
{kind=link}
{kind=link}
{kind=link}
{kind=link}
荒漠植物红砂根系对干旱胁迫的生理响应
[种培芳 , 李航逸, 李毅
, 李航逸, 李毅* ]
, 李航逸, 李毅]
|
|


作者简介:种培芳(1977-),女,甘肃永登人,副教授,博士。E-mail:zhongpf@gsau.edu.cn
以抗旱性不同的红砂2个种源2年生苗木为试材,通过盆栽实验,测定干旱胁迫后红砂苗木根系的活力、膜透性(RPP)、脯氨酸 (Pro)、可溶性糖 (SS)、可溶性蛋白 (SP)、丙二醛(MDA)含量和抗氧化物酶系(SOD、POD、CAT)的活性变化,并通过灰色关联度法分析了各指标与红砂根系抗旱性的关系。结果表明,水分胁迫导致红砂根系活力下降,膜透性RPP和膜质过氧化产物MDA 含量增加,渗透调节物质Pro含量、SS含量和SP含量明显增加,抗衰老酶SOD、POD和CAT活性显著升高,且抗性强的种源比抗性弱的种源对干旱胁迫的生理生化反应更明显。红砂根系的RPP、Pro和根系活力的关联度虽然都大于0.9,但红砂根系并不是完全通过这几种机理适应干旱,而是发生了一系列适应性的生理变化,以提高其根系的抗旱性。
The physiological responses of seedling roots to drought stress were investigated in the desert plant Reaumuria soongorica from two provenances: highly drought resistant (Wuwei) and weakly drought resistant (Lanzhou). Under drought stress, root vigor, relative permeability of plasma (RPP), contents of proline (Pro),soluble sugars (SS),soluble protein (SP),malondialdehyde (MAD) and activities of superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT)were measured with two-year nursery stocks. Grey correlative analysis was used to analyse drought resistance and its correlation with indexes of roots. Root vigor decreased significantly under drought stress, while RPP and the contents of MDA, Pro, SS and SP, the activities of SOD, POD and CAT of roots increased significantly. Germplasm of highly drought-resistant provenance (Wuwei) was more sensitive to drought stress than germplasm of weakly drought-resistant provenance (Lanzhou). The grey correlative degrees for root vigor, RPP and Pro and drought resistance were greater than 0.9. However, R. soongorica enhanced drought resistance through a wider series of physiological variations, beyond these three.
根系与植物的耐旱性关系十分密切, 是决定其生物量和抗旱性的重要方面。根系是植物吸收水分的主要部位, 干旱来临时其最先感知, 迅速产生化学信号向地上部传递, 促使气孔关闭以减少水分散失[1], 并通过自身形态和生理生化特征的调整以适应变化后的土壤水分环境[2, 3]。
