{kind=link}
北方白菜型冬油菜F2主要生理生化特性的变异与抗寒性相关分析
引用本文
孔德晶, 王月, 孙万仓, 曾秀存, 方彦, 鲁美宏, 杨宁宁. 北方白菜型冬油菜F2主要生理生化特性的变异与抗寒性相关分析 . 23(4): 79-86
KONG De-jing, WANG Yue, SUN Wan-cang, ZENG Xiu-cun, FANG Yan, LU Mei-hong, YANG Ning-ning. Analysis of variation in physio-biochemical characteristics and cold resistance in winter rapeseed F2 populations . Acta Prataculturae Sinica, 23(4): 79-86
Permissions
KONG De-jing, WANG Yue, SUN Wan-cang, ZENG Xiu-cun, FANG Yan, LU Mei-hong, YANG Ning-ning. Analysis of variation in physio-biochemical characteristics and cold resistance in winter rapeseed F2 populations . Acta Prataculturae Sinica, 23(4): 79-86
Copyright©2014, 草业学报编辑部
本文属于开放获取期刊。
北方白菜型冬油菜F2主要生理生化特性的变异与抗寒性相关分析
作者简介:孔德晶(1988-),女,山东滕州人,在读硕士。E-mail:kongying19881004@163.com
摘要
对白菜型冬油菜F2群体主要生理生化特性的变化进行分析,利用陇油7号和陇油9号杂交得到103个F2单株,对越冬率、幼苗习性、超氧化物歧化酶(SOD)等8个指标进行调查、测定。对所得数据进行方差分析、聚类分析、相关分析、通径分析。结果表明,除SOD活性外,其余各指标在F2群体中呈连续变异,分布频率大致接近正态分布,同时存在双向超亲分离现象。在聚类距离为3.58处,F2群体分为 8个类群,群体间具有较大的遗传差异。相关分析表明所有指标与越冬率均有显著或极显著相关。各生理生化指标对越冬率的通径系数与相关系数方向均一致,但通径系数更有利于解释其本质,通过通径分析得到各指标对白菜型冬油菜越冬率相对重要性从大到小的顺序是:幼苗习性,SOD活性,MDA含量,游离脯氨酸含量,POD活性,CAT活性,可溶性蛋白含量。幼苗习性对越冬率主要是通过直接影响起作用,影响率达79.8%,CAT活性对越冬率的影响主要是通过间接影响起作用的,间接影响率达68.7%,而其余各指标对越冬率的影响均是通过直接和间接影响共同起作用的。这些结果可为白菜型冬油菜数量性状抗寒基因定位分析提供有用的信息。
关键词:
白菜型冬油菜; F2群体; 分离; 聚类分析; 相关分析; 通径分析
中图分类号:S634.01
文献标志码:A
文章编号:1004-5759(2014)04-0079-08
Analysis of variation in physio-biochemical characteristics and cold resistance in winter rapeseed F2 populations
Abstract
Overwintering rates, seedling habits, SOD activity, and five other physio-biochemical indicators in 103 F2 populations of a winter rapeseed ( Brassica rapa) cross between Longyou 7 and Longyou 9 were studied using variance analysis, correlation analysis, cluster analysis and path analysis. The frequency of all indicators (except SOD) activity in the F2 populations was approximately normally distributed but with a slight skew. A tremendous transgressive segregation for all indicators was observed in the populations. At a cluster distance of 3.58, F2 populations were divided into eight groups, with great genetic differences. Correlation analysis showed that all indicators had significant differences in overwintering rates. The path coefficient and correlation coefficient between all indicators and overwintering rate were consistent, but the explanation of their relationships were better explained by the path coefficient. Through path analysis, the relative importance in descending order of B. rapa winter survival rate was: Seedling habits, SOD activity, MDA content, free proline content, POD activity, CAT activity, and soluble protein content. Seedling habits directly impacted winter survival rates, and the influence reached 79.8%, but CAT activity was mainly through indirect effects and the indirect affect rate was 68.7%, with the rest of the indexes co-acting through both direct and indirect effects. These results provide useful information for quantitative trait loci (QTL) mapping.
