{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
16种野豌豆属植物花粉形态的扫描电镜观察
[刘鹏 , 马利超, 王宇, 刘志鹏
, 马利超, 王宇, 刘志鹏* ]
, 马利超, 王宇, 刘志鹏]
|
|


作者简介:刘鹏(1989-),女,甘肃兰州人,在读硕士。E-mail:liupeng1989.happy@163.com
野豌豆属牧草适宜在高寒和高海拔地区栽培,是我国高原牧区重要的蛋白饲料来源。该属有190个物种,4个亚属: Vicia, Cracca, Ervum和 Faba。本研究从中国、美国、土耳其和叙利亚等11个国家收集了16份野豌豆属牧草种质,利用扫描电镜对这16份材料的花粉形态进行观察。结果表明,野豌豆属植物花粉均以单粒形式存在,极面观为三裂圆形和三角形,赤道面观为扁圆形、长椭圆形和椭圆形。三沟萌发孔,沟长深入两端;极轴长为(9.60±1.52)(30.40±2.07) μm,赤道轴长为(11.80±0.84)(38.20±0.84) μm。花粉的表面纹饰呈3种类型:网状纹饰,脑状纹饰和疣状纹饰。网状纹饰根据外壁纹饰疏松程度不同,又可分为2种亚类型:粗网状纹饰和细网状纹饰。本研究较为系统地观察了16个野豌豆属牧草的花粉形态,探讨了其系统分类学特性,为揭示野豌豆属种质资源的遗传多样性奠定了基础。
Plants of the genus Vicia are readily cultivated in alpine and high-altitude areas, such as the Tibetan plateau in China, where they are used as a source of protein. There are about 190 species in this genus, and four subgenera: Vicia, Cracca, Ervum and Faba. We studied 16 Vicia accessions sourced from China, USA, Turkey, Syria, and 7 other countries. The pollen morphology of the 16 accessions was observed by scanning electron microscopy. Pollen of Vicia is single-grained, with three lobes and a triangular outline in polar view. The pollen studied ranged in shape from prolate-spheroidal to prolate andoblate in equatorial view with three channel apertures reaching both poles. Polar axis length was (9.60±1.52)-(30.40±2.07) μm, and equatorial axis length was (11.80±0.84)-(38.20±0.84) μm. Three types of pollen surface ornamentation were observed: reticulate, brain-like and warty. Furthermore, depending on the degree of protrusion, the reticulation type can be divided into coarse and fine reticulation sub-types. In this study,the pollen morphologies of 16 Vicia accessions were observed in detail, and their taxonomic characteristics are discussed as a basis for categorising the diversity in the Vicia genus.
野豌豆属(Vicia)植物是一种优良的牧草, 具有很好的防风固沙保水作用[1]。因其具有生物产量高[2]、生育周期短[3]、种子粗蛋白含量高[4, 5]和抗寒性较强等特点[6], 而被科技工作者广泛关注。对野豌豆属植物的花粉生物学特性研究, 可为植物物种及其种质资源鉴定提供重要的形态理论依据。野豌豆属植物约有190个物种[7], Ball[8]将该属划分成4个亚属:Vicia, Cracca, Ervum和Faba。随后研究人员进一步分析了Vicia和Vicilla两个亚属的生物学特性[9, 10, 11]。Maxted[12]在原表型分类的基础上又将Vicia亚属细分为9大系列, 38个物种, 14个亚种。本属植物主要分布在北半球温带地区, 拉丁美洲有零星分布[13]。根据中国数字植物标本馆(http://www.cvh.org.cn/cms/)和宋敏等[14]的统计结果, 在我国本属植物广布于全国各省, 华北、西北、西南较多, 其中栽培种5种, 野生种36种, 华南和台湾地区分布很少, 仅有野生种1种。
关于野豌豆属植物花粉形态的研究, 国内学者曾开展过一些工作。苏亚拉图等[15]使用扫描电镜观察了我国内蒙古境内野豌豆属13种植物的花粉结构, 发现野豌豆属植物的花粉形态、萌发孔和外壁纹饰等特征在种内是稳定的, 但该研究的取材地域存在一定的局限性。苏亚拉图等[16]研究发现野豌豆属新变种狭柳叶野豌豆(V. venosa var. ich-hsinganica)的花粉形态为长球形, 具三孔沟, 沟缘加厚, 孔膜高升, 沟浅, 外壁具不规则网状纹饰。Erdtman[17]认为利用花粉粒形态特征可以提高植物分类的精确度。深入观察花粉的形态特征可为植物分类学和授粉生物学等研究提供有价值的参考信息[18, 19]。虽然很多学者在野豌豆属植物花粉形态上做了一定研究工作, 但仍存在一些不足, 如物种数目较少、来源地单一、未使用高分辨率电镜等。本研究利用高分辨率扫描电镜, 较为系统地对来自11个国家的16份野豌豆属物种的花粉形态进行了观察, 探讨其系统分类学特性, 为野豌豆属植物的孢粉学相关研究积累了相关信息[20]。
