




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
不同水分条件下冬小麦灌浆期茎叶可溶性碳水化合物积累转运与籽粒灌浆的关系
引用本文
马召朋, 栗孟飞, 杨德龙, 陈晓平, 陈菁菁, 刘媛, 李唯. 不同水分条件下冬小麦灌浆期茎叶可溶性碳水化合物积累转运与籽粒灌浆的关系. 23(4): 68-78
MA Zhao-peng, LI Meng-fei, YANG De-long, CHEN Xiao-ping, CHEN Jing-jing, LIU Yuan, LI Wei. Relationship between grain filling and accumulation and remobilization of water soluble carbohydrates in leaf and stem of winter wheat during the grain filling in different water conditions. Acta Prataculturae Sinica, 23(4): 68-78
Permissions
MA Zhao-peng, LI Meng-fei, YANG De-long, CHEN Xiao-ping, CHEN Jing-jing, LIU Yuan, LI Wei. Relationship between grain filling and accumulation and remobilization of water soluble carbohydrates in leaf and stem of winter wheat during the grain filling in different water conditions. Acta Prataculturae Sinica, 23(4): 68-78
Copyright©2014, 草业学报编辑部
本文属于开放获取期刊
不同水分条件下冬小麦灌浆期茎叶可溶性碳水化合物积累转运与籽粒灌浆的关系
作者简介:马召朋(1987-),男,河北石家庄人,在读硕士。E-mail:mazhaopeng303@126.com
摘要
以抗旱性有显著差异的两个冬小麦品种为材料,在干旱胁迫和正常灌溉条件下,研究了小麦灌浆期主茎旗叶和不同节位可溶性碳水化合物(WSC)积累转运与籽粒灌浆的关系。结果表明,小麦不同器官WSC含量,WSC转运率、WSC贡献率、主穗粒重和灌浆速率均受水分环境、发育阶段和/或器官更为显著的调控,但不同因子对各性状表型影响具有显著的特异性。小麦灌浆期不同器官WSC含量均呈现先升后降趋势。在干旱胁迫条件下,抗旱品种陇鉴19不同器官WSC含量峰值时间与正常灌溉相同,均在花后18 d,而水分敏感品种Q9086在花后12 d。干旱胁迫显著提高了不同器官WSC含量,促进了花前WSC转运,及其对籽粒粒重的贡献,其效应在陇鉴19主茎穗下节和倒二节更为显著,其WSC含量分别高达196.83和242.35 mg/g DW,WSC转运率高达66.15%和63.21%,WSC贡献率高达12.48%和8.67%。干旱胁迫缩短了籽粒灌浆进程,但陇鉴19较Q9086仍保持较高的灌浆速率和较大主穗粒重。小麦不同器官WSC含量与主穗灌浆速率以及WSC转运率与贡献率均呈正相关,且在干旱胁迫条件下,这种相关性更高。说明,干旱胁迫能促进小麦各器官WSC积累和转运,尤其是花前在穗下节和倒二节积累更多的WSC对补偿籽粒灌浆具有重要作用。
关键词:
小麦; 干旱胁迫; 可溶性碳水化合物; 积累转运; 籽粒灌浆
中图分类号:S512.1+1
文献标志码:A
文章编号:1004-5759(2014)04-0068-11
Relationship between grain filling and accumulation and remobilization of water soluble carbohydrates in leaf and stem of winter wheat during the grain filling in different water conditions
Abstract
Two winter wheat cultivars with different drought tolerance were performed to investigate the relationship between the grain filling and the accumulation and remobilization of water soluble carbohydrates (WSC) in flag leaves and different internodes of main stems during the grain filling in the drought stress (DS) and well watered (WW) conditions. Such tested traits as WSC contents, WSC remobilization rates and WSC contribution rates of different organs, grain weight and grain-filling rate were more significantly affected by the water condition, growth stage and/or organ, whereas different tested traits showed significant specificity in response to above-mentioned factors. The variation trend in WSC contents of different organs was of a rise first and then fall during the grain filling. The maximum WSC content of drought-tolerant cultivar Longjian 19 occurred on 18 DAA (days after anthesis) under both the DS and the WW, while did on 12 DAA in water-sensitive cultivar Q9086 in the DS. The DS might substantially increase WSC accumulation in different organs and improve WSC contribution rates to grain weight by remobilizing WSC built up in different organs during the pre-anthesis. The profit was more conducted in peduncle and penultimate internodes of Longjian 19. In this case, WSC contents, in turn, were up to 196.83 and 242.35 mg/g DW, WSC remobilization rates reached to 66.15% and 63.21%, and WSC contribution rates were up to a maximum of 12.48% and 8.67%, respectively. Although the DS shortened the course of grain filling, Longjian 19 still performed higher grain-filling rate and greater grain weight of main spikes. Positive correlations were found not only between WSC contents of different organs and grain-filling rates of main spikes, but also between WSC remobilization rates and its contribution rates. Furthermore, these correlations were significantly higher under the DS than the WW condition. It indicated that drought stress could enhance the accumulation and remobilization of WSC of different organs in wheat, and more buildup of WSC in peduncle and penultimate internodes might play an important role in compensating grain filling under the DS.
