{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
暗紫贝母的繁殖器官在海拔梯度上的变异
[陈文年1, 2  , 肖小君
, 肖小君1, 2 , 陈发军1, 2 , 王辉1, 2 , 张志勇1, 2 , 齐泽民1, 2 , 黄作喜1, 2 ]
, 肖小君|
|


作者简介:陈文年(1968-),男,四川成都人,教授,博士。E-mail: jinzc89@yeah.net
在青藏高原东部的一个高山坡面上,沿着海拔设置高、中、低3个海拔部位,并在高、低海拔之间进行暗紫贝母植株的对换移植实验,然后测定各部位的生态因子、暗紫贝母植株花、果实和种子的特征。结果表明,1)生态因子中积雪融化时间、气温及土壤含水量在各海拔之间有明显的差异;2)花部特征中花被片的长度和宽度,雄蕊花药和花丝的长度,花柱和柱头裂片的长度都是高海拔部位最长,低海拔部位最短;3)对花生物量来说,高海拔部位比低海拔部位大39.4%;从花与地上部分二者生物量的比率来看,高海拔部位比低海拔部位大12.3%,两个指标都是高海拔部位排名第一;4)单粒果实重以低海拔部位最重,达到1282 mg,比高海拔部位大48.3%;5)单粒果实种子数以高海拔部位最少,比低海拔部位少了38.1%;6)高海拔部位的种子百粒重达到142.3 mg,而低海拔部位种子百粒重只有94.6 mg。移植实验进一步验证了上述的测定结果,这说明暗紫贝母繁殖器官确实要受海拔的影响,而且主要跟海拔梯度上的融雪时间、气温(尤其是零下低温)、紫外线强度、访花昆虫等有关。
, XIAO Xiao-JunThree sites (high, medium and low altitude) were established in an alpine mountainous region in the eastern Tibetan Plateau. The reproductive organs of Fritillaria unibracteata at each site were measured. A transplant experiment using F. unibracteata was also carried out; plants from low altitude were transferred to the high site and plants from the high site transferred to the low altitude site. Ecological factors such as snow-melt duration, air temperature, soil water content and soil pH were measured. Reproductive organs including flower, fruit and seed of F. unibracteata were also measured. Snow-melt duration, air temperature and soil water content differed among the different altitudinal sites significantly. Snowpack at the low site melted earlier than those at the other two sites; average air temperature (in March, April and May) at the low site was higher than the other two sites; however soil water content at the low site was the lowest. The highest values for petal length and width, filaments, anthers, styles and style ramifications were measured at the high site and the least at the low site. Flower biomass at the high site was 39.4% heavier than that at the low site, and the ratio of flower biomass to aboveground biomass at the high site was 12.3% higher than that at the low site. Dry weight per fruit at the low site was 48.3% heavier than that at the high site, reaching 1282 mg. Seed number per fruit at the high site was the lowest, 38.1% less than that at the low site. The 100-seed weight at the high site was 142.3 mg, 94.6 mg at the low site. The transplant experiment verified the results described above. We concluded that the reproductive organs of F. unibracteata were significantly affected by altitude factors, particularly snow-melt duration, air temperature (especially sub-zero temperatures), ultraviolet light and insect pollinators.