野豌豆属植物种子由美国国家种质资源库(NPGS)提供。2012年8月初, 于兰州大学榆中校区智能温室种植(16 h光照, 8 h黑暗; 光照时22℃, 黑暗时20℃)。在当年10-11月份盛花期分别采集成熟花药作为扫描电镜观察的材料。材料具体名称及来源地见表1。
| 表1 16种野豌豆属物种名称及其来源 Table 1 Name and origin of sixteen Vicia accessions |
植物盛花期进行取样, 每个物种采集5朵鲜花, 用镊子将花药取出, 放入1.5 mL EP管中, 常温下干燥48 h后存放于4℃冰箱中。为了获得自然真实的表面纹饰, 采用自然散出的花粉扫描, 个别材料的花粉是从刺破的成熟花药中释放得到, 扫描前均未经任何试剂处理[21]。在导电胶上将花粉均匀散布, 置于金属载物台上, 用常规真空喷镀法喷金90 s后, 置于扫描电镜下观察, 选取有代表性的视野, 分别进行显微拍摄。
每份材料随机取30粒花粉进行大小的测量和统计。取平均值和变异系数表示变化幅度。花粉粒大小以极轴长乘以赤道轴长(P× E)来表示, 其形状以极轴长和赤道轴长的比值(P/E)来表示。花粉的表面纹饰形态特征用Punt等[22]文章中的专业术语来描述。P/E在0.40.8为扁圆形, P/E在1.02.0为椭圆形, P/E在0.81.0为长椭圆形。
| 表2 16份野豌豆属植物花粉形态学特征 Table 2 Main characteristics of the pollen morphology in sixteen accessions of Vicia |
对16种野豌豆属植物的花粉进行扫描电镜观察, 发现野豌豆属植物的花粉为单粒花粉, 形状是扁圆形的9种(V. aintabensis、V. galeata和V. costata等)、长椭圆形的5种(V. sativa subsp. cordata、V. sativa subsp. amphicarpa和V. lutea等)和椭圆形的2种(V. narbonensis和V. monantha)(表2)。对花粉粒大小的比较发现, 不同物种花粉粒大小差异较明显, V. monantha极轴长显著大于其他物种(P< 0.05)。V. costata赤道轴长显著大于其他物种(P< 0.05)。此外, 野豌豆属植物花粉外壁表面纹饰可分为网状纹饰(V. monantha和V. ervilia等11种)、脑状纹饰(如V. aintabensis和V. grandiflora等4种)和疣状纹饰(V. articulate)3种类型, 其极面观为三裂圆形和三角形(仅V. grandiflora)。萌发孔沟接近两极且宽窄不一, 沟内光滑或有疣状凸起。
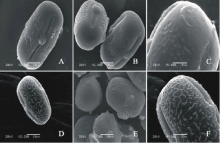
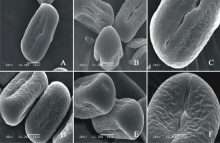
V. aintabensis(图1AC)极轴长(24.40± 2.77) μ m, 赤道轴长(35.40± 0.89) μ m, P/E值为0.69。赤道面观为扁圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 前者萌发沟深且宽, 沟内有细小的疣状凸起。表面为脑状纹饰, 较浅。