Keyword:
wheat; drought stress; water soluble carbohydrates; accumulation and remobilization; grain filling
引言
干旱是限制小麦(Triticum aestivum)产量的重要非生物胁迫因子, 每年因干旱造成小麦产量大量损失, 其危害相当于其他自然灾害之和[1]。灌浆期是小麦最终籽粒产量形成的关键时期, 同时也是对水分环境非常敏感的时期[2]。在我国北方雨养农业区, 小麦灌浆期的高频率季节性干旱往往导致小麦产量大幅度下降[3]。因此, 为了降低干旱对小麦生产的影响, 通过发掘小麦灌浆期抗旱的生物学机制, 改善小麦自身抗旱能力, 实现高产稳产, 是当今小麦抗旱研究的热点问题。
小麦营养器官中暂贮性可溶性碳水化合物(water soluble carbohydrates, WSC)是维系其生存和产量形成的重要代谢物质, 不仅通过渗透调节来缓冲逆境胁迫对小麦的伤害[4], 而且也是小麦籽粒灌浆所需的重要碳源[2]。尤其是在小麦灌浆期, 当干旱等逆境胁迫严重破坏小麦正常光合作用, 致使光合产物量无法满足冠层呼吸消耗和保持籽粒正常灌浆需要时, 小麦营养器官中暂贮性WSC对籽粒产量的形成尤为重要[2, 5, 6]。研究表明, 干旱胁迫能促进小麦花前积累在茎、叶、鞘和颖壳等营养器官中的WSC向籽粒转运, 对籽粒干物质积累的贡献率达10%~70%[5, 6, 7, 8, 9, 10], 从而改善籽粒灌浆和产量形成[5, 6, 8]。因此, 在干旱胁迫条件下, 促进小麦灌浆期各营养器官中WSC高效运转到籽粒中, 能不同程度地加速小麦灌浆而增加粒重, 不但可以补偿小麦籽粒产量损失[5, 6, 7, 8, 9, 10], 而且也是提高其水分利用效率的重要途径[11, 12, 13]。然而, 干旱调控小麦营养器官中暂贮性WSC高效运转到籽粒受到基因型、环境和基因型× 环境互作的显著影响[5, 6, 14]。进一步研究表明, 小麦不同器官和同一器官不同位置也对小麦WSC转运及其对粒重贡献具有显著影响[15, 16]。由此看出, 干旱调控小麦灌浆期营养器官WSC转运与产量形成的关系非常复杂。因此, 加速开展不同水分环境条件下不同基因型小麦灌浆期营养器官WSC转运与籽粒灌浆和产量形成关系研究, 有利于发掘小麦灌浆期更为丰富的抗旱生物学信息和解析籽粒灌浆的抗旱机制。为此, 本研究选用不同抗旱小麦品种陇鉴19(抗旱品种)和Q9086(高水肥品种)为供试材料, 设置不同水分处理, 研究不同水分条件下冬小麦灌浆期不同发育阶段主茎旗叶和不同节位WSC动态积累和转运与籽粒灌浆的关系, 旨在揭示不同水分环境下小麦主要营养器官WSC 积累和转运在籽粒灌浆中的作用及其相互关系, 为干旱区小麦品种改良及高效栽培提供理论依据。
1 材料与方法
1.1 供试材料
利用抗旱性强的冬小麦品种陇鉴19和高水肥品种Q9086为供试材料。陇鉴19 是甘肃省农业科学院旱地农业研究所选育的新品种, 为甘肃省第一个大面积应用的抗旱北移冬小麦新品种, 抗旱性强, 耐瘠薄, 对水肥反应不敏感。Q9086 是西北农林科技大学农学院小麦研究所提供的品系, 对水肥反应敏感, 尤其灌浆期对水分反应敏感。
1.2 试验设计
试验于2011年10月-2012年6月在甘肃省兰州市安宁区小麦试验点(36° 04' N, 103° 51' E)进行。小麦播前基肥施用量为N 180 kg/hm2, P2O5 120 kg/hm2, K2O 75 kg/hm2, 在整个生育期内均不再施肥。试验为随机区组设计, 每处理3次重复, 均为稀条播, 行长2 m, 行距0.2 m, 每行点播120粒, 12行区。田间试验水分管理分为雨养, 即干旱胁迫(DS)和灌溉(WW)2个处理, 2011年播前均统一灌底墒水(900 m3/hm2)。灌溉处理在2012年拔节期, 抽穗期和开花期补充灌水, 每次灌水量为750 m3/hm2。干旱胁迫处理仅在拔节期灌水750 m3/hm2, 其后完全依靠自然降水(2011年10月-2012年6月小麦全生育期降水量为116 mm)。