Keyword:
winter rapeseed (Brassica rapa ); F2 populations; segregation; cluster analysis; correlation analysis; path analysis
引言
白菜型油菜(Brassica rapa)是芸薹属6个栽培物种之一, 也是世界上广泛栽培的3大类型油菜之一, 其栽培历史悠久, 不仅是重要的油料作物, 而且是极其重要的蔬菜作物[1, 2, 3]。目前, 我国油菜的栽培面积约700万hm2, 其中, 白菜型油菜面积15%左右[4]。中国是世界公认的白菜型油菜起源地之一[2], 具有包括农家品种和推广栽培品种在内的丰富资源, 它们在我国各地具有悠久的栽培历史。其中, 白菜型冬油菜具有耐迟播、生育期短, 耐贫瘠、抗寒性强的突出优点。在长期的自然和栽培条件的选择下, 白菜型冬油菜表现出较好的抗寒性和适应性, 具有甘蓝型油菜和芥菜型油菜所不可替代的作用。近年来, 由于较好地解决了品种的抗寒问题[5, 6], 白菜型冬油菜已经成功引入甘肃河西走廊、新疆阿勒泰、塔城、乌鲁木齐、青海、宁夏、北京等地, 扩大了冬油菜在我国的种植范围, 使种植区域向北推进纬度约13° , 海拔提高到2300 m左右, 取得了显著经济效益、生态效益与社会效益[7, 8, 9, 10, 11]。由于白菜型冬油菜可以有效的减少我国北方春季沙尘源, 保护环境, 同时对改革北方传统的一年一熟种植制度为两年三熟制或一年两熟制, 提高复种指数, 增加单位土地面积经济效益具有重要意义。北方冬油菜研究愈来愈引起人们的广泛关注[10, 12]。
目前, 对白菜型冬油菜抗寒性的研究主要集中在冬油菜品种的适应性[8]、生长发育特性[13]、产量及经济特性方面[14, 15]、相关抗寒基因克隆及表达分析[16, 17], 如孙万仓等[5]对不同冬油菜类型在西北地区的越冬性等进行了分析比较, 认为不同白菜型冬油菜品种(系)抗寒性存在巨大差异; 魏文慧等[14]对冬油菜的越冬率、产量及经济性状的影响进行了分析研究; 张腾国等[17]从油菜中克隆了抗寒基因 BnICE1, 并分析了其在油菜适应低温胁迫中发挥的重要作用。此外也有对构建白菜型油菜连锁图谱[18, 19]和白菜型油菜幼苗绒霉病的基因定位研究, 如Su和Graham[18]利用分子标记技术对白菜型油菜连锁图谱的构建, 为白菜型油菜基因序列和遗传信息的研究提供依据; Yu和Zhang[20]利用分子标记技术对白菜型油菜幼苗绒霉病进行QTL定位的研究。而目前对白菜型冬油菜F2群体生理生化的变异与抗寒性相关分析及抗寒性遗传的研究鲜见报道。研究表明, 植物的抗寒性状常是由微效多基因或由具有加性效应的主基因与微效基因所控制[21, 22]的, 表现为数量性状, 盖钧镒等[23]提出了针对植物数量性状分析的一整套方法, 被广泛应用于植物各类数量性状, 包括产量[24]、品质[25]、抗病虫性[26, 27]和耐逆性[28, 29, 30]等的遗传分析, 确定控制目标性状主基因的数目及它们的基因效应, 为植物目标性状的遗传改良提供了指导。本研究对103株F2代白菜型冬油菜抗寒性相关的8个生理生化指标的变异进行了统计与测定, 分析各指标在群体中的遗传变异及分布情况, 以期为白菜型冬油菜的抗寒性遗传研究提供理论依据。