V. articulata(图1DF)极轴长(13.60± 1.34) μ m, 赤道轴长(17.20± 1.48) μ m, P/E值为0.79。赤道面观为扁圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 前者萌发沟深且宽, 沟内有细小的疣状凸起。表面为疣状纹饰, 明显。
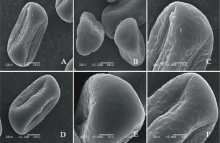
V. galeata(图2AC)极轴长(21.60± 1.14) μ m, 赤道轴长(34.20± 0.84) μ m, P/E值为0.63。赤道面观为扁圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟均较深。表面为脑状纹饰, 较深, 比较明显。
V. grandiflora(图2DF)极轴长(22.80± 1.79) μ m, 赤道轴长(29.20± 1.48) μ m, P/E值为0.78。赤道面观为扁圆形, 极面观为三角形。三孔萌发沟并延伸达两极, 萌发沟均较深。表面为脑状纹饰, 比较模糊。
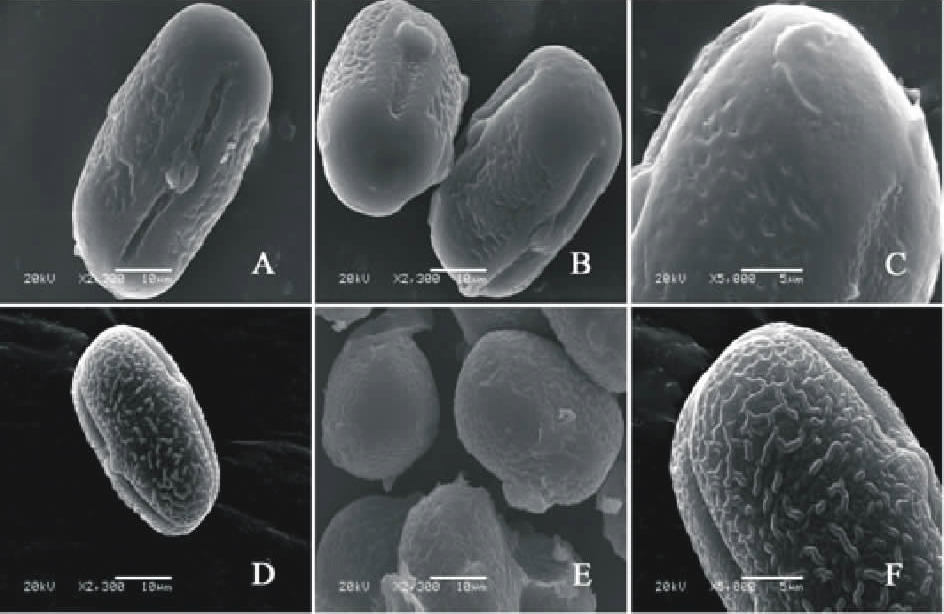 | 图1 V. aintabensis(AC)和V. articulate(DF)花粉粒形态(图A、B、D和E中的图例为10 μ m, 图C和F中的图例为5 μ m。)Fig.1 Pollen morphology of V. aintabensis (A-C) and V. articulata (D-F)(The scale bars in A, B, D and E is 10 μ m, and in C and F is 5 μ m.) |
 | 图2 V. galeata(AC)和V. grandiflora (DF)花粉粒形态(图A、B和D中的图例为10 μ m, 图C、E和F中的图例为5 μ m。)Fig.2 Pollen morphology of V. galeata (A-C) and V. grandiflora (D-F)(The scale bars in A, B, D and E is 10 μ m, and in C and F is 5 μ m.) |
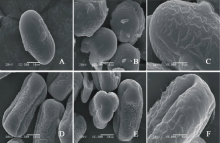
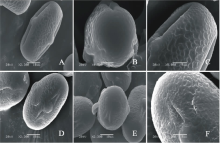
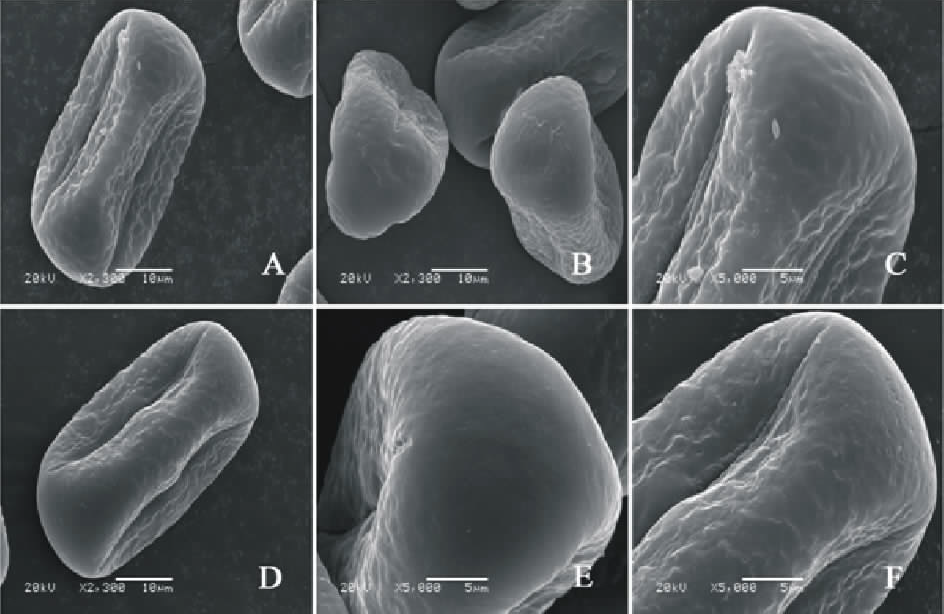
V. benghalensis(图3AC)极轴长(21.40± 1.14) μ m, 赤道轴长(21.60± 1.67) μ m, P/E值为0.99。赤道面观为长椭圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟细窄。表面为网状纹饰, 较浅。网眼大小不一。