1.3 目标性状测定
随机选择供试小麦品种开花期和长势一致的主茎挂牌标记, 从花后当天开始, 每6 d取一次样直到成熟期。每个处理每次取15个主茎, 取旗叶(包括叶鞘, FL), 穗下节(PedI)、倒二节(PenI)和倒三节(ThiI), 105℃杀青30 min, 80℃烘干至恒重, 分别测定旗叶和主茎不同节位的干重, 求其平均值; 然后将样品剪成1~2 mm长的碎片, 贮存在干燥器中待测。穗部风干后, 脱粒测定主穗粒重(GWMS), 计算灌浆速率(GFR)[17]。不同处理各营养器官WSC总含量测定采用蒽酮比色法[18], 所有测定均3次重复。小麦旗叶和主茎不同节位WSC花前转运率=(花后当天WSC绝对含量-成熟期WSC绝对含量)/花后当天WSC绝对含量× 100%; WSC花后转运率=(最大WSC绝对含量-花后当天WSC绝对含量)/最大WSC绝对含量× 100%; WSC花前贡献率=(花后当天WSC绝对含量-成熟期WSC绝对含量)/(1000× GWMS)× 100%; WSC花后贡献率=(最大WSC绝对含量-花后当天WSC绝对含量)/(1000× GWMS)× 100%。其中, WSC绝对含量=WSC浓度× 干品质。
1.4 数据处理
采用DPS v7.05统计软件进行不同处理间各目标性状方差(ANVOA)分析、多重比较(Duncan’ s法)、相关分析和通径分析。
2 结果与分析
2.1 小麦各性状表型方差分析
小麦灌浆期主茎不同器官WSC含量, WSC转运率, WSC贡献率, 主穗粒重和灌浆速率受基因型, 水分环境, 发育阶段和/或器官的显著(P≤ 0.05)或极显著(P≤ 0.01)影响; 并且, 各因子间还普遍存在显著或极显著的互作效应(表1)。根据各变异来源占总处理变异(平方和)的百分比(SSa/SST), 可比较各因子对目标性状变异的相对贡献率[19]。从表1看出, 水分和发育阶段是影响WSC含量变异的主要因素, 二者对WSC含量表型变异的相对贡献率分别达32.35%和25.96%; 器官、发育阶段及发育阶段与水分互作对WSC转运率变异具有重要作用, 其对表型变异的相对贡献率分别达30.19%, 20.20%和10.91%; WSC贡献率受器官和水分的影响较大, 其对表型变异的相对贡献率分别为54.16%和12.61%; 发育阶段是影响主穗粒重和灌浆速率变异的主要因子, 其对两性状表型变异的相对贡献率分别高达85.88%和52.90%, 并且发育阶段与水分互作对灌浆速率变异也具有重要影响, 对表型变异的相对贡献率为36.63%。其他因子及其互作均对各目标性状影响相对较小。
2.2 不同水分条件下小麦灌浆期不同器官WSC含量变化
小麦主茎旗叶和不同节位WSC含量随着小麦花后生育期进程, 均呈现不同程度先升后降的趋势(图1), 但WSC含量峰值出现的时间受水分环境、基因型和器官的影响而不同。在正常灌溉条件下, 所有处理WSC含量峰值均出现在花后18 d; 不同基因型和器官间WSC含量差异较小。在干旱胁迫条件下, 陇鉴19所有器官WSC含量峰值均出现在花后18 d, 但Q9086的主茎旗叶、穗下节和倒二节WSC含量峰值较陇鉴19提前6 d, 倒三节与陇鉴19相同。在WSC含量达到峰值前, 干旱胁迫条件下的陇鉴19各个器官WSC含量显著高于Q9086, 尤其是主茎旗叶, 穗下节和倒二节这种变化更为显著; 但随后两品种随着WSC含量下降其差异减小, 甚至在花后24 d后, 陇鉴19主茎旗叶, 穗下节和倒二节的WSC含量显著低于Q9086。在不同器官间, 两品种主茎旗叶、穗下节和倒二节WSC含量显著高于倒三节。