红砂(Reaumuria soongorica)为柽柳科红砂属小灌木[4], 因其生态可塑性强, 具有很强的抗旱、耐盐和集沙能力[5, 6], 所以, 人们对红砂尤其是关于其茎、叶抗旱性的生理研究也越来越多[7, 8, 9, 10, 11, 12]。而对红砂根系的研究由于取样难度较大、研究技术和手段也比较缺乏等原因研究相对较少。现有的研究表明, 土壤干旱促进红砂根系伸长, 有利于对深层土壤水分养分的吸收利用[13]。并且不同土壤水分状况下红砂的根系构型表现出显著差异, 进而影响其对养分和水分的吸收能力[13]。那么红砂根系形态对水分的响应必然会引起相关的生理变化。本研究通过盆栽试验对不同干旱胁迫条件下荒漠植物红砂根系的生理特性进行了较为系统的研究, 以期为揭示红砂根系的抗旱机理及丰富其抗旱机制资料方面提供有力的参考依据。
本实验以抗旱性不同的红砂兰州(抗旱性弱)和武威(抗旱性强)种源2年生实生苗为材料[14], 籽种是2010年10 月初在兰州和武威采集的, 2011年3月上旬在甘肃农业大学林学院实验基地内盆栽土培。挑选生长均匀的两年生红砂苗木, 于2013年4月栽植于塑料桶中(高37 cm, 直径32 cm), 沙壤土, 每桶1株, 每个种源各60桶, 充分浇水。
待苗木全部缓苗成活后于5月17日开始进行干旱处理。设对照(CK, 土壤含水量21.76%23.81%)、轻度干旱胁迫(light drought, LD; 土壤含水量13.27%15.78%)、中度干旱胁迫(middle drought, MD; 土壤含水量9.49%10.54%)和重度干旱胁迫(high drought, HD; 土壤含水量4.32%5.38%)4个处理, 每个处理15桶(每5桶为1个重复, 重复3次)。为防止自然降雨对水分胁迫的影响, 在盆栽苗上方设遮雨棚, 每天称重并及时补充当天损失水分。
待每个处理达到设定土壤含水量并维持40 d后(7月底)开始取样, 每次随机取5 桶, 共5 株苗木(每隔7 d取一次样, 共取样3次)。将整株根系连同土壤从桶中取出, 小心去除根系表面的大块泥土, 置于冰盒中, 然后带回室内冲洗干净后, 置于冰箱中备用。每次取母根(平均直径为0.28 cm)样根鲜重约5 g进行生理指标分析。
根系活力(root vigor)采用 TTC 染色法测定[15]。细胞质膜透性(relative permeatbility of plasma, RPP)采用电导法测定[16]。脯氨酸(proline, Pro)含量采用茚三酮比色法测定[17]。可溶性糖(soluble sugur, SS)含量采用蒽酮乙酸乙酯比色法测定[18]。可溶性蛋白(soluble protein, SP)采用考马斯亮蓝比色法测定[16]。丙二醛(malondialdehyde, MDA)含量采用硫代巴比妥酸法测定[17]。超氧化物歧化酶(superoxide dismutase, SOD)活性采用核黄素-NBT法测定[18], 过氧化氢酶(catalase, CAT)活性采用紫外吸收法测定[17], 过氧化物酶(peroxidase, POD)活性采用愈创木酚比色法测定[17]。所有生理指标均3次重复测定。
灰色关联度分析方法已被广泛应用于许多研究领域中。设参考数列为X0, 比较数列为Xi(i=1, 2, …, n), 则参考数列X0={X0(1), X0(2), …, X0(n)}, 比较数列Xi={X1(1), X1(2), …, X1(n)}。利用下列公式对各指标进行处理:
用X'i(k)=Xi(k)-XiSi对原始数据进行无量纲化处理, 其中Xi(k)是原始数据, Xi和Si是同一指标的平均值和标准差。
ε i(k)=
ri=
式中, ε i(k)为关联系数; ri为灰色关联度; Δ i(k)=|X0(k)-Xi(k)|为X0数列与Xi数列在第k点的绝对值; minminΔ i(k)为二级最小差, maxmaxΔ i(k)为二级最大差; ρ 为分辨系数, 取值范围为01, 文中试验取值0.5[19]。权重计算公式:
W(k)=
式中, W(k)为第k个指标的权重值; R(k)为第k个指标的关联度。
采用 SPSS 13.0 软件进行方差分析, 然后进行LSD检验(α =0.05)。多重比较分析采用Duncan法。采用Excel 2003软件作图。
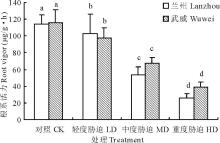
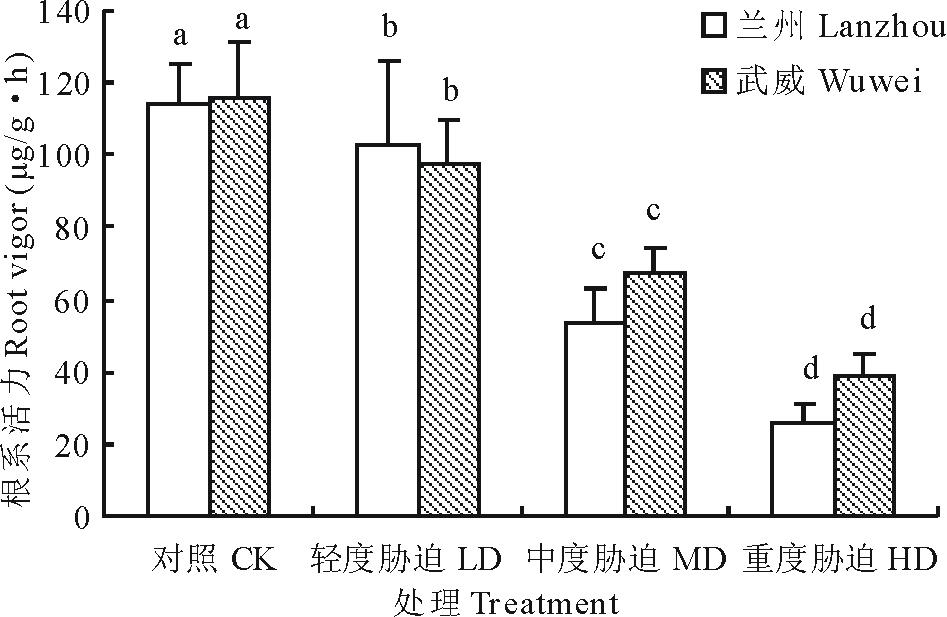 | 图1 干旱胁迫对红砂两个种源苗木根系活力的影响Fig.1 Effect of drought stress on root vigor of two provenances (Lanzhou and Wuwei) of R. soongorica seedlings |
不同字母表示差异显著(P< 0.05) Different letters mean significantly different at P< 0.05. 下同The same below.
