
{kind=link}
苜蓿锈病病菌侵染条件的研究
[李跃 , 袁庆华
, 袁庆华* ]
, 袁庆华]
|
|


作者简介:李跃(1986-),男,河北承德人,在读硕士。E-mail:liyue_s@163.com
通过恒温箱和温室试验,探索了不同的保湿时间和温度对苜蓿锈菌孢子堆密度的影响以及不同培养温度对病害潜育期的影响。19℃下保湿(相对空气湿度100%)一定时间,再在25℃下生长15 d,可见保湿时间在12~48 h处理中植株叶片上孢子堆密度高于4~8 h。不同保湿温度下,保湿(相对空气湿度100%)24 h,再在25℃下生长15 d,试验结果表明随着保湿温度的升高,孢子堆密度先逐渐增加,在19℃左右达到最大,之后逐渐降低。19℃下,100%相对湿度下保湿24 h,然后分别置于15,20,25,30和35℃下生长,只有35℃下不发病,其余温度下潜育期分别为11,10,7和7 d;培养温度在25和30℃下的孢子堆密度显著高于15和20℃;培养温度对孢子堆显现时间的长短影响不大。
Uromyces striatus is the causal organism of alfalfa rust. The effects of humidity, temperature and time on disease severity and the effects of incubation temperature on the latent period of alfalfa rust were investigated using growth chamber and greenhouse experiments, respectively. Alfalfa plants were inoculated with U. striatus and incubated at 19℃ with 100% relative humidity (RH) for different periods before being transferred to 25℃ conditions for 15 days. Subsequently, spore densities on the leaves of inoculated plants were assessed. The results showed that spore density of treatments incubated at 19℃ for 12 h to 48 h was significantly higher than those incubated for 4 h to 8 h. Plants were also inoculated and incubated at 19℃ with 100% RH for 24 h and then transferred to different temperature environments; 15, 20, 25, 30 and 35℃. Plants transferred to 35℃ conditions did not display any alfalfa rust symptoms. The disease latent period on inoculated plants grown at 15, 20, 25 and 30℃ was 11, 10, 7 and 7 days, respectively. The spore density on leaves in the 25 and 30℃ treatments was significantly higher than the 15 and 20℃ treatments. There was no relationship between incubator temperature and disease latent period.