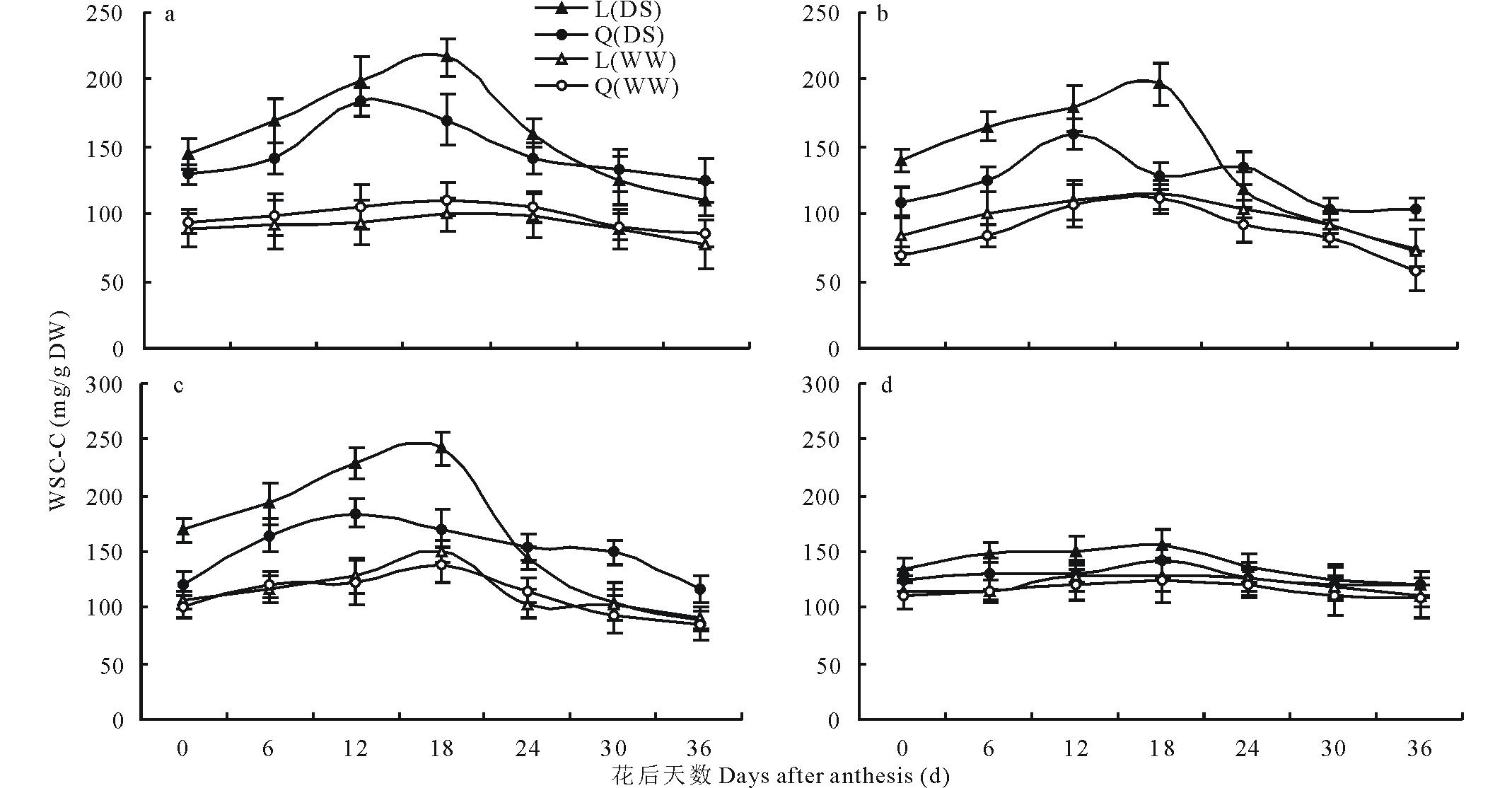 | 图1 不同水分条件下小麦灌浆期不同器官WSC含量变化Fig.1 Changes in WSC contents of different organs of wheat during grain filling in different water conditions ▲和●分别表示干旱胁迫(DS)条件下陇鉴19(L)和Q9086(Q)的WSC含量(WSC-C); △ 和○分别表示灌溉(WW)条件下陇鉴19(L)和Q9086(Q)的WSC含量。图1a~图1d分别表示旗叶、穗下节、倒二节和倒三节的WSC含量。The symbols, ▲ and ●, represent WSC contents (WSC-C) of Longjian 19 (L) and Q9086 (Q) in the drought stress (DS), respectively; △ and ○ represent WSC contents of Longjian 19 (L) and Q9086 (Q) in the well-watered (WW), respectively. Fig.1a-Fig.1d show WSC contents of such organs as the flag leaf, peduncle internode, penultimate internode, and the third internode from top. |
2.3 不同水分条件下小麦主穗粒重和灌浆速率变化
小麦主穗粒重随着发育进程而逐渐增加, 但不同水分环境和基因型具有显著差异(图2)。在干旱胁迫条件下, 陇鉴19和Q9086主穗粒重在花后18 d前快速增加, 随后缓慢增加, 至成熟期陇鉴19主穗粒重显著高于Q9086达19.79%。正常灌溉条件下, 小麦主穗粒重增加趋势与干旱胁迫的相似, 但其主穗粒重快速增长期延续到花后24 d, Q9086成熟期主穗粒重显著高于陇鉴19达32.80%。在干旱胁迫条件下, 陇鉴19成熟期主穗粒重下降仅为正常灌溉的11.61%, 而Q9086高达44.43%。在两种水分条件下, 陇鉴19和Q9086的灌浆速率均呈现单峰变化趋势(图2)。其中, 在干旱胁迫条件下, 两品种在花后6~18 d时灌浆速率急剧升高, 随后显著降低, 二者灌浆速率在花后18 d时差异达到最大, 陇鉴19高出Q9086达44.56%; 而在正常灌溉条件下, 两品种在花后12~24 d时灌浆速率急剧升高, 随后显著降低, 二者灌浆速率最大差值出现在花后24 d, Q9086高出陇鉴19达64.96%。
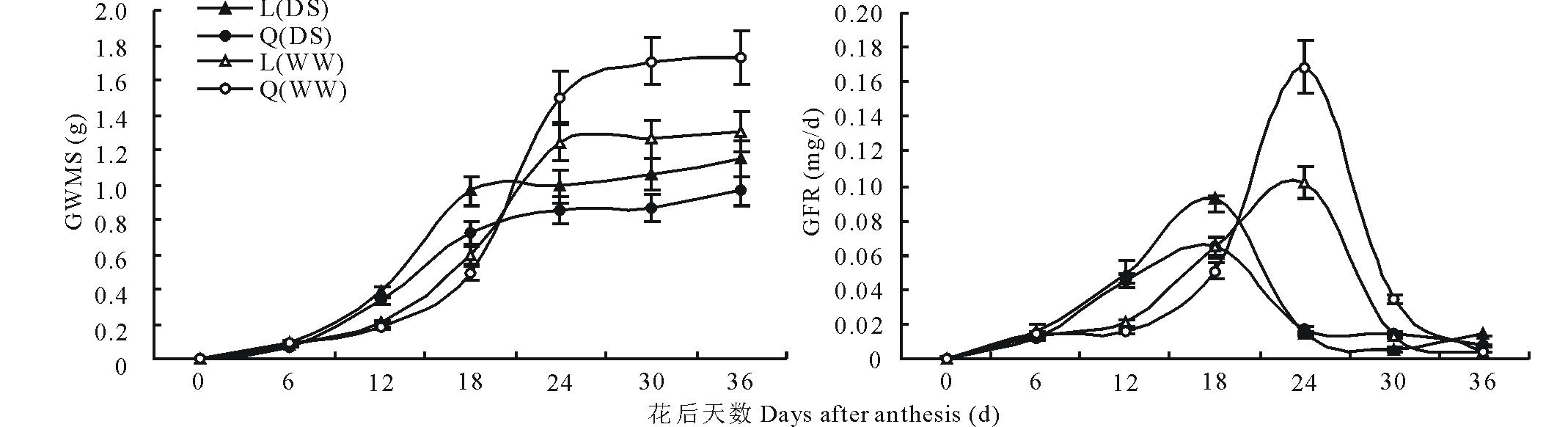 | 图2 不同水分条件下小麦主穗粒重(GWMS)和灌浆速率(GFR)变化Fig.2 Changes in grain weight per main spike (GWMS) and its grain-filling rate (GFR) of wheat in different water conditions ▲和●分别表示干旱胁迫(DS)条件下陇鉴19(L)和Q9086(Q)的表型性状(GWMS和GFR); △ 和○分别表示灌溉(WW)条件下陇鉴19(L)和Q9086(Q)的表型性状(GWMS和GFR)。The symbols, ▲ and ●, represent phenotypes (GWMS and GFR) of Longjian 19 (L) and Q9086 (Q) in the drought stress (DS), respectively; △ and ○ represent phenotypes (GWMS and GFR) of Longjian 19 (L) and Q9086 (Q) in the well-watered (WW), respectively. |