1 材料与方法
1.1 试验材料及试验区自然概况
供试品种为白菜型冬油菜, 利用亲本陇油7号、陇油9号杂交配制F1代, 以及F1代自交得到的103株F2代个体为材料。母本陇油7号抗寒性超强, 为国内外最优抗寒品种, 父本陇油9号抗寒性较弱。 试验设于甘肃农业大学试验基地甘肃省兰州沙井驿试验田, 2012年8月, 将亲本、F1代和F2代分别种植于4个小区, 小区长为2.5 m, 宽1.4 m, 小区面积3.5 m2, 每小区8行, 小区间距为40 cm。试验地前茬为小麦(Triticum aestivum), 施农家肥60000 kg/hm2、磷酸二铵300 kg/hm2。田间管理按大田常规生产进行。
1.2 生理生化指标的测定
在2012年11月, 在亲本和F1代种植小区内, 各处理随机取样10株, 对F2代单株定株、取样, 各取样植株进行生理生化指标测定, 设3次重复。超氧化物歧化酶(SOD)活性的测定采用氮蓝四唑(NBT)光还原法[31] , 过氧化物酶(POD)活性测定采用愈创木酚比色法[32], 过氧化氢酶(CAT)活性测定采用紫外吸收法[31], 丙二醛含量测定采用硫代巴比妥酸(TBA)显色法[32], 游离脯氨酸含量测定采用甲苯萃取法[31], 可溶性蛋白含量测定采用考马斯亮蓝G-250 法[31]。
1.3 幼苗习性调查与越冬率的统计
对幼苗生长习性进行目测评定, 1级(匍匐型):叶片全部匍匐在地面; 2级(半匍匐型):叶片的叶尖着地面, 而叶身还是离开地面; 3级(半直立型):叶片全部离开地面, 但没完全直立; 4级(直立型):叶片全部直立。调查各处理小区越冬前苗数, 返青期苗数, 计算各处理越冬率(返青期苗数与越冬前苗数的百分比)。另外, 对103株F2群体进行单株定株, 存活植株记为1, 死亡植株记为0。
1.4 数据处理及分析
利用Microsoft Office Excel 2010对数据进行处理并作图; 利用SPSS软件[33] 对数据进行方差分析, 全子集回归分析, 相关分析, 通径分析; 利用DPS 3.01对数据进行聚类分析。
2 结果与分析
2.1 F2群体主要生理生化特性的分离分析
由亲本(P1、P2)的主要生理生化特性的表现(表1)看出, 两亲本之间8个指标均达极显著差异, 除陇油7号(P1)的MDA含量小于陇油9号(P2)外, 其余指标值均表现为P1大于P2。亲本陇油7号和陇油9号的SOD活性分别为318.49和240.57 U/mg, F2群体SOD活性出现较大变异, 在40.37~325.34, 平均为254.35 U/mg。F2群体的POD活性变异范围在18.35~187.55 U/g, 介于双亲之间, 平均为63.70 U/g。F2群体的平均CAT活性为8.73 U/g, 高于双亲值, 变异范围为1.30~27.05 U/g。F2群体的平均MDA含量为23.22 μ mol/g, 介于双亲之间, 变异范围在9.98~48.36 μ mol/g。游离脯氨酸含量在双亲之间存在极显著差异, 而且在F2群体中变化也较大, 为95.94~906.01 μ g/g。