暗紫贝母(Fritillaria unibracteata)是百合科贝母属植物, 为多年生草本, 其野生资源主要生长于四川、西藏、云南、甘肃、青海等地的高山环境中, 海拔一般为3200~4500 m, 分布范围十分有限[1, 2, 3]。
在自然条件下, 由于暗紫贝母种子存在着形态和生理上的后熟作用[4], 其种子的萌发率十分低下[5], 因此种群密度较低, 数量十分有限。再加上人们长期大规模采挖, 其生境日益遭到破坏, 野生资源已趋于枯竭[6, 7]。因此, 如何扩大暗紫贝母种群的数量、提高其产量显得十分必要。暗紫贝母药用部分虽为其干燥鳞茎, 但其有性繁殖器官(花、果实、种子)与幼苗数量以及种群密度有着很大的关系。以前的研究很多是关于暗紫贝母的药用成分的提取和分析[8, 9], 还有些是关于器官(鳞茎)的组织培养[10, 11], 也还有部分是关于药理[12]、药效[13]和内生真菌[14]方面的。在宏观生态学领域, 有人用数学方法对暗紫贝母群落进行了分类[15], 并研究了群落类型与其品质之间的关系, 认为产于松潘等地的“ 松贝” 为最佳商品规格[16]。随着研究的不断深入, 各方面的工作也逐渐展开。有人研究了暗紫贝母生物量的分配特征对高山环境条件的生态适应[17], 也有人就暗紫贝母的鳞茎和物候在海拔梯度上的变化趋势展开了较为详细的讨论[18]。但对野生暗紫贝母繁殖器官在不同环境梯度下的比较研究还少有报道。那么, 暗紫贝母繁殖器官的特征在不同海拔又会如何变化?这是一个值得探讨的科学问题, 同时与生产实践也有着密切的联系。为此, 笔者以海拔作为环境梯度, 研究其与暗紫贝母有性繁殖器官的关系, 旨在揭示高山地区海拔对暗紫贝母繁殖特征的影响, 为暗紫贝母自然种群的扩大和产量提高奠定科学基础。
研究地位于青藏高原东部的四川省松潘县章腊乡卡卡山(32° 59' N, 103° 40' E)[19, 20], 该地区年均温为2.8℃, 1月均温-7.6℃, 7月均温9.7℃, 年降雨量634.8 mm, 其中的72%发生在6-8月的夏季, 土壤类型为黑毡土[21]。在卡卡山山脚, 海拔为3400 m左右, 山顶海拔为3980 m左右, 垂直高度差将近580 m。
研究地点设在卡卡山北坡, 取低、中、高3个海拔部位(海拔3500, 3750, 3950 m)。每个部位沿水平方向设置A、B1、B2三个30 m× 15 m(长× 宽)的调查样方(三样方间水平距离15 m左右), 在A、B1、B2样方中分别随机选取60, 30, 30株2年生(所谓“ 一匹叶” 阶段, 2~3年后开花结果)的暗紫贝母个体作为研究对象, 以A样方中植株作为对照, 在不同海拔间作对比分析。所有样方中被选择的植株都作了标记, 并于2010年7月开始在各个样方周围设置铁丝围栏以防人畜干扰。