由图1可知, 随着干旱胁迫的增加, 红砂两个种源苗木根系活力均呈下降趋势, 且各处理间根系活力差异均达显著水平(P< 0.05)。与对照相比, 兰州种源在轻度干旱、中度干旱和重度干旱胁迫下分别下降了12.71%, 47.57%和51.85%, 而武威种源分别下降了15.51%, 31.63%和41.79%。可以看出, 随着干旱程度的加剧, 抗旱性弱的兰州种源根系活力比抗旱性强的武威种源下降幅度大。
膜透性是表征细胞结构完整性的主要特征之一。研究结果显示, 干旱胁迫对红砂苗木根系的RPP变化产生显著影响(图2A)。与对照相比, 轻度干旱胁迫处理下兰州和武威种源的根系RPP为22%和26%, 显著高于对照(P< 0.05)。随着胁迫程度的加剧, 兰州种源根系RPP增加幅度高于武威种源的, 在中度胁迫时兰州种源增加幅度比武威种源多31%。在重度胁迫下, 两种源增加幅度虽有所减缓, 但兰州种源(46%)仍显著高于武威种源(40%)的。3个干旱处理与对照的差异在武威和兰州种源间均达到显著水平(P< 0.05)。
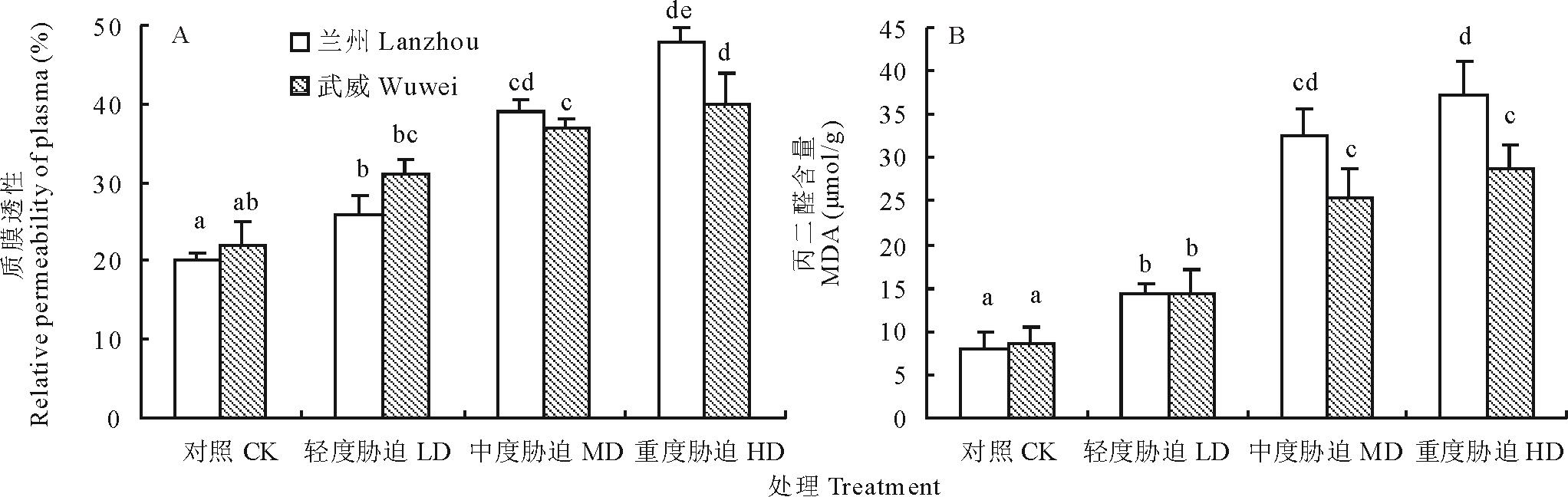 | 图2 干旱胁迫对红砂两个种源苗木根系质膜透性和丙二醛含量的影响Fig.2 Effect of drought stress on relative permeability of plasma and MDA content of two provenances (Lanzhou and Wuwei) of R. soongorica seedlings |
红砂两种源苗木根系的MDA含量随着干旱胁迫程度呈逐渐增加趋势(图2B), 在重度干旱胁迫下上升幅度最大, 分别比对照上升了 370.93%(兰州)和237.94%(武威), 达显著水平(P< 0.05)。可以看出, 兰州种源提高幅度明显大于武威种源, 说明在干旱胁迫下, 武威种源的细胞膜伤害程度轻于兰州种源, 这是因为其抗旱性强于兰州种源的缘故。
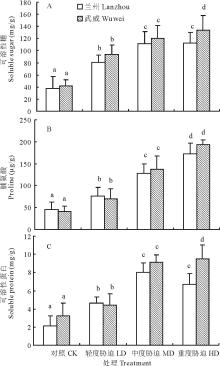
由图3可知, 干旱胁迫对红砂两个种源苗木根系的可溶性糖(SS)、脯氨酸(Pro)和可溶性蛋白(SP)含量均产生显著的影响。随干旱胁迫的加剧, 红砂根系SS含量(图3A)和Pro含量(图3B)均显著增加(P< 0.05), 尤其是从对照到中度胁迫, 增加幅度最大:兰州和武威种源根系SS含量从轻度到中度胁迫分别上升了38.27%和27.97%, 而从中度胁迫到重度胁迫仅上升了0.80%和10.88%。说明干旱胁迫可促进SS的合成, 但重度干旱胁迫时, 糖合成能力会有所下降, 尤其是抗性差的兰州种源比抗性强的武威种源这种能力减弱更多。与对照相比, 两种源的Pro含量在中度胁迫时分别增加了282.22%(兰州)和376.63%(武威), 达显著差异(P< 0.05)。 说明在遇到干旱胁迫时, Pro是降低红砂根系渗透势, 维持细胞膨压的重要渗透调节物质, 种源抗性越强, 这种作用越明显。