2.4 不同水分条件下小麦不同器官WSC含量和主穗灌浆速率的相关分析和通径分析
在不同水分条件下, 小麦不同器官WSC含量间, 及其与主穗灌浆速率间均呈不同程度的正相关, 相关系数(r)为0.27~0.99* * (表2)。其中, 干旱胁迫条件下陇鉴19不同器官间WSC含量的相关系数(rDS=0.95* * ~0.99* * )显著高于正常灌溉的(rWW=0.70~0.92* * ), 但Q9086相反(rDS=0.56~0.87* , rWW=0.90* * ~0.98* * )。两小麦品种不同器官WSC含量与主穗灌浆速率间的相关系数均表现为干旱胁迫(rDS=0.60~0.89* * )显著高于正常灌溉(rWW=0.27~0.73* ), 且正常灌溉的普遍相关不显著。在干旱胁迫条件下, 品种间和器官间WSC含量与主穗灌浆速率的相关性差异较小, 而正常灌溉条件的差异较大, 其中, Q9086和倒二节的相关系数显著降低。
| 表2 不同水分条件下小麦不同器官WSC含量和主穗灌浆速率间的相关系数 Table 2 Correlation coefficients among WSC contents of different organs and grain-filling rates of main spike of wheat in different water conditions |
通径分析表明(表3), 在不同水分条件下, 小麦两品种旗叶WSC含量对主穗灌浆速率均具有正向直接作用(P0i=0.84~2.12), 其效应值在干旱胁迫处理和陇鉴19中较高, 而该性状与其他器官WSC含量的间接总效应为负向作用(P0i=-0.23~-1.29)。在干旱胁迫条件下, 陇鉴19穗下节、倒二节和倒三节WSC含量分别对主穗灌浆速率较Q9086具有较高的直接作用效应, 但品种间作用方向相反。其中, 陇鉴19穗下节表现出正向直接作用(P0i=0.83), 倒二节(P0i=-1.14)和倒三节(P0i=-1.01)表现出负向直接作用。主茎各节位对应的间接总效应, 陇鉴19穗下节表现出较低的负向作用, 倒二节和倒三节表现出较高正向作用, 但Q9086与之相反。在正常灌溉条件下, 两小麦品种各节位WSC含量对主穗灌浆速率直接和间接作用方向相同, Q9086的效应值普遍高于陇鉴19。
2.5 不同水分条件下小麦不同器官WSC转运率
不同水分条件下, 陇鉴19和Q9086主茎不同器官花前和花后WSC转运率有显著差异, 不同处理间WSC转运率在3.27%~66.19%之间变化(图3)。两品种主茎不同器官花前WSC转运率普遍高于花后的, 二者依器官不同其差值为0.72%~54.29%, 干旱胁迫条件下这种差异较正常灌溉更为显著。在干旱胁迫条件下, 陇鉴19主茎不同器官花前WSC转运率显著高于Q9086, 二者差异为8.05%~36.35%。但两品种间不同器官花后WSC转运率普遍差异不显著。在正常灌溉条件下, 小麦两品种间和花前花后间WSC转运率差异较小, 甚至不显著, 但更多表现出器官间的差异, 即主茎穗下节和倒二节WSC转运率较高(21.92%~37.65%), 旗叶次之(10.44%~21.92%), 倒三节较小(5.78%~10.06%)。相似地, 在干旱胁迫下, 陇鉴19和Q9086 WSC转运率也表现出更为显著的器官间差异, 主茎穗下节和倒二节WSC转运率达3.27%~66.19%, 旗叶为13.65%~41.87%, 倒三节为6.83%~21.52%。