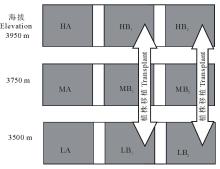
为了确定海拔对暗紫贝母的繁殖器官(花、果实和种子)是否有确切的影响, 在高海拔部位(3950 m)和低海拔部位(3500 m)的B样方之间进行植株的对换移植试验(图1)。即把高海拔B1样方中被选择的30株植株移植到低海拔部位的B1样方中, 同时把低海拔部位B1样方中被选择的30株植株移植到高海拔部位的B1样方中; 高、低海拔部位B2样方中的30株植株也采用相同的移植方法。移植过程中采用连土移植, 即每株暗紫贝母连同其植株下面的土块一起移植, 土块的长、宽、深大小尺寸为0.20 m× 0.20 m× 0.25 m。整个移植试验于2010年9月上旬进行, 所有的移植植株都作了标记, 以A样方中的植株作为对照。移植实验完成之后, 在高、低海拔部位就产生出6种植株类型, 即高海拔部位A样方的对照植株(HA)、高海拔部位B1样方中的来自低海拔的移植植株(HB1)、高海拔部位B2样方中的来自低海拔的移植植株(HB2); 低海拔部位A样方的对照植株(LA)、低海拔部位B1样方中的来自高海拔的移植植株(LB1)、低海拔部位B2样方中的来自高海拔的移植植株(LB2)。另外, 中间海拔部位(海拔3750 m)还有3个样方, 即中间部位A样方中的植株(MA)和中间部位B1和B2样方植株(MB1、MB2), 如图1所示。
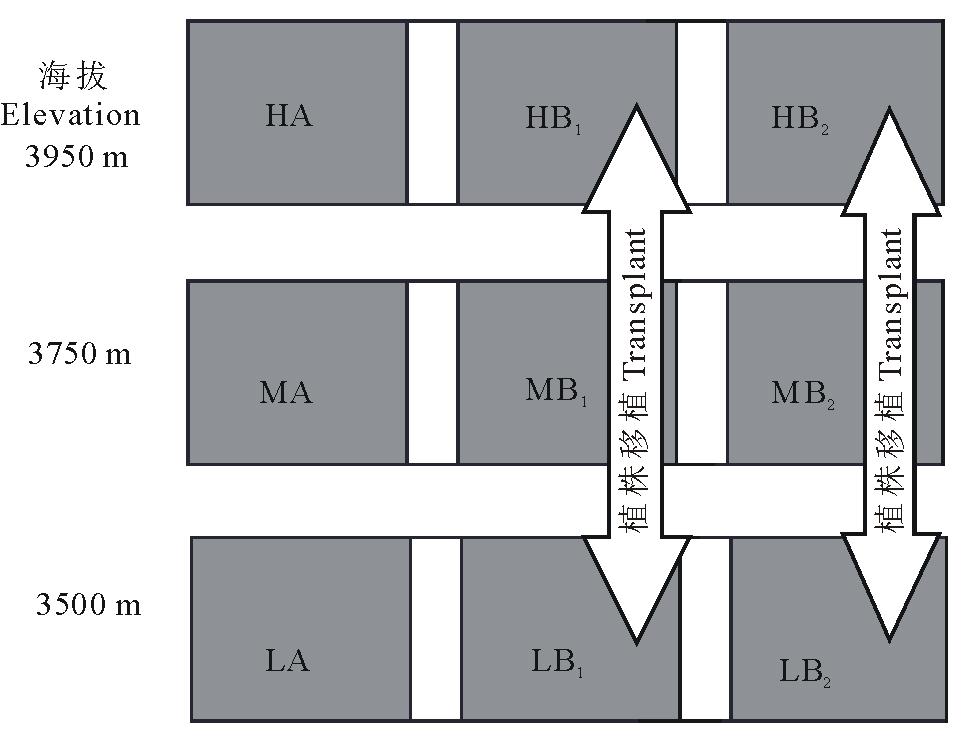 | 图1 样方设计示意图Fig.1 A sketch map showing plots design |
1.3.1 各部位生态因子的测量
在2012和2013年的3-5月, 测量各样方中各标记植株从积雪中融化出来的时间(以1月1日开始的序日表示); 同时在高、中、低各海拔部位, 分别放置5个自动温度记录仪(型号:ZDR-01)以测定3-5月的气温变化, 记录仪离地高度为50 cm, 设定为每h自动记录1次; 在5月中旬(此时积雪已融化完, 冻土已经解冻)到8月底, 用便携式水分测量仪(型号:Mpkit-B)和pH计(型号:PHT-810)测量每个样方中的土壤含水量及土壤酸碱度, 每个样方取中心和四周测量点5个, 每周1次。
1.3.2 花部特征的测定
2013年7月上旬, 正值暗紫贝母的开花期, 在A样方标记的60株中随机选30株用于测定花部特征, 剩下的30株留着在下一年的8月下旬测定果实和种子特征, B1样方中标记的30株也用于测定花部特征。用镊子和解剖针轻轻地拔开花朵, 用数显游标卡尺(型号:SM208-1)测量各部位的A样方中选出的30株和B1样方中标记的30株花器官的组成成分长度, 包括各花瓣片的长与宽、雄蕊花丝和花药的长度、雌蕊花柱和柱头裂片的长度。