 | 图3 干旱胁迫对红砂两个种源苗木根系可溶性糖、脯氨酸和可溶性蛋白含量的影响Fig.3 Effect of drought stress on soluble sugar, proline and soluble protein content of two provenances (Lanzhou and Wuwei) of R. soongorica seedlings |
如图3C所示, 随着胁迫程度的增加, 兰州种源根系的SP含量呈先升高后降低的趋势。与对照相比, 轻度胁迫和中度胁迫下, SP含量增加, 最高可达8.04 mg/g。但随着胁迫程度的加剧, 在重度胁迫下, SP含量又降低至6.71 mg/g, 比中度胁迫时降低了16.54%。与对照相比, 武威种源根系的SP含量随干旱胁迫程度的加剧显著增加(P< 0.05)。轻度和对照, 中度和轻度间以及重度与中度间差异显著(P< 0.05)。
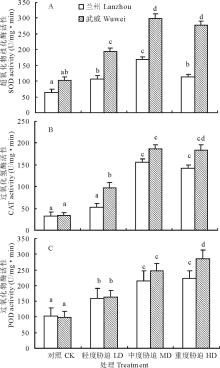
如图4所示, 红砂两种源苗木根系的3种保护酶(SOD, CAT, POD)活性在不同干旱处理下均受到显著影响。其中, SOD(图4A)和CAT(图4B)活性随胁迫程度的加剧均呈先上升后下降的趋势, 而POD (图4C)活性则呈持续升高的趋势。与对照相比, 兰州和武威种源幼苗根系的SOD 和CAT均在中度胁迫时达最大值, 分别比对照增加了159.20%和192.62%(SOD), 374.49%和455.19(CAT), 差异达显著水平(P< 0.05)。两种源苗木根系的POD均在重度胁迫时达最大值, 分别比对照增加了119.01%(兰州)和190.39%(武威), 也达显著水平(P< 0.05)。两种源间比较, 武威种源的SOD、CAT和POD活性增加幅度均比兰州种源的大, 尤其在重度胁迫下的差异均达显著水平(P< 0.05)。
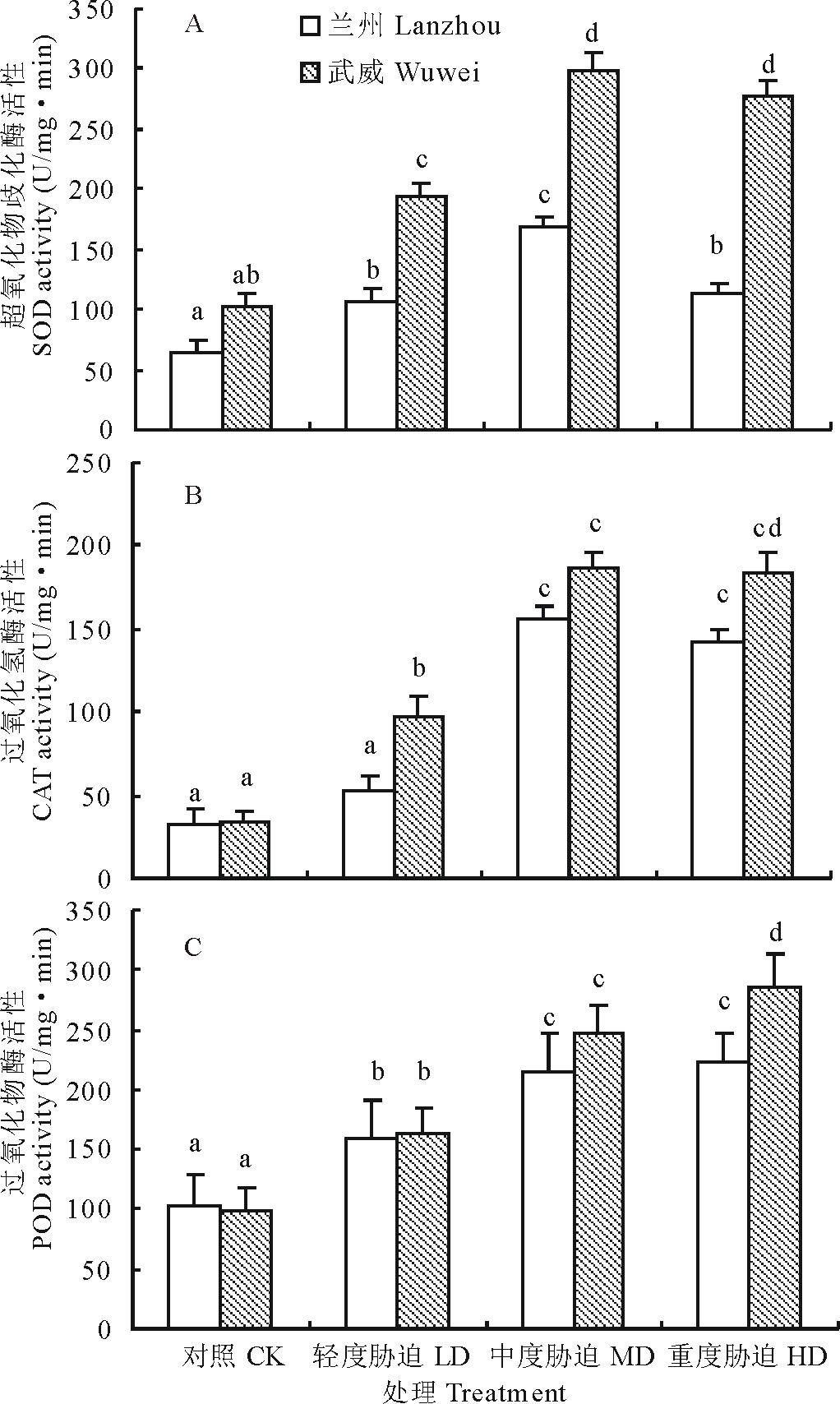 | 图4 干旱胁迫对红砂两个种源苗木根系SOD、CAT和POD活性的影响Fig.4 Effect of drought stress on SOD, CAT and POD activity of two provenances (Lanzhou and Wuwei) of R. soongorica seedlings |