两亲本的可溶性蛋白含量相差约3.5 mg/g, F2群体的可溶性蛋白含量平均值介于双亲之间, 平均值为14.68 mg/g, 变异范围在8.08~21.90 mg/g。越冬率统计结果P1为95.2%, P2为45.8%, F2群体的越冬率为66.1%。陇油7号(P1)幼苗生长习性为匍匐性或半匍匐性, 陇油9号(P2)的幼苗习性主要表现为直立型或半直立型, 而在F2群体中出现了性状分离, 变异范围在1~4级。
2.2 F2群体主要生理生化特性的分布
如图1所示, 以各生理指标分布范围为横坐标, 频率为纵坐标, 以F2群体同一指标的次数分布进行作图, 可以看出, F2群体间存在着较大的遗传差异, 各生理指标的测定值均呈不同程度的连续分布, 它们的偏度和峰度见表1, 除SOD活性的偏度外都小于1, 分布频率大致接近正态分布, 且存在明显的双向超亲分离。大多数指标在F2群体中的平均值介于双亲之间, 只有CAT活性和游离脯氨酸含量在F2群体中的平均值超过双亲。
| 表1 亲本和F2群体的主要生理指标表现及分布 Table 1 Performance and distributions of physiological indicators in the F2 population and its parents |
2.3 F2群体的聚类分析
利用 DPS 对各个性状值进行标准化转换后, 计算欧氏遗传距离, 进行 UPGMA 聚类分析(图2)。从图中可以看出, 在聚类距离约为 4.78 处, F2群体分为 3个类群, 其中第1亚群包括38个F2个体, 第2亚群包含64个F2个体, 第3亚群包含1个F2个体, 在聚类距离为 3.58处, F2群体分为 8个类群, 其中第1亚群包括29个F2个体, 第2亚群包含9个F2个体, 第3亚群包含56个F2个体, 第4亚群包含1个F2个体, 第5亚群包含2个F2个体, 第6亚群包含4个F2个体, 第7亚群包含1个F2个体, 第8亚群包含1个F2个体。
2.4 F2单株的主要生理生化特性与抗寒性相关分析
本研究中, 白菜型冬油菜F2群体的各个指标间不是相互独立的, 而是存在着一定的相关关系(表2)。从表中可以看出, 越冬率与SOD活性、POD活性、CAT活性、游离脯氨酸含量和可溶性蛋白含量呈显著或极显著正相关, 相关系数分别为0.512, 0.472, 0.444, 0.550, 0.250; 与幼苗习性和MDA含量呈极显著负相关, 其相关系数为-0.863和-0.562, 说明白菜型冬油菜在低温胁迫下通过增加SOD活性、POD活性、CAT活性、游离脯氨酸含量和可溶性蛋白含量, 降低MDA含量, 提高抗寒性。由表2还可以看出, 白菜型冬油菜所有指标与越冬率的相关系数均达到显著或极显著水平, 说明幼苗习性, SOD活性, POD活性, CAT活性, MDA含量, 游离脯氨酸含量和可溶性蛋白含量在一定程度上均可作为白菜型冬油菜单一指标进行抗寒性鉴定[34]。
| 表2 越冬率与相关指标在F2群体中的相关分析 Table 2 Correlation analysis among overwintering rates and related indicators in the F2 population |