1.3.3 植株地上部分及花生物量的测定
2013年7月, 在测定完上述A和B1样方中各30株植株的花部特征之后, 紧接着对这些测定了花部特征的植株进行地上部分生物量及花部生物量的测定, 所用方法为收割法。将这些植株地上部分平地割下放入信封袋中, 信封袋上作好编号, 将装有植株的信封放入恒温箱中65℃烘干至恒重[22], 用万分之一电子天平(型号:FA1004)将各植株的地上部分进行称量、记数。再摘下花器官, 对花器官单独进行称量和记数, 并计算每一植株花的生物量与其地上部分生物量的百分比值作为该植株繁殖分配的一个指标(花/地上部分)。
1.3.4 植株地上部分及果实和种子生物量的测定
2014年8月下旬, 在果实和种子的成熟期, 用收割法将各海拔部位A样方标记植株(共60株)中剩下的30株、B2样方中标记的30株平地割起, 分别装在不同的信封袋中, 作好编号。将装有植株地上部分(含果实)的信封袋放入恒温箱中65℃烘干至恒重[22]。用万分之一电子天平(型号:FA1004)将各植株干重进行称量、记数。然后从植株上轻轻剪下果实, 称量其干重, 并计算果实生物量与植株地上部分生物量的百分比值。再从果实中取出种子, 记录种子的个数, 同时称量每个果实中的种子干重。
用SPSS 17.0对不同海拔部位间生态因子、花部组成特征、果实和种子特征进行方差分析(one-way ANOVA)和多重比较, 所得数据以“ 平均值± 标准误” 表述。
如表1所示, 3个海拔的融雪时间有明显的差别, 低海拔部位的积雪融化最早, 在3月下旬和4月初即可全部融化, 而高海拔部位则要在4月下旬才可全部融化, 高低部位之间相差可达25 d左右(表1)。土壤含水量在4个部位之间差异显著, 高海拔部位比低海拔部位高16.7%。3个部位的pH相差不大, 都呈弱酸性(表1)。 另外, 从3-5月的气温来看(表1), 每个月都是以低海拔部位平均气温最高, 以高海拔部位的平均气温最低, 高、低部位之间的平均气温差异分别达到了2.5(3月), 2.8(4月), 6.6℃(5月)。
| 表1 3个海拔部位环境因子的比较 Table 1 Comparison of environmental factors among three portions |
2.2.1 花被片长与宽
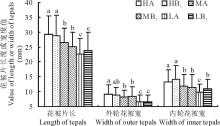