紫花苜蓿(Medicago sativa)是世界上栽培和种植非常广泛的豆科牧草, 是家畜的主要蛋白饲料, 也是我国近年来大面积推广和种植的优良牧草[1, 2, 3, 4]。随着苜蓿种植面积的不断扩大, 病害问题越来越严重, 尤其是苜蓿锈病, 该病由苜蓿条纹单孢锈菌(Uromyces striatus)引起, 是苜蓿上的重要病害之一, 在我国各地均有发生, 主要发生在苜蓿种植较多的地区, 如:西北、华北和东北等地, 常造成不同程度的危害, 严重时苜蓿减产10%~30%, 种子减产50%[5], 粗蛋白含量下降18%, 粗纤维含量增加15%[6]。因此, 近年来许多研究者对苜蓿锈病及其病原菌进行了研究。国外对苜蓿锈病及其病原菌的研究较早, 而且较为深入。Webb和Nutter[7]探索了不同温湿度和保湿时间对苜蓿条纹单孢锈菌侵染效率以及潜育期等的影响。Kemen等[8]在被苜蓿条纹单孢锈菌侵染的蒺藜苜蓿(Medicago truncatula)细胞中发现了锈菌转移蛋白(Us-RTP1), 证明苜蓿条纹单孢锈菌吸器不仅仅是从寄主细胞吸取营养, 二者之间还存在其他联系。Castillejo等[9]对蒺藜苜蓿-苜蓿条纹单胞锈菌互作系统进行了蛋白质组学研究, 并鉴定出了27个与光合作用、能量代谢通路和胁迫反应相关的差异表达蛋白。在转录水平上, 不同抗锈机制的蒺藜苜蓿在受到锈菌侵染时, 与细胞防御相关的转录因子区别表达, 这些转录因子有组成表达的也有诱导表达的[10]。Abdel-Monaim等[11]发现在紫花苜蓿叶面施以过氧化氢、甲基茉莉酮酸酯或β 氨基丁酸可以增强紫花苜蓿抗锈性。Kemen等[12]对5份抗苜蓿锈病的蒺藜苜蓿种质进行了侵染初期的扫描电镜组织观察和荧光显微观察, 发现了两种抗锈机制:一种发生在吸器形成以后, 表现为叶片细胞不同程度的过敏性坏死; 另一种发生在吸器形成以前, 没有过敏反应发生。Van Der Merwe和Walker[13]及Barilli等[14]分别得到了苜蓿条纹单胞锈菌的微管蛋白基因编码序列和核糖体RNA基因及内部转录间隔区序列。国内主要对苜蓿锈病的发生及危害进行了田间调查, 初步开展了田间防治试验, 抗病种质材料的筛选等研究[15, 16, 17, 18, 19, 20]。例如, 南志标等[15]对104份紫花苜蓿材料和4份黄花苜蓿(Medicago falcata)材料进行了田间条件下的抗锈性筛选, 结果显示阳高、富平和咸阳等品种的抗锈性较强。周淑清等[16]对112份苜蓿种质材料进行了室内接种抗锈性鉴定和田间自然条件下的抗锈性鉴定, 筛选出了阳高苜蓿和草原2号等抗锈性较好的品种, 并认为鄂旗苜蓿可作为抗病育种的抗源材料。而关于苜蓿锈病病菌侵染条件的研究, 国内还未见报道。由于国内的气候条件、病原的习性存在一定差异, 其侵染条件及潜育期可能存在一定差异, 故本研究的目的是摸清国内苜蓿锈病菌的侵染条件和潜育期, 以期对国内苜蓿锈病的防治和预测提供参考。
试验所用苜蓿品种为新疆大叶苜蓿。供试菌种为苜蓿条纹单孢锈菌, 于2012年10月采自中国农业科学院北京畜牧兽医研究所廊坊苜蓿试验地, 田间发病率为50%。大量摘取苜蓿锈病病叶, 荫干后装入自封袋中, 于2~4℃冰箱内保存。2个月后在该地温室内扩繁, 扩繁时在烧杯中用少量石油醚将夏孢子从叶片上洗下, 然后取出叶片, 将烧杯中的夏孢子石油醚混合物转移到离心管中, 在1000 r/min转速下离心30 s, 弃掉上清液, 待剩余的石油醚挥发干净后, 将夏孢子粉加入到0.1%的吐温20溶液中, 搅拌, 形成孢子悬浮液, 并稀释到浓度为105个孢子/mL。将配制好的孢子悬浮液喷洒在生长60 d以上的苜蓿幼苗上, 在19℃下保持100%相对空气湿度24 h; 在25℃下培养15 d后将发病叶片上的夏孢子粉敲落到锡箔纸上并收集到1.5 mL离心管中, 贮存于2~4℃冰箱内备用, 45 d后用于接种。