2.5 F2群体主要生理生化特性的回归分析
对幼苗习性, SOD活性, POD活性, CAT活性, MDA含量, 游离脯氨酸含量, 可溶性蛋白含量等7个性状, 通过SAS用全子集法计算所有可能的回归[35], 经过共线性诊断后挑出最好的回归结果。Y=0.899-0.299X1+0.001X2+0.001X3+0.015X4-0.01X5+0.014X6。其中Y为越冬率, X1为幼苗习性, X2为SOD活性, X3为POD活性, X4为CAT活性, X5为MDA含量, X6为可溶性蛋白含量。其中, X1, X5对Y的偏回归系数分别为-0.299, -0.01, 与越冬率呈负相关, 该方程R=0.912* * 。
2.6 F2群体主要生理生化特性的通径分析
2.6.1 主要生理生化指标的直接通径关系
表3为7个指标对越冬率影响的通径系数, 从表中可以看到各指标对越冬率的直接通径系数从大到小分别为:-0.689, 0.304, -0.293, 0.218, 0.176, 0.138, 0.115(正号是指促进作用, 负号是指抑制作用)。由此可以看出, 幼苗习性和SOD活性对越冬率的直接影响较大, CAT活性和可溶性蛋白含量对越冬率的直接影响较小。
| 表3 各指标对越冬率的通径分析 Table 3 Path analysis of overwintering rates and its indicators |
2.6.2 主要生理生化指标的间接通径关系
从表3中可以看出, 各指标对越冬率的总间接通径系数分别为:-0.1706, 0.2070, 0.2960, 0.3052, -0.2685, 0.3319, 0.1347。通过与直接通径系数相比较, 可以看出, 幼苗习性对越冬率的相关系数为r=-0.863, 直接通径系数为-0.689, 可见, 幼苗习性对越冬率主要是通过直接影响起作用, 而通过其他指标的间接影响较小。CAT活性与越冬率的相关系数为r=0.444, 间接通径系数为0.3052, 可以看出, CAT活性对越冬率的影响主要是通过间接影响起作用的。其余各指标对越冬率的影响均是通过直接和间接影响共同起作用的, 其中, SOD活性通过POD活性对越冬率的间接影响较大, 间接通径系数分别为0.0630; POD活性通过SOD活性对越冬率的间接影响较大, 间接通径系数为0.1088; MDA含量通过游离脯氨酸含量对越冬率的间接影响最大, 间接通径系数为-0.0983; 脯氨酸含量通过MDA含量对越冬率的间接影响最大, 间接通径系数为0.1321; 可溶性蛋白含量通过SOD活性和游离脯氨酸含量对越冬率的间接影响最大, 间接通径系数分别为0.0608和0.0606。
3 讨论