由于内轮与外轮花被片的长度几乎是等长的, 故统计花被片长度时不作外轮与内轮的区分; 但内轮与外轮花被片的宽度却有相当大的差异, 因此, 在统计时要作区分。从图2可以看出, 花被片的长度和宽度都是以高海拔部位(HA, HB1)最大, 低海拔部位(LA, LB1)最小, 而中间部位(MA, MB1)居于二者之间。从长度来看, 高海拔部位(HA样方)比低海拔部位(LA样方)长29.1%; 从外轮花被宽来看, 高海拔部位(HA样方)比低海拔部位(LA样方)宽40%; 从内轮花被宽来看, 高海拔部位(HA样方)比低海拔部位(LA样方)宽36.1%。除了低海拔部位的内轮花被片外, 同一海拔部位的对照样方(A)和移植样方(B1)之间差异几乎都不显著。高海拔植株被移植到低海拔部位后其花被片的特征表现出低海拔部位对照样方中植株的特点; 反之, 低海拔部位的植株被移植到高海拔部位后同样也表现出高海拔部位对照样方中植株的特点(图2)。
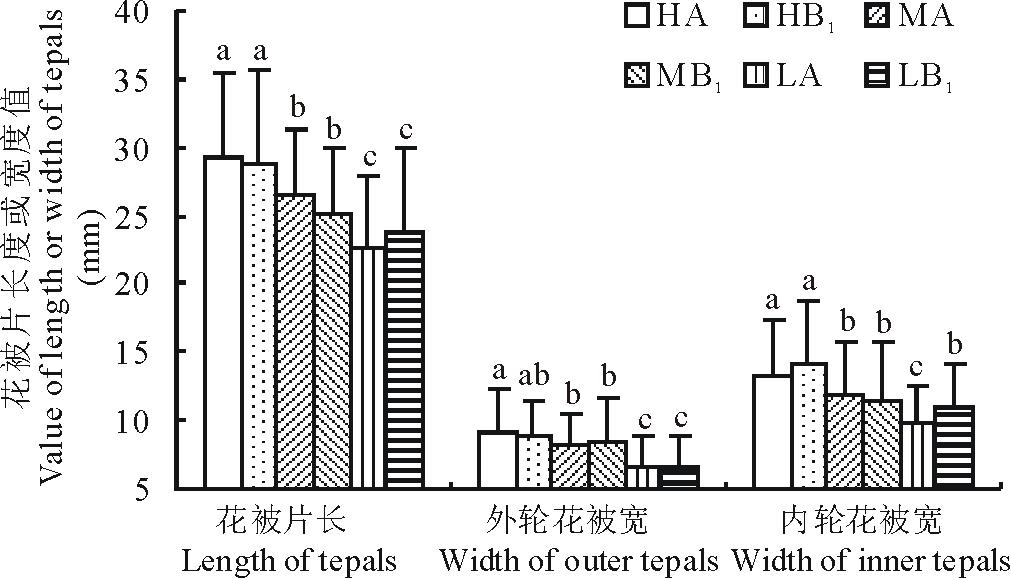 | 图2 不同处理对花被片长或宽的影响Fig.2 Comparison of length or width of tepals among all plots |
2.2.2 雄蕊花丝和花药的长度
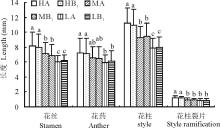
从图3可以看出, 暗紫贝母雄蕊的花药和花丝以高海拔部位最长, 低海拔部位最短, 中间部位居于二者之间。且花丝的长度在高、中、低海拔部位之间的差异显著, 但花药只在高、低海拔部位之间才达到显著。从花丝来看, 高海拔部位(HA)比低海拔部位(LA)长约36.6%; 从花药来看, 高海拔部位(HA)比低海拔部位(LA)长约22.0%。无论在哪一个海拔上, 对照样方与移植样方之间的差异都未达显著水平。
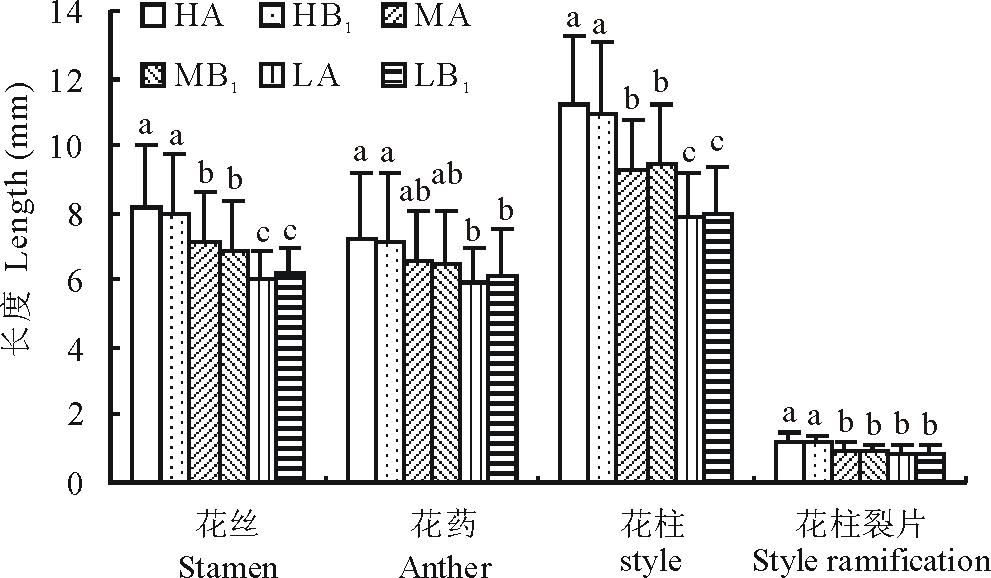 | 图3 不同处理对雄蕊和雌蕊性状的影响不同字母表示差异显著(P< 0.05), 下同。Fig.3 Effects of differrnt treatments on characters of stamen and pistil Different letters mean the significant differences at P< 0.05, the same below. |
2.2.3 雌蕊花柱和柱头裂片的长度
如图3所示, 花柱长度和柱头裂片长度都是高海拔部位最长, 低海拔部位最短, 而中间部位居中。就花柱来说, 高海拔部位比低海拔部位长41.8%; 对柱头裂片而言, 高海拔部位(HA)比低海拔部位(LA)长42.3%。同一部位的对照植株(A)和移植植株(B1)之间表现出了几乎相同的特征(图3)。花柱长度在各部位间的差异显著; 而柱头裂片则在中、低海拔部位间未达显著水平。