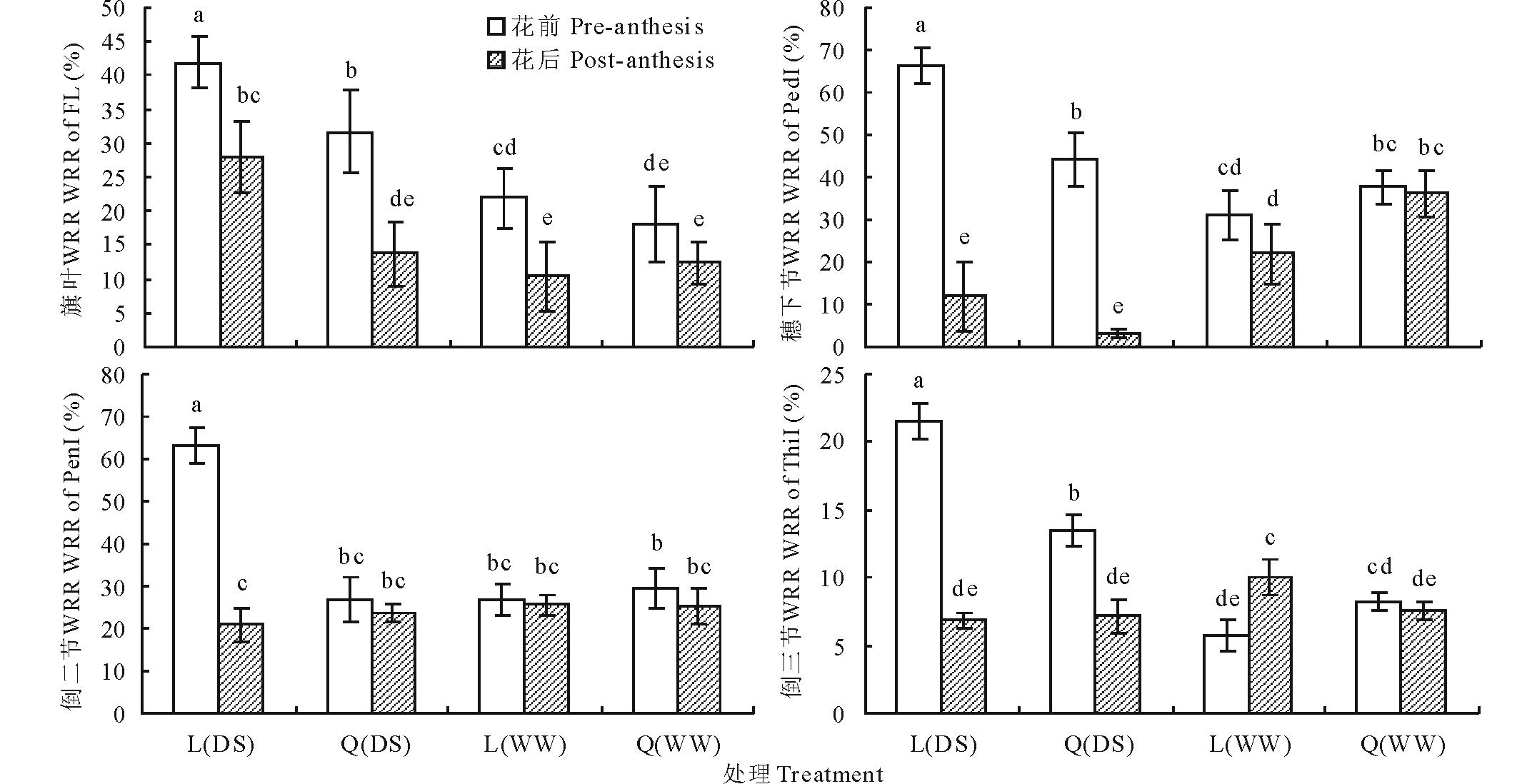 | 图3 不同水分条件下小麦不同器官花前和花后WSC转运率(WRR)Fig.3 WSC remobilization rate (WRR) of different organs of wheat during the pre- and post-anthesis in different water conditions |
FL, PedI, PenI和ThiI分别表示旗叶、穗下节, 倒二节和倒三节。L和Q分别表示陇鉴19和Q9086; DS和WW分别表示干旱胁迫和灌溉条件。图中不同小写字母表示同一因素处理间差异显著(P< 0.05)。下同。FL, PedI, PenI and ThiI show the flag leaf, peduncle internode, penultimate internode, and the third internode from top, respectively. L and Q mean Longjian 19 and Q9086, respectively; DS and WW mean the drought stress and well-watered conditions, respectively. Different small letters mean significant difference (P< 0.05) between treatments under the same factor.The same below.
2.6 不同水分条件下小麦不同器官WSC转运对籽粒粒重的贡献率
不同水分条件下, 陇鉴19和Q9086主茎不同器官花前和花后WSC转运对籽粒粒重的贡献率在0.33%~12.48%之间变化, 受基因型、水分环境、器官和发育阶段的显著影响(图4)。两品种主茎不同器官花前WSC贡献率普遍高于花后的, 二者依器官不同其差值为0.04%~5.76%; 干旱胁迫条件下两品种主茎不同器官WSC贡献率普遍显著高于正常灌溉。在干旱胁迫条件下, 陇鉴19主茎不同器官花前和花后WSC贡献率显著高于Q9086; 而在正常灌溉条件下, 品种间差异普遍较小, 大部分差异不显著。并且, 在不同水分条件下, 两品种主茎穗下节和倒二节均表现出较高的WSC贡献率(1.80%~12.48%), 而旗叶和倒三节的较低, 仅为0.33%~1.58%。从两小麦品种主茎不同器官花前和花后WSC转运率及其对籽粒粒重的贡献率之间的线性相关分析(图5)看出, WSC转运率及贡献率之间普遍呈极显著正相关。在干旱胁迫条件下, 花前WSC转运率与贡献率之间的相关性较高(r=0.87* * ), 而花后的几乎不相关(r=0.01); 在正常灌溉条件下, 二者相关性均较高, 且花后(r=0.92* * )的高于花前的(r=0.81* * )。
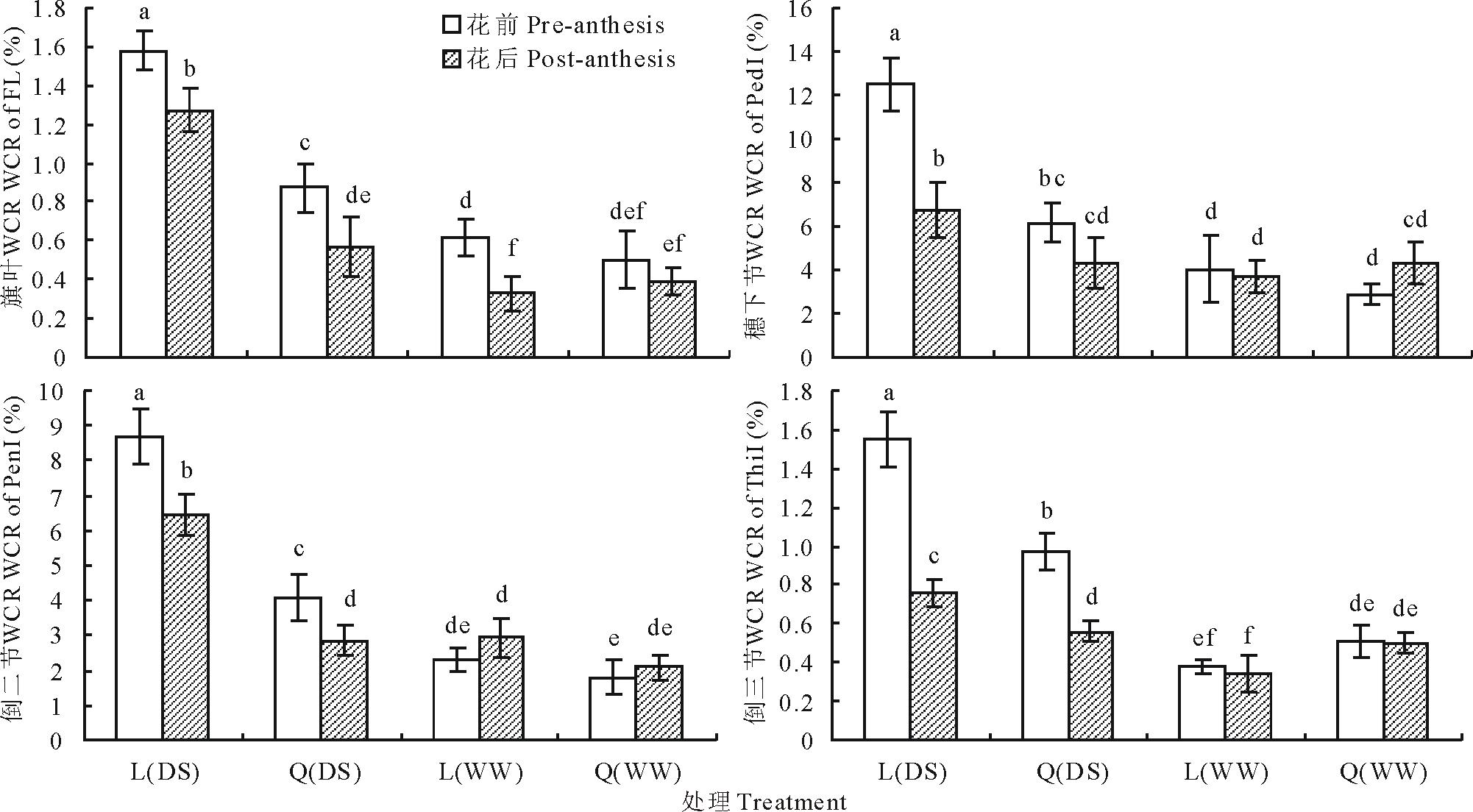 | 图4 不同水分条件下小麦不同器官花前和花后WSC贡献率(WCR)Fig.4 WSC contribution rate (WCR) of different organs of wheat during the pre- and post-anthesis in different water conditions |