V. costata(图3DF)极轴长(17.80± 2.17) μ m, 赤道轴长(38.20± 0.84) μ m, P/E值为0.47。赤道面观为扁圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟细窄。表面为脑状纹饰, 明显。
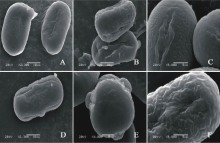
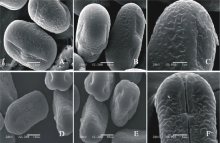
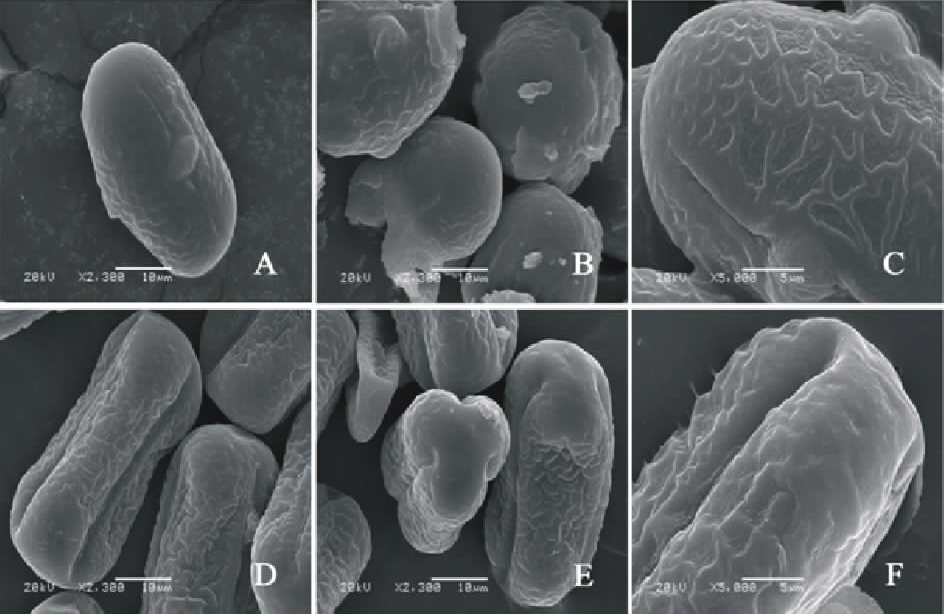
V. sativa subsp. amphicarpa(图4AC)极轴长(17.80± 1.22) μ m, 赤道轴长(21.00± 1.22) μ m, P/E值为0.85。赤道面观为长椭圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟较宽, 沟内有细小的疣状凸起。表面为细网状纹饰, 较浅。网眼大小不一。
V. sativa subsp. cordata(图4DF)极轴长(9.60± 1.52) μ m, 赤道轴长(11.80± 0.84) μ m, P/E值为0.81。赤道面观为长椭圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟细窄。表面为网状纹饰, 明显。
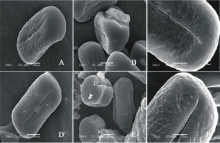
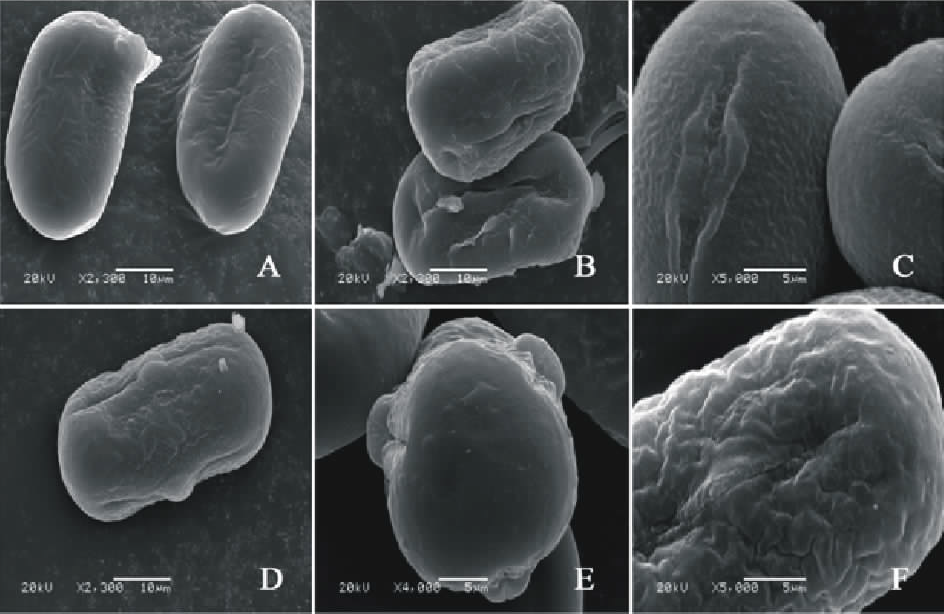
V. monantha(图5AC)极轴长(30.40± 2.07) μ m, 赤道轴长(16.00± 2.24) μ m, P/E值为1.90。赤道面观为椭圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟细窄。表面为网状纹饰, 比较模糊。
V. narbonensis(图5DF)极轴长(24.40± 1.34) μ m, 赤道轴长(19.60± 1.67) μ m, P/E值为1.24。赤道面观为椭圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟较狭窄。表面为网状纹饰, 较浅。网眼大小不一。
V. ervilia(图6AC)极轴长(17.80± 1.58) μ m, 赤道轴长(20.80± 1.30) μ m, P/E值为0.85。赤道面观为长椭圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟细窄。表面为网状纹饰, 明显。网眼大小不一。
 | 图3 V. benghalensis(AC)和V. costata (DF)花粉粒形态(图A、B、D和E中的图例为10 μ m, 图C和F中的图例为5 μ m。)Fig.3 Pollen morphology of V. benghalensis (A-C) and V. costata (D-F)( The scale bars in A, B, D and E is 10 μ m, and in C and F is 5 μ m.) |