从表2可以看出, 各海拔部位各样方植株的花生物量、地上部分生物量及二者的比率存在明显的差异。对花生物量来说, 高海拔部位(HA样方)比低海拔部位(LA样方)大39.4%; 而对地上部分总生物量而言, 高海拔部位(HA样方)却比低海拔部位(LA样方)小17.2%。从花与地上部分二者生物量的比率来看, 高海拔部位(HA样方)比低海拔部位(LA样方)大12.3%。无论是花生物量、地上部分生物量还是二者的比率在各部位之间都达到差异显著的程度。但同一海拔的对照样方与移植样方之间差异不显著(表2)。
| 表2 花和地上部分生物量及其比例 Table 2 Flower biomass, aboveground biomass and their ratio |
2.4.1 单粒果实重及果实与地上部分之比
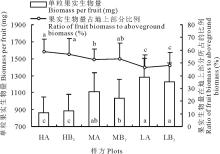
如图4所示, 单粒果实重以低海拔部位的样方(LA)最重, 达到1282 mg, 比高海拔部位样方(HA)的864 mg高48.3%, 中海拔部位(MA)介于高、低海拔之间, 各海拔部位之间的差异达到显著水平。从果实在地上生物量中所占的比例来看, 各海拔部位之间的差异也基本达到显著水平, 但表现出的趋势却与单粒果实生物量相反, 即从低海拔到高海拔逐渐增加(图4)。低海拔部位(LA)所占的比例只有46.6%, 比高海拔部位(HA)的58.7%少了12.1%。另外, 不管是单粒果实重还是果实在地上部分所占的比率, 同一部位的对照样方与移植样方之间差异并不显著, 只有不同部位之间的差异才明显。
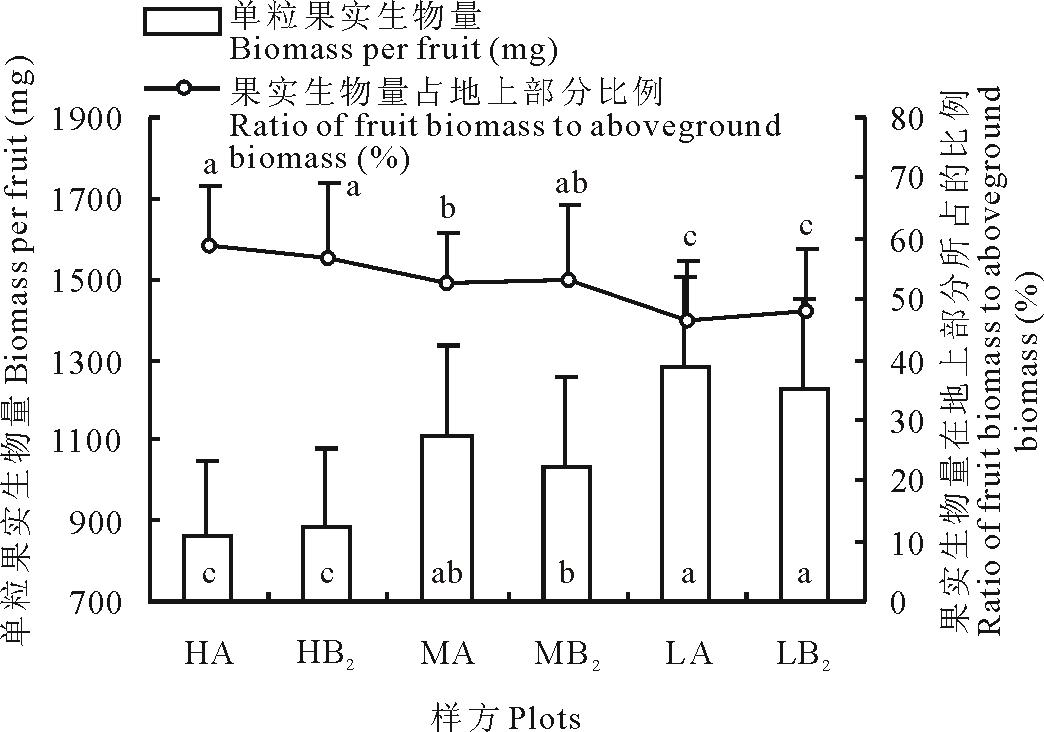 | 图4 果实生物量及其在地上部分所占的百分比Fig.4 Biomass per fruit and the ratio of its biomass to aboveground biomass |
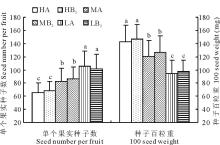
2.4.2 单粒果实种子数及种子百粒重
如图5所示, 单粒果实平均种子数以高海拔部位(HA)最少, 仅有65.4粒, 比低海拔部位(LA)的105.7粒少40.3粒, 少了38.1%。但高海拔部位(HA)的种子要明显地大一些, 其百粒重达到142.3 mg; 而低海拔部位(LA)种子百粒重只有94.6 mg, 比高海拔部位小了33.5%。不论是单粒果实种子数还是种子百粒重, 在各部位之间的差异都显著, 但同一部位之间的移植样方和对照样方之间的差异不显著(图5)。
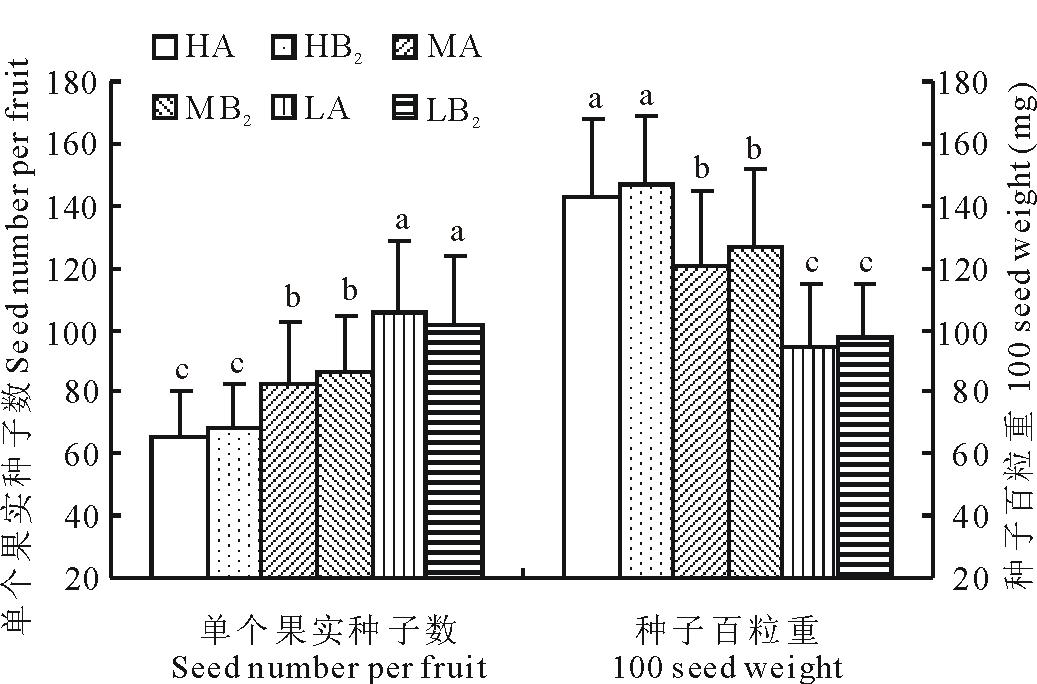 | 图5 单个果实种子数及种子百粒重Fig.5 Seed number per fruit and 100 seed weight |
