅分析了这两种气候变量对其适生区分布的影响。
本研究对紫茎泽兰的入侵概率进行等级划分(L1~L5), 认为L1、L2入侵等级为不入侵或入侵概率很低, 而L3~L5表示为发生入侵, 且入侵概率依次增加, L5表示为极易入侵等级。针对不同的入侵概率等级应采取不同的防治策略。对于低入侵等级L2, 可作为防御区, 对该区域进行定时观测, 一旦发现需及时根除; 而L3等级可作为扩散区, 采取人工拔除、化学防治相结合的手段, 避免对本地物种生长和物种多样性造成更深更恶劣的影响; 由于L4和L5等级紫茎泽兰的长势更甚, 难以根除, 所以需要采取人工拔除、化学防治、生物防治、生态调控、替代控制等综合防控技术, 并可进行加工利用, 将其作为饲料、肥料、卫生杀虫剂等。此外, 还需及时实施人工种植和自然恢复相结合的方法, 建立有益植被群落, 从而达到生态修复、长久控制害草的目标[2]。
在本研究中, 在计算不同入侵等级的中心点位置及中心点位移时采用Hart[34]和Yue等[35]的研究, 以各等级的面积作为权重进行计算。结果显示不同概率等级的中心点位移没有明显规律性, 其原因可能在于当前和未来气候数据的不连贯。当前气候数据起止时间为1961-1990年, 而未来气候数据起止时间为2010-2099年, 1990-2010年的数据缺失可能造成中心点移动方向的往复。其次, A1b数据的情景描述为全球经济快速增长, 人口在21世纪中期达到顶峰后下降, 科技迅速发展, 所有资源达到平衡[9, 49, 50]。而这种前提与现实情况显然还存在一些差距。在这种前提下, 由于人类在环境保护、资源平衡等方面的意识增强, 科技水平的迅速提高, 可能会对紫茎泽兰的入侵和扩散造成影响, 并缓解其继续扩散的趋势, 使中心点未出现非常明显的移动趋势, 且出现往复移动的情况。
王运生等[12, 51]通过使用6种生态位模型对胜红蓟的分布情况进行研究发现, 胜红蓟入侵我国后生态位已经发生改变, 并猜测可能是物种进化的原因使其适应新的生境, 但具体原因尚不确定。总的来说, 物种能够成功入侵主要原因在于在新的生存地缺乏竞争者和天敌, 存在生态位的空缺。入侵物种的原产地生态位和入侵地生态位可能会不同, 也就是可能存在生态位漂移的现象。根据Hutchinson[10]的生态位理论, 我们认为决定生态位的因素是多方面的, 不能仅根据气候和地形因素判断入侵物种的生态位是否发生漂移。判断生态位漂移的影响因子, 研究紫茎泽兰在我国的生态位是否发生偏移, 产生生态位漂移的原因及其造成的影响还有待在未来的研究中进一步的探讨。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|