 | 图4 V. sativa subsp. amphicarpa(AC)和V. sativa subsp. cordata(DF)花粉粒形态(图A、B和D中的图例为10 μ m, 图C、E和F中的图例为5 μ m。)Fig.4 Pollen morphology of V. sativa subsp. Amphicarpa (A-C) and V. sativa subsp. cordata (D-F)( The scale bars in A, B and D is 10 μ m, and in C, E and F is 5 μ m.) |
 | 图5 V. monantha(AC)和V. narbonensis (DF)花粉粒形态(图A、B、D和E中的图例为10 μ m, 图C和F中的图例为5 μ m。)Fig.5 Pollen morphology of V. monantha (A-C) and V. narbonensis (D-F)( The scale bars in A, B, D and E is 10 μ m, and in C and F is 5 μ m.) |
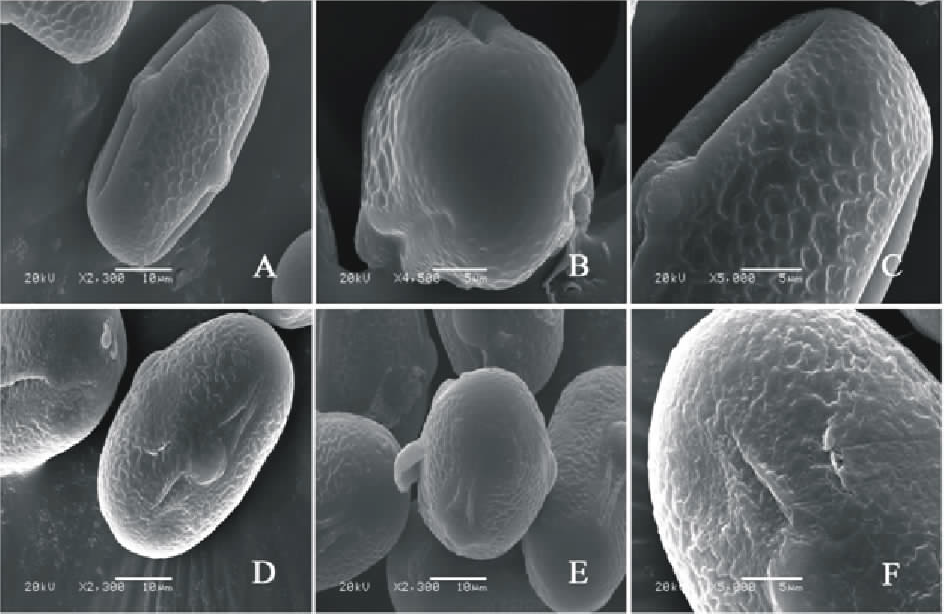 | 图6 V. ervilia(AC)和V. lutea(DF) 花粉粒形态(图A、D和E中的图例为10 μ m, 图B、C和F中的图例为5 μ m。)Fig.6 Pollen morphology of V. ervilia (A-C) and V. lutea (D-F)( The scale bars in A, D and E is 10 μ m, and in B, C and F is 5 μ m.) |
V. lutea(图6DF)极轴长(25.00± 1.58) μ m, 赤道轴长(31.00± 1.58) μ m, P/E值为0.80。赤道面观为长椭圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟较宽, 沟内有细小的疣状凸起。表面为细网状纹饰, 较浅。
V. hybrid(图7AC)极轴长(25.80± 0.84) μ m, 赤道轴长(36.00± 0.84) μ m, P/E值为0.71。赤道面观为扁圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟较宽, 沟内有细小的疣状凸起。表面为网状纹饰, 较浅。网眼大小不一。
V. hyrcanica(图7DF)极轴长(20.40± 1.67) μ m, 赤道轴长(34.80± 1.48) μ m, P/E值为0.59。赤道面观为扁圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟细窄。表面为网状纹饰, 明显。
V. peregrine(图8AC)极轴长(24.40± 1.34) μ m, 赤道轴长(37.00± 1.58) μ m, P/E值为0.65。赤道面观为扁圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟细窄, 沟内有细小的疣状凸起。表面为网状纹饰, 较浅。网眼大小不一。