在高山生态系统中, 风媒传粉的植物类型所占比例很低, 仅4.32%, 而虫媒植物所占比例极高, 可达95.62%[23]。暗紫贝母每株只开1朵单花, 种群密度比较低, 再加上高山区域降雨较为频繁, 因此靠风媒传粉很难完成其生活史; 另一方面, 其花粉粒较大, 表面粗糙, 完全符合虫媒传粉的特征。据笔者在研究区域内的长时间观察, 其传粉昆虫主要为熊蜂和苍蝇, 因此属于典型的虫媒传粉植物。
在本研究中, 随着海拔的升高, 暗紫贝母花被片的长与宽都有所增加, 其雄蕊、花柱和花柱裂片都变长(图2, 图3)。花被片的增大, 会使得整个花在外形上变大, 加上花被片的颜色为紫红色, 这在高山生境中就显得更加醒目, 有利于吸引更多的昆虫前来传粉。同时其花药的变长, 表明其可以产生更多的花粉; 花丝的变长使得花药更好地展布于空中, 有利于接触传粉昆虫; 而花柱和花柱裂片的变长则有利于接受昆虫携带的来自其他植株的花粉。Bingham和Orthner[24]的研究认为, 在高山生态系统中, 随着海拔升高, 访花昆虫的种类和活动能力均逐渐降低, 暗紫贝母花在形态上的这些变化正好使得在高海拔处由于传粉昆虫种类和活动减少而造成的传粉限制得到补偿; 同时高海拔处紫外线更强烈, 相对较大的花瓣, 还可以减少强烈的紫外线伤害, 从而对花的内部结构起到很好的保护作用。这些显然是暗紫贝母植株对高海拔环境因子适应的结果。
随着海拔的升高, 暗紫贝母地上部分生物量逐渐降低(表2)。这表明, 随着海拔的升高暗紫贝母的植株有变小、变矮的趋势。其原因跟海拔升高后, 融雪时间的推迟(表1)及无雪期的缩短有关。因为在积雪晚融的区域, 无雪期长度的缩短限制了植物积累更多的营养物质, 而这些营养物质是来年生长所必需的[25]。其他学者的研究也认为, 在积雪环境中, 无雪期长度是决定植物生长的主要因素[26], 生活在温带地区的高山植物, 更多的是受无雪期长度的影响[27]。显然本研究中的地上部分生物量随海拔升高而降低与上述这些学者研究的结果是一致的。另外, 在高海拔部位, 这种植株高度的降低对吸引昆虫访花传粉也有着积极的作用, 因为有研究认为较矮的株高所形成的温暖花部环境可以吸引更多的传粉昆虫访花[28], 这对高海拔部位植株完成生活史显然有重要的意义。
与地上生物量不同, 花生物量及其在地上部分中所占比率(繁殖分配)随海拔的升高而逐渐升高(表2)。Kawano和Masuda[29]对其他植物居群的研究发现, 随着海拔升高, 繁殖分配增加。Fabbro和Kö rner[28]比较了生长在低海拔和高海拔地区的多种植物后, 发现生长在高海拔地区植物的繁殖分配远远高于低海拔地区的植物, 说明了有性繁殖对高海拔植物而言具有更为重要的意义。显然本研究中暗紫贝母花生物量的表现与这些学者的结果也具有一致性, 符合Weiner等[30]提出的植物繁殖分配与个体大小间的线性关系模型, 即繁殖分配与个体大小是一种简单的线性负相关, 因为在本研究中从低海拔到高海拔, 暗紫贝母的植株大小正好是逐渐减小的。