本文对103份冬油菜F2群体的SOD活性、POD活性和CAT活性等指标进行分析, 系统地评价了各指标在群体中的表现。试验结果发现, F2群体的6个指标均呈不同程度连续变异, 分布频率大致接近正态分布(图1), 表现出数量性状遗传的特点。聚类分析中, 在聚类距离为 4.78 处, F2群体分为 3个类群, 第1亚群是未能安全越冬植株, 第2亚群和第3亚群为基本可以安全越冬植株, 比例约为1∶ 2, 与F2群体越冬情况的田间统计值较一致; 在聚类距离为 3.58 处, F2群体分为 8个类群, 以聚类结果来看, F2群体间存在着较大的遗传差异, 例如在第2亚群中, 此亚群包含9个F2个体, 该亚群的性状中包含的幼苗习性表现为直立型, SOD活性、POD活性、CAT活性和游离脯氨酸含量等4个指标接近最小值, MDA含量接近最大值; 在第8亚群中, 此亚群只包含1个F2个体, 但该亚群的性状中包含的幼苗习性表现为匍匐型, 另有SOD活性、POD活性、CAT活性和游离脯氨酸含量等4个指标接近最大值, MDA含量接近最小值。
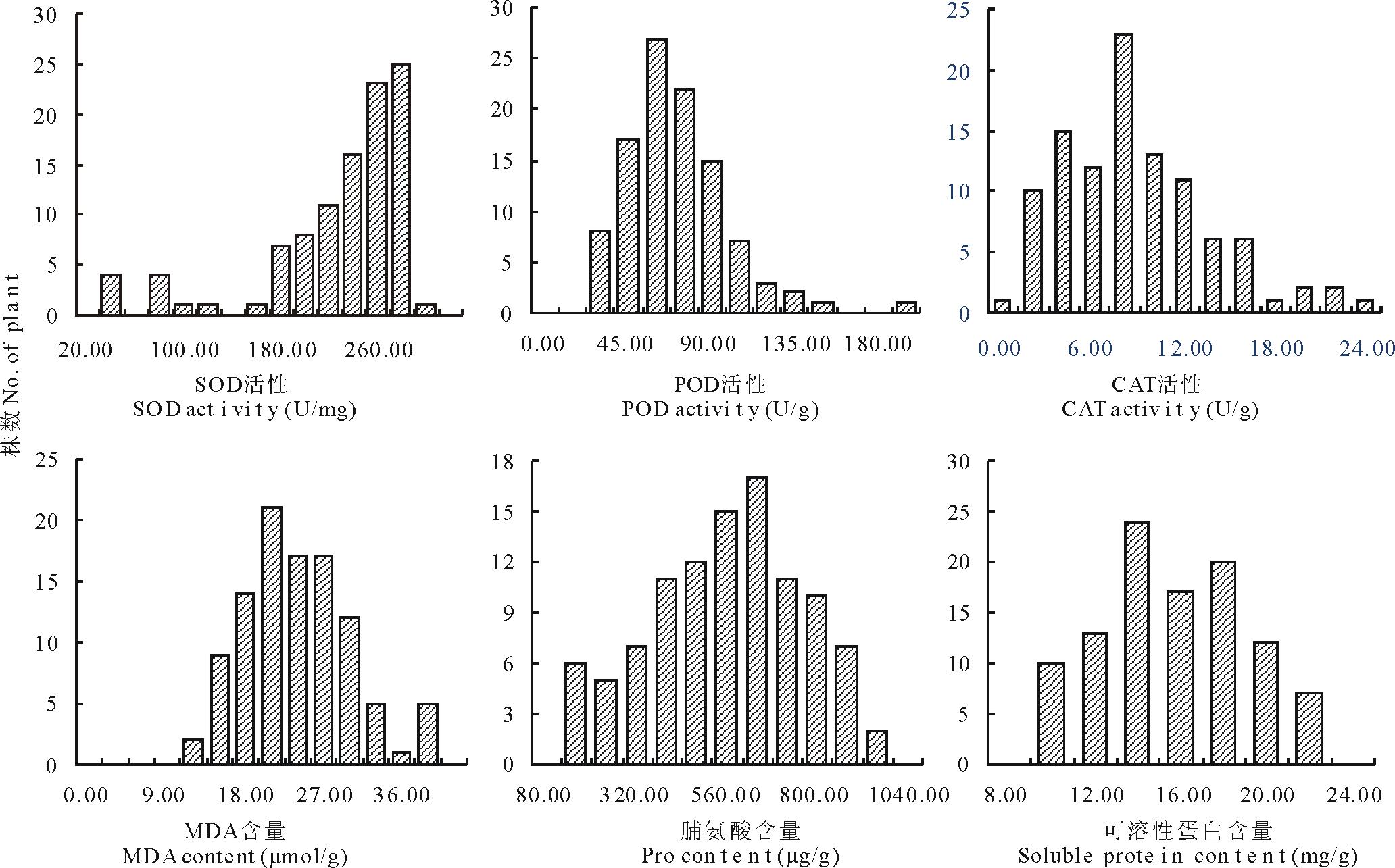 | 图1 F2群体相关指标的次数分布Fig.1 Frequency distribution of related indicators for F2 population |
在本研究中, 通过对白菜型冬油菜F2群体的供试指标间进行相关分析, 发现各性状间不是相互独立的, 而是存在着一定的相关关系。结果分析表明, 白菜型冬油菜F2群体的越冬率与幼苗习性、MDA含量间呈极显著负相关, 与SOD活性、POD活性、CAT活性、游离脯氨酸含量、可溶性蛋白含量呈极显著或显著性正相关, 可见, 幼苗越匍匐, 低温逆境下MDA积累少, 膜质过氧化程度小, 冬油菜的越冬情况越好; 反之, 幼苗越直立, MDA含量较高, 低温逆境下MDA积累多, 膜质过氧化程度大, 越冬情况越差, 与此同时, 植株通过增加SOD活性、POD活性、CAT活性以及低温逆境下积累游离脯氨酸含量和可溶性蛋白的含量增强其抗寒性。这与之前的研究结果较一致[8, 36, 37, 38, 39, 40, 41, 42, 43, 44]。
通径系数就是标准化的偏回归系数, 表示变数间的直接关系。从所得通径系数的大小可以知道自变数对依变数作用的大小, 通过对白菜型冬油菜F2群体各指标的通径分析, 发现各生理生化指标对越冬率的相关系数与直接通径系数方向一致, 但通径系数更能体现其本质, 因此得出各指标对白菜型冬油菜越冬率相对重要性从大到小的顺序是:幼苗习性, SOD活性, MDA含量, 游离脯氨酸含量, POD活性, CAT活性, 可溶性蛋白含量; 除直接通径系数外的通径系数为间接通径系数, 它反映的是某一性状对越冬率的影响是通过对其他组分的影响而间接影响到越冬率的, 幼苗习性对越冬率主要是通过直接影响起作用, 影响率达79.8%, CAT活性对越冬率的影响主要是通过间接影响起作用的, 间接影响率达68.7%, 而其余各指标对越冬率的影响均是通过直接和间接影响共同起作用的。通径分析更直观、具体的解释了各生理生化特性之间的相互关系。
Zhu等[45]认为性状间显著相关暗示相应性状的部分QTL间可能存在连锁或一因多效。所以对上述生理生化指标进行相关分析可以进一步了解指标间的相互关系, 从而为今后的白菜型冬油菜抗寒性遗传分析提供科学依据。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|