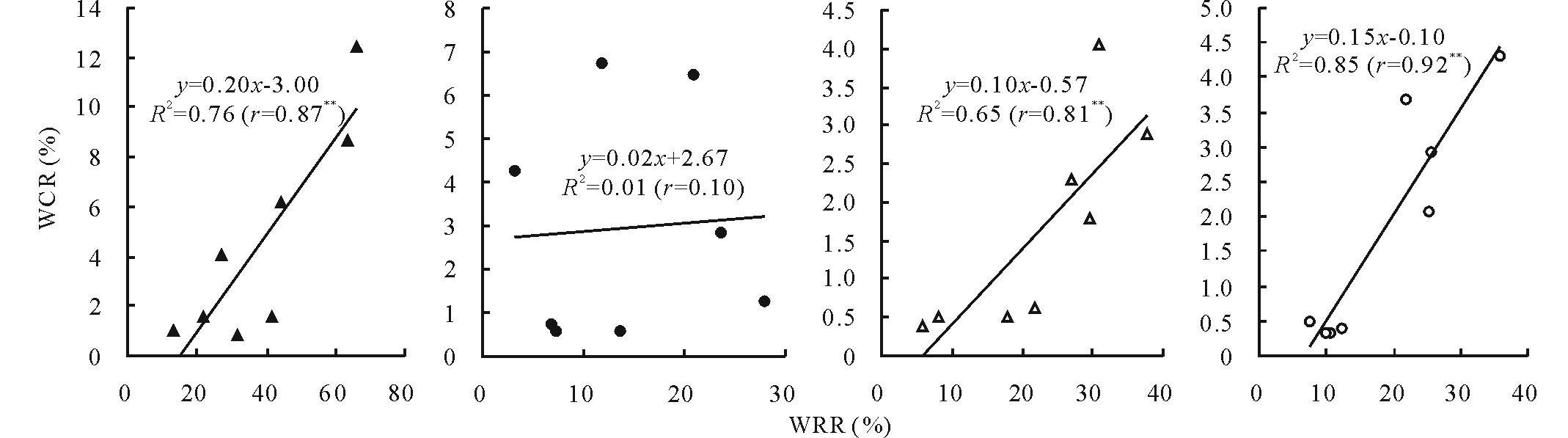 | 图5 不同水分条件下小麦不同器官WSC转运率(WRR)与贡献率(WCR)的相关性Fig.5 Correlations between WSC remobilization rate (WRR) and contribution rate (WCR) of different organs of wheat under different water conditions ▲和●分别表示干旱胁迫条件下花前和花后WRR和WCR之间的相关性; △ 和○分别表示正常灌溉条件下花前和花后WRR和WCR之间的相关性。▲ and ● mean correlations between WRR and WCR during the pre- and post-anthesis under the drought stress, respectively; △ and ○ mean correlations between WRR and WCR during the pre- and post-anthesis under the well-watered, respectively. |
3 讨论
3.1 基因型和非基因型因子对小麦WSC积累转运和籽粒灌浆的影响
研究表明, 可溶性碳水化合物(WSC)是小麦各营养器官主要暂贮性的生理代谢物质, 对调节小麦生长发育中的新陈代谢[20], 维系生存和产量形成具有重要作用[2, 4, 5, 6]。但小麦WSC积累转运和籽粒灌浆的生理过程易受基因型、环境、器官及其互作效应等因子的显著影响[5, 6, 9, 14, 15, 16]。在本研究中发现, 小麦灌浆期主茎不同器官WSC含量、WSC转运率和贡献率, 以及主穗粒重和灌浆速率受基因型, 水分环境, 发育阶段和/或器官, 及其各因子互作效应的显著或极显著影响, 但不同因子对各性状表型变异的主导作用具有显著的特异性。其中, 水分和发育阶段对WSC含量, 水分、器官、发育阶段及发育阶段与水分互作对WSC向籽粒转运, 以及发育阶段和发育阶段与水分互作对主穗粒重和灌浆速率表型变异的影响分别表现出显著的主导作用。非基因型因子对小麦WSC积累转运和籽粒灌浆的影响显著高于基因型。Ehdaie等[5]也发现小麦灌浆期茎秆不同节位干物质积累转运显著受到年份、水分、基因型, 发育阶段, 及基因型× 发育阶段的显著影响; 进一步分析花后不同节位WSC含量变化发现, 发育阶段是影响小麦茎秆WSC积累转运相关性状最为主要的因子, 其次是基因型, 水分和年份, 并且基因型与发育阶段的互作对影响WSC积累转运具有重要作用[6]。Yang等[14]和孟维伟等[15]认为发育时期和水分对小麦茎秆WSC含量, WSC转运和灌浆速率的影响显著大于基因型; 并且, 小麦不同器官和同一器官不同位置也对小麦WSC积累转运具有显著影响[15, 16]。由此说明, 水分、发育阶段和器官是影响小麦WSC积累转运和籽粒灌浆最为主要的调控因子, 在基因型基础上, 小麦WSC积累转运可能通过一定的时空表达, 协同优化各调控因子, 来最大限度适应环境, 维系其生存和产量形成。
3.2 干旱胁迫条件下小麦灌浆期各器官WSC含量动态变化与籽粒灌浆的关系
在干旱胁迫条件下, 小麦不同营养器官WSC作为重要的渗透调节物质通过提高其含量来调控植株生理代谢, 以减小干旱胁迫对其正常生长发育的伤害[4]; 在农艺性状上, 通过降低地上部分生长量, 减小粒重和产量等来适应干旱胁迫[3, 10, 21]。Rebetzke等[22]发现小麦茎秆WSC含量分别与产量和穗粒重呈显著或极显著的正相关。Yang等[14]发现小麦灌浆中期整个主茎WSC含量与籽粒千粒重和灌浆效率均表现出显著或极显著的正相关性, 且干旱胁迫条件下的相关系数显著高于灌溉条件。在本研究中, 干旱胁迫显著提高了小麦灌浆期旗叶、穗下节和倒二节WSC含量, 促进了籽粒提前灌浆, 缩短了灌浆进程, 降低了主穗粒重, 但抗旱品种陇鉴19仍保持较高WSC含量、灌浆速率和主穗粒重。