1.2.1 幼苗的培养 首先将苜蓿种子放在培养皿内的滤纸上, 加适量的水以保持种子吸水膨胀, 然后放入20℃的生长箱内, 发芽2 d, 待幼根长到1 cm长时, 移植到装有消毒土壤的塑料花盆(直径10 cm, 深15 cm)中, 每盆3株苜蓿, 放入20~30℃温室内培养, 待幼苗生长60 d后进行病原菌接种。
1.2.2 不同保湿时间处理 将已喷洒夏孢子悬浮液的苜蓿幼苗分别在19℃下保湿4, 8, 12, 24, 36和48 h。然后, 在25℃温室中培养15 d。
1.2.3 不同保湿温度处理 将已喷洒夏孢子悬浮液的苜蓿幼苗分别放置在13, 16, 19, 22, 25, 28和31℃下保湿24 h。保湿结束后, 在25℃温室中培养15 d。
1.2.4 不同培养温度处理 将苜蓿幼苗接种(接种温度为19℃, 保湿时间为24 h)后分别置于15, 20, 25, 30和35℃的恒温培养箱中继续培养。
1.3.1 统计及接种方法 以上3组试验中的每个处理3株幼苗, 每处理接种量均为2 mg, 并均设置4次重复。接种前标记每株最顶端完全展开的3枚叶片。接种时, 将夏孢子粉加入到0.1%的吐温溶液[21]中, 充分搅拌, 制成夏孢子悬浮液; 用喉头喷雾器将孢子悬浮液均匀喷洒在苜蓿植株上, 然后将接种的植株转移到相对空气湿度为100%的保湿容器中, 将容器放在黑暗的生长箱中, 保湿时间、保湿温度和培养温度根据各处理来确定。记录首个孢子堆出现时间以及孢子堆不再增加时的时间, 以确定孢子堆显现时间。孢子堆显现时间是指第一个孢子堆出现时开始至最后一个孢子堆出现时为止所经历的时间。接种完毕继续培养15 d后统计已标记叶片上的孢子堆数和叶面积, 计算孢子堆密度。
1.3.2 孢子堆密度统计方法 统计标记叶片上孢子堆数目, 并测得相应叶片的叶面积。叶面积测定方法为:将标记的发病叶片制成蜡叶标本, 放在坐标纸上, 编号, 用佳能单反相机拍照后通过Photoshop cs 软件按照崔华威等[22]的方法测量单枚叶片的像素数以及坐标纸上1 cm2小格的像素数。用叶片的像素数比上1 cm2小格的像素数即为该叶片的叶面积。孢子堆密度即为每片叶上的孢子堆数与该叶片的叶面积之比。
用Excel 2003软件进行数据整理和作图, SAS 9.2软件对试验中各个处理的平均值进行相关的回归分析和方差分析(P< 0.05)。
从表1可以看出, 保湿时间在12, 24和48 h处理中植株叶片上孢子堆密度显著高于4和8 h(P< 0.05)。保湿时间为36 h处理时植株叶片孢子堆密度也高于8 h, 但差异不显著。
| 表1 保湿时间为4, 8, 12, 24, 36和48 h时孢子堆密度的LSD方差分析 Table 1 LSD variance analysis of density of pustule when leaf wetness duration was 4, 8, 12, 24, 36 and 48 hours respectively |
不同保湿温度与孢子堆密度之间的二项式回归分析结果表明(图1), 在13℃至31℃范围内, 随着温度的升高, 孢子堆密度逐渐增加, 在19℃时达到最大值, 之后逐渐下降, 培养温度升高至31℃时孢子堆密度为0。
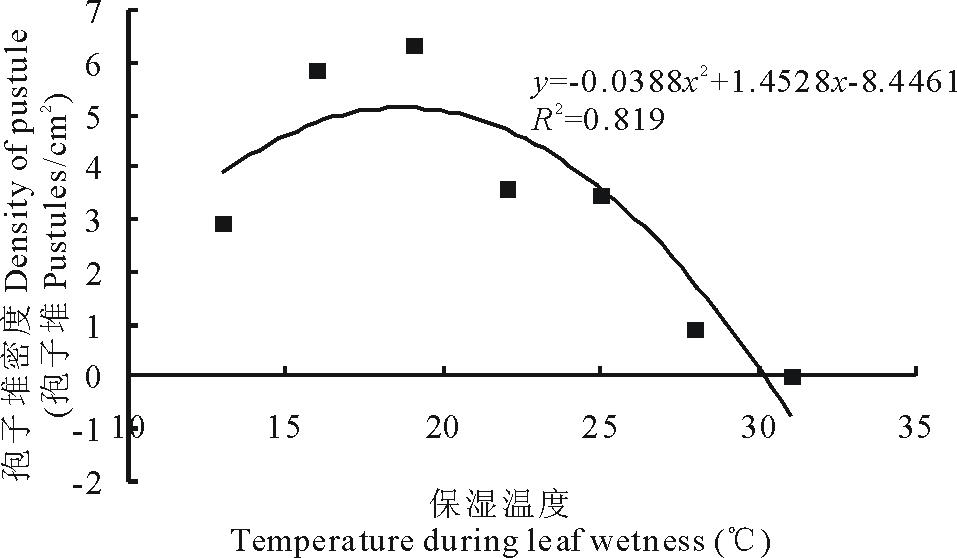 | 图1 保湿温度对孢子堆密度的线性回归Fig.1 Linear regression of temperature during the 24 hours period of leaf wetness following inoculation on density of pustule of U. striatus on the alfalfa cultivar Xinjiangdaye |