表1显示, 红砂2个种源的根系活力、RPP以及Pro、SS等9个指标在轻度、中度和重度胁迫与对照间差异显著(P< 0.05)。在种源间根系活力、RPP、Pro、SP、MDA和POD等指标无显著差异, 而其他几个指标在这两个种源间差异显著(P< 0.05)。表2结果表明, 红砂2个种源苗木的根系活力与RPP呈极显著负相关(
| 表1 干旱胁迫下2个种源红砂苗木根系生理指标方差分析中的Duncan多重比较检验 Table 1 Duncan multiple range test of ANOVA for two provenances of root of R. soongorica under drought stress |
| 表2 干旱胁迫下红砂2个种源苗木根系9个生理指标间的相关性分析 Table 2 Correlation analysis of nine physiology index of root in R. soongorica in two provenances under drought stress |
研究结果显示, 各项生理指标与红砂苗木根系抗旱性的关联顺序为:RPP> Pro> 根系活力> SS> POD> MDA> SP> CAT> SOD(表3)。其中, RPP、Pro和根系活力的关联度都大于0.9; SS和POD的关联度在0.8以上, 且较为接近; MDA、SP和CAT的关联度在0.7以上; 而SOD的关联度为0.5667, 远低于其他指标的关联度。这说明, RPP、Pro和根系活力3个指标对红砂根系的抗旱能力影响最大, SS和POD两个指标对抗旱能力的影响次之, MDA、SP和CAT几个指标则对抗旱能力的影响介于中间, 而SOD可能在红砂根系抗旱性中不占据重要地位。
| 表3 红砂根系各项抗旱指标的关联度及权重 Table 3 The grey correlative degree and weight of nine drought resistance indexes of root of R. soongorica |
根系是植物维持生命的主要器官, 能最早、最直接地感受到土壤水分含量的变化, 从而对干旱胁迫做出形态、生理等方面的迅速反应, 因此研究植物根系与抗旱性的关系具有重要意义[20]。试验结果显示, 随干旱胁迫的加剧, 红砂苗木根系活力逐渐减小, 且抗旱性强的武威种源比抗性弱的兰州种源下降幅度要小。这一结果与卫星等[3]在水曲柳(Fraxinus mandshurica)及单长卷等[21]在冬小麦(Triticum aestivum)上的研究结果相似。根系活力是衡量根系新陈代谢活动强弱的重要指标[2], 红砂根系活力的这种变化趋势说明, 在干旱胁迫下红砂根系可通过减弱新陈代谢活动来增强抗旱性。