出现这种结果的原因可能与高山环境中积雪过早融化而导致的较低零下低温有关。因为在研究地所在的高山环境中, 早春(3-5月)夜间气温可降至-6℃, 而在较高海拔处融雪时间推迟, 由于有积雪层的覆盖, 地面温度不至于降的太低[19, 20], 而高山植物的繁殖器官对零下低温十分敏感[31], 植物繁殖器官从雪被中过早地暴露会增加霜冻的风险。因此, 在低海拔部位积雪早融的情况下, 生长季早期积雪层的丧失更容易引起雪生植物繁殖器官的冻害, 从而影响花生物量的积累及其在地上生物量中所占的比例。
在高海拔部位, 由于花的生物量相对较大, 在花期维持花的生长和发育就耗去了植物体相对更高的营养资源, 再加上高海拔部位植株相对较为矮小, 因此在完成开花、传粉及受精等过程后, 果实的生长相对营养受限, 于是表现出从低海拔到高海拔果实逐渐减小的趋势。另外, 在高海拔处, 环境条件趋于更加恶劣, 降雨相对更为频繁, 这会增强对花药和柱头上花粉的冲刷作用, 无疑增加了花粉传递过程中的风险; 同时, 高海拔处相对恶劣的自然条件也降低了昆虫的多样性和活动频率[24], 因此暗紫贝母种子的产生难免不受到花粉限制[32, 33], 胚珠的受精率自然下降, 果实中的种子数就趋于减少。但另一方面, 高海拔部位的暗紫贝母还是将有限的营养资源用在了种子的生长上, 由于种子数量减少, 单粒种子得到的营养资源反而有所增加, 于是种子相对变大, 这从种子的百粒重上可以反映出来。有研究表明, 高山地区不同部位融雪时间的差异可以影响雪生植物种子的大小[34], 而种子的大小又会进一步影响幼苗的生长发育及形态大小[35], 本研究中不同海拔部位融雪时间的不同导致的种子大小差异, 与上述的研究结果一致。而较大的种子可以提升幼苗的适合度或存活力[36], 这对于暗紫贝母幼苗在环境条件相对更为恶劣的高海拔部位顺利成长并完成其生活史有着十分重要的意义。
从移植实验结果来看, 不管高海拔还是低海拔植株在被移植后都表现出其移栽后所在部位的对照植株的特点。这表明, 暗紫贝母繁殖器官的特征在海拔梯度上的变化可能更多的是受与海拔紧密相关的一些环境因子的影响, 如融雪时间、气温(尤其是零下低温)、紫外强度、访花昆虫等。在不同的海拔部位, 暗紫贝母植株形成了与这里的环境因子相适应的繁殖器官特征, 在环境发生变化时, 也能在相对较短的时间内作出调整, 从而保证了该物种能够在不同的海拔梯度上都能完成自己的生活史, 使自己能够在各种恶劣的条件下存活下来。相对而言, 土壤水分及pH等土壤因子对其繁殖器官的影响可能较小。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [21] |
|
| [22] |
|
| [36] |
|