V. michauxi(图8DF)极轴长(21.60± 1.58) μ m, 赤道轴长(36.60± 1.67) μ m, P/E值为0.59。赤道面观为扁圆形, 极面观为三裂圆形。三孔萌发沟并延伸达两极, 萌发沟细窄。表面为细网状纹饰, 明显。
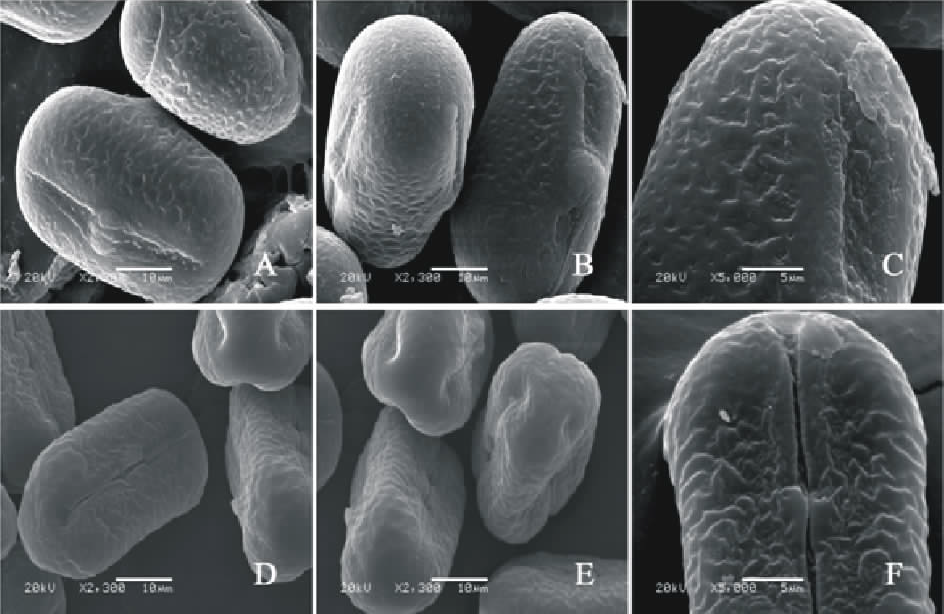 | 图7 V. hybrid(AC)和V. hyrcanica(DF)花粉粒形态(图A、B、D和E中的图例为10 μ m, 图C和F中的图例为5 μ m。)Fig.7 Pollen morphology of V. hybrid (A-C) and V. hyrcanica (D-F)( The scale bars in A, B, D and E is 10 μ m, and in C and F is 5 μ m.) |
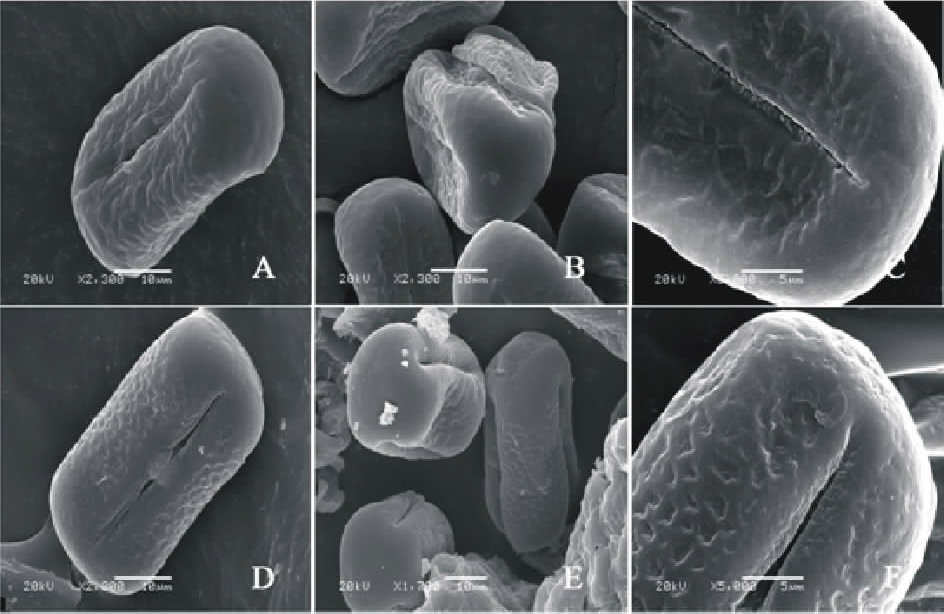 | 图8 V. peregrine(AC)和V. michauxi(DF)花粉粒形态(图A、B、D和E中的图例为10 μ m, 图C和F中的图例为5 μ m。)Fig.8 Pollen morphology of V. peregrine (A-C) and V. michauxi (D-F)( The scale bars in A, B, D and E is 10 μ m, and in C and F is 5 μ m.) |
张庭龙等[23]认为花粉粒大小可能与植物生长环境有关, 花粉形态要比花粉大小更加稳定。从表2可以看出, 16种野豌豆属植物花粉大小差异明显, 从单一性状指标难以看出进化关系。16种野豌豆属植物花粉的赤道面观为扁圆形的9种(V. aintabensis、V. articulate和V. costata等)、长椭圆形的5种(V. sativa subsp. amphicarpa、V. ervilia和V. lutea等)和椭圆形2种(V. narbonensis和V. narbonensis)。从萌发沟来看, 均为三孔萌发沟, 萌发沟直达两极。说明花粉的形态比其大小稳定。
有研究资料表明, 可将花粉形态特征作为植物的分类依据[24]。花粉作为植物的保守结构, 其形态特征具有可遗传性, 同时具有科属的共同特征和种的特异性[25]。野豌豆属植物花粉均为三孔萌发沟, 沟长直达两极, 但每个物种之间有差异, 如V. sativa subsp. amphicarpa沟较深, 沟内有疣状凸起, 这一特性为我们鉴别该物种提供了重要线索。从表面纹饰分析, V. benghalensis等11份植物材料为网状纹饰, 在参试材料中比较普遍。V. articulate表面为疣状纹饰, V. costata花粉表面纹饰为脑状纹饰, 说明每个物种都有自己独有的特点, 其形态特征具有种的特异性, 这为鉴定物种的特异性提供了依据[26]。