细胞膜是植物细胞内外物质和信息交流的界膜, 具有选择透性, 干旱引起的膜伤害将导致膜透性增大, 同时也会使膜脂过氧化的产物丙二醛(MDA)含量增加。因此, MDA含量和细胞膜透性是反映细胞膜脂过氧化作用强弱和质膜破坏程度的重要指标[22]。以往研究表明。红砂叶片在干旱胁迫下出现质膜透性增加和膜脂过氧化的产物MDA含量升高[14]的现象。本研究结果发现, 干旱胁迫下红砂根膜透性变化特性与叶片相似, 抗性越强的种源膜透性增加越小, 说明MDA和膜透性可以作为评价红砂根系抗旱性大小的指标。
渗透调节是植物在干旱胁迫下, 降低渗透势和维持一定膨压, 抵御干旱胁迫的重要生理对策[23]。前人研究认为, 荒漠植物的渗透调节能力与抗旱性呈显著正相关, 抗旱性强的种源渗透调节能力大于抗旱性弱的种源[14]。本研究发现, 干旱胁迫显著增加了红砂幼苗根系的脯氨酸和可溶性糖含量, 且抗性强的种源增加比抗性弱的种源多。这正好证实了我们以往采用叶片研究红砂种源抗旱性的结果[14]。渗透调节物质的相对贡献率由大到小的排序为K+、可溶性糖(SS)、其他游离氨基酸、Ca2+、Mg2+、脯氨酸(Pro)[24]。本研究发现, 脯氨酸所起的渗透调节能力要大于可溶性糖的, 这一结果和前人的不一致, 说明不同渗透调节物质在不同的植物及同一植物的不同器官中的作用大小不同。
蛋白质是生命活动的体现者, 在受到干旱胁迫时, 它们会发生一定的变化, 测定其含量是了解植物抗逆性的一个重要指标[3]。本研究发现, 红砂苗木根系在遭遇水分胁迫时其根系内的可溶性蛋白含量显著升高, 分析可能是胁迫条件下抗氧化酶系合成增加所致。但是种源抗性能力不同, 其根系蛋白质含量对于重度干旱胁迫的反应不同。重度干旱胁迫使得兰州种源红砂根系蛋白质合成能力减弱(或降解过程加强), 而使武威种源的升高, 说明武威种源对重度胁迫下的水分条件有一定的抵抗力。
研究表明, 干旱胁迫下叶片中的SOD、POD和CAT对红砂的抗旱性起着关键作用[8, 14], 那么这几种酶在根系中所起的作用是否和叶片中的一样呢?本研究结果表明, 红砂2个种源苗木根系在干旱胁迫下保持较高的SOD和CAT活性以降低细胞内的活性氧对细胞膜的伤害, 但当干旱胁迫超过一定限度的时候, 根系内活性氧的积累对细胞膜的伤害增大, 保护酶活性降低。而红砂两个种源苗木根系的POD活性在干旱胁迫下, 不但没有减小反而呈持续增高趋势, 这与以往研究认为胁迫状态下红砂体内POD保护酶对活性氧的清除能力有一个阈值的结论不同[14]。这是因为, 虽然植物根系最先感知干旱胁迫, 但也是最先做出形态和生理反应和调整的器官, 根系首先优化了自己的生理特性, 再去保证地上部生理功能增强[25]。