相关分析表明, 在干旱胁迫条件下, 小麦不同性状间普遍呈显著或极显著的正相关, 且普遍具有较高相关系数(r=0.56~0.99* * )。综上说明, 在干旱胁迫条件下, 小麦不同器官WSC含量与灌浆速率和产量相关因子密切相关, 即高的WSC含量可能诱导较高灌浆速率、粒重和最终产量, 而这种关系在抗旱品种更为显著。通径分析表明, 旗叶WSC含量对主穗灌浆速率均具有正向直接作用和负向间接作用, 但效应值在干旱胁迫处理和陇鉴19中较高; 但主茎不同节位WSC含量对主穗灌浆速率作用因基因型和水分有显著差异, 甚至作用方向和效应值大小完全相反(表3)。说明小麦旗叶作为重要的WSC的“ 源” 对籽粒灌浆的作用相对稳定, 而茎秆不同节位的WSC易受水分和基因型调控而动态作用于籽粒灌浆, 尤其在干旱胁迫条件下这种作用可能更有利于籽粒灌浆, 即表现出更为显著的正相关性。
| 表3 不同水分条件下小麦不同器官WSC含量对主穗灌浆速率的通径分析 Table 3 Path analysis of WSC contents of different organs to grain-filling rates of main spike of wheat in different water conditions |
3.3 干旱胁迫条件下小麦灌浆期各器官WSC转运与籽粒灌浆的关系
在干旱胁迫条件下, 由于小麦光合器官的加速衰老, 光合能力下降, 当光合产物量无法满足冠层呼吸消耗和保持籽粒正常灌浆需要时, 小麦籽粒灌浆和最终产量形成更加依赖于花前积累在各营养器官WSC转运[2, 5, 6], 这种贡献率可达10%~70%[5, 6, 7, 8, 9, 10]。并且, 适度干旱有利于小麦茎秆WSC积累转运[5, 6, 15], 但不同基因型所达到WSC高效积累转运的需水模式不同[15]。本研究发现, 在不同水分条件下, 两品种主茎不同器官花前WSC转运率和贡献率显著高于花后, 干旱胁迫条件下的普遍高于正常灌溉, 抗旱品种陇鉴19显著高于水分敏感品种Q9086。尤其是在干旱胁迫条件下, 抗旱品种主茎穗下节和倒二节均表现出较高的花前WSC转运率和贡献率; 且花前WSC转运率与贡献率具有较高相关性, 而花后的几乎不相关。Ma等[23]利用不同抗旱性的春小麦品种研究干旱调控茎秆干物质转运对籽粒产量补偿效应中发现, 在干旱胁迫条件下, 上部茎秆(穗下节和倒二节)干物质转运率和对籽粒的贡献率显著高于下部茎节(倒三节以下), 抗旱品种显著高于水分敏感品种; 干旱胁迫能显著促进花前积累在茎秆中的干物质向籽粒高效转运, 来补偿产量损失。由此说明, 在干旱胁迫条件下, 小麦穗下节和倒二节茎中暂贮性WSC积极转运对促进小麦籽粒灌浆和补偿产量具有重要作用。因此, 在小麦抗旱育种中, 小麦灌浆期上部茎秆高的含糖量可以作为一个有效的指标进行选择育种。4 结论水分、发育阶段和器官是影响小麦灌浆期主茎旗叶和不同节位WSC积累转运和籽粒灌浆最为主要的影响因子。
干旱胁迫缩短了小麦灌浆进程, 显著提高了小麦主茎不同器官WSC含量, 促进了花前WSC转运及其对籽粒粒重的贡献, 其效应在抗旱品种陇鉴19主茎穗下节和倒二节更为显著, 其WSC含量分别高达196.83和242.35 mg/g DW, WSC转运率高达66.15%和63.21%, WSC对籽粒灌浆的贡献率高达12.48%和8.67%。
在干旱胁迫条件下, 小麦主茎各器官WSC积累转运与籽粒灌浆间普遍呈显著或极显著的正相关, 抗旱品种陇鉴19相关性更高。因此, 在小麦抗旱育种中, 小麦灌浆期上部茎秆高的含糖量可以作为一个有效的指标进行选择育种。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|