本文通过观察16种野豌豆属材料的花粉形态特征发现萌发孔沟均为三孔沟, 极面观为三裂圆形(除V. grandiflora极面观为三角形), P/E值均小于1(除V. monatha), 说明野豌豆属物种间花粉粒萌发孔的类型以及极面观等方面相似性较高, 这与苏亚拉图等[15]观察得出的内蒙古野豌豆属植物花粉的形状、萌发孔及外壁纹饰等特征在种内具有稳定性一致, 表明该属的花粉形态保守性较高。但是, 每个物种的赤道轴和极轴长大小不一, 赤道轴长从大到小排列, 即V. costata、V. peregrine、V. michauxi、V. hybrid、V. aintabensis、V. hycanica、V. galeata、V. lutea、V. grandiflora、V. benghalensis、V. sativa subsp. amphicarpa、V. ervilia、V. narbonensis、V. articulate、V. monantha、V. sativa subsp. cordata。极轴长从大到小排列, 即V. monantha、V. galeata、V. narbonensis、V. peregrine、V. aintabensis、V. grandiflora、V. galeata、V. benghalensis、V. michauxi、V. hycanica、V. hybrid、V. costata、V. ervilia、V. sativa subsp. amphicarpa、V. sativa subsp. cordata。这对鉴定野豌豆属亲缘关系方面具有重要的分类学价值。
Beck[27]指出植物花粉在原始时期都是光滑的且具有无定型的外壁, 演化趋势为表面光滑→ 表面具小穴、小沟状雕纹→ 粗糙的、具疣状纹饰→ 表面颗粒状→ 表面网状。16种野豌豆属植物的花粉外壁纹饰形态特征所表现的差异表明它们的进化程度不同, 存在物种多样性。根据Beck[27]进化理论得出:V. aintabensis、V. galeata、V. grandiflora、V. costata花粉表面纹饰属于脑状纹饰, 比较模糊, 较原始; 而V. benghalensis、V. sativa subsp. cordata、V. monantha、V. narbonensis、V. ervilia、V. lutea、V. hybrid、V. hycanica、V. michauxi、V. peregrine、V. sativa subsp. amphicarpa外壁纹饰为网状纹饰, 较进化。V. articulate的进化程度介于两者之间。从花粉形态学角度探讨野豌豆属植物系统进化具有重要意义, 所观察的16个物种表现了野豌豆属植物的演化规律, 反应了野豌豆属的共同特征, 为野豌豆属牧草系统分类学奠定了基础, 为孢粉学相关研究积累信息。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [1] |
|
| [2] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [20] |
|
| [21] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|