红砂适应干旱的方式是多种多样的, 其具有综合的、几种机理共同起作用的抗旱特征[5, 12, 14]。本研究发现, 水分胁迫导致红砂根系活力下降, 膜透性和膜质过氧化产物MDA 含量增加, 渗透调节物质脯氨酸含量、可溶性糖含量和可溶性蛋白明显增加, 抗衰老酶SOD、CAT和POD活性显著升高, 说明红砂根系的抗旱性同样也是几种机理共同起作用的结果, 但每种抗旱机理所占的重要性不同。灰色关联度法分析结果表明, 红砂根系的膜透性、脯氨酸(Pro)和根系活力的关联度都大于0.9, 3个生理指标之间具有明显的相关性, 说明红砂根系通过减少膜透性的增加, 形成以Pro为主的渗透调节以及维持高水平的根系活力来提高其对干旱胁迫的适应能力。有研究认为, 红砂叶片内Pro和蔗糖含量与土壤含水量密切相关, 随着土壤含水量的下降, Pro含量明显增加, 蔗糖含量迅速积累[8, 9]。本研究结果表明, 红砂根系的SS与其抗旱性的关联度高, 说明红砂根系通过积累脯氨酸和可溶性糖而主动降低其体内渗透势, 提高其渗透调节能力, 增强自身的抗旱能力。SOD、CAT和POD 3种酶与红砂抗旱性的关联度大小为POD> CAT> SOD, 这一结果和种培芳等[14]在红砂叶片抗旱性中的结果不同(SOD> POD> CAT)。这可能与所选研究材料和抗旱指标不同有关。红砂根系MDA和SP的关联度在0.7以上, 且比较接近。说明红砂根系可通过可溶性蛋白含量的增加和膜脂过氧化程度的降低等途径来抵御和适应干旱环境。本研究通过分析发现, 红砂幼苗根系的各生理指标间存在显著相关关系, 说明虽然红砂根系各指标与抗旱性的关联性大小有明显差异, 但红砂根系并不是通过一种或少数几种机理来适应干旱, 而是发生了一系列适应性的生理变化, 来提高其根系抗旱性, 使其能更好地适应干旱逆境。
总之, 本研究结果表明, 红砂根系对干旱的适应和抵御是一个由多因素相互作用、相互控制的特征。通过分析也发现, 红砂叶片和根系对于干旱的反应既有区别又有联系, 但它们是如何协同调节红砂的抗旱性, 还需进行进一步的研究。同时, 对于红砂根系的抗旱性还应结合不同根级和不同土层进行更深层次地研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|