对不同培养温度处理下孢子堆密度的方差分析结果(表2)显示:15和20℃两个温度处理下的孢子堆密度与25和30℃两个温度处理下的孢子堆密度差异显著(P< 0.05)。培养温度为35℃时孢子堆密度为0, 即该温度下不能发病。
| 表2 接种后温度对苜蓿锈菌孢子堆密度的影响 Table 2 Effect of temperature after inoculation on density of pustule |
不同培养温度下的苜蓿锈病潜育期和锈菌孢子堆显现时间的长短如表3所示。保湿后培养温度越低锈病的潜育期越长, 相反, 当温度越高时潜育期越短。15和20℃时, 潜育期分别为11和10 d, 当培养温度达25和30℃时, 锈病的潜育期只有7 d, 比15和20℃时分别少了4和3 d。不同培养温度处理间对锈菌孢子堆显现时间长短的影响不大, 4个温度处理, 除20℃外, 其他3个温度处理孢子堆显现时间基本都是5 d, 这说明不同温度处理, 对锈病孢子堆显现时间没有影响。
| 表3 接种后温度对潜育期以及孢子堆显现时间的影响 Table 3 Effect of temperature after inoculation on latent period and time of pustule appearance |
对不同保湿时间对孢子堆密度的影响进行了试验研究, 结果显示, 苜蓿条纹单孢锈菌接种的最佳保湿时间为12~24 h, 低于12 h会降低孢子堆密度, 高于24 h孢子堆密度也不会增加, 这一结论与Webb和Nutter[7]在Ranger品种上进行的抗锈病试验的结果相一致, 即保湿时间为24 h时侵染效率(每叶片孢子堆数)最高。这也符合锈菌夏孢子在自然感病条件下的特性。不同保湿温度对苜蓿锈菌孢子堆密度有较大的影响, 本研究显示在13~31℃范围内, 接种温度过高或过低都不利于夏孢子的侵染; 温度低于19℃时, 孢子堆密度随着温度的升高而增加; 19℃时孢子堆密度最高; 超过19℃时, 试验结果与Webb和Nutter[7]的研究结果基本一致, 即在17.5℃到28℃之间, 接种温度与侵染效率(每叶片孢子堆数)呈负相关。许多研究表明, 苜蓿锈病喜欢凉爽湿润的气候, 在我国东北、华北和西北地区苜蓿锈病往往是秋季发生。本论文通过研究苜蓿锈菌夏孢子侵染对温度及湿度的要求, 可以更好地了解和摸清苜蓿锈病的发生情况, 为病的预测预报和防治提供依据。
不同培养温度对苜蓿锈菌孢子堆密度有显著的影响, 这一研究结果与Al-Hamdany[23]的研究结果相一致, 而与Webb和Nutter[7]的研究结果不同。Webb和Nutter[7]的研究结果表明不同培养温度对苜蓿锈菌孢子堆密度没有显著的影响。这可能是因为各研究者使用的品种不同, 因各品种之间抗病性存在一定的差异, 从而导致研究结果的不同。当培养温度达到35℃时不发病, 说明过高的温度不利于苜蓿锈病的发生。不同培养温度对苜蓿锈病害潜育期的长短有一定的影响, 研究结果表明, 培养温度为25℃时苜蓿锈病的潜育期最短, 当温度升高时病害的潜育期不会缩短, 降低温度则会延长潜育期, 这一结论支持Stuteville[24]的苜蓿锈病鉴定标准, 即接种苜蓿锈菌后苜蓿植株的培养温度设置为25℃为宜。有关不同培养温度对苜蓿锈病孢子堆显现时间的影响不明显, 4个温度处理, 孢子堆显现的时间均需要5~6 d, 这一结果与Webb和Nutter[7]的试验结果有差距, Webb和Nutter[7]认为孢子堆的显现时间受温度的影响较大, 温度在15~25℃时, 孢子堆的显出需要较长的时间, 只有当温度较高时(30℃), 孢子堆出现的时间才与本实验结果相一致, 这可能是因为苜蓿锈病具有不同的生理小种, 生理小种不同致病力也就不同, 另外, 苜蓿品种不同抗病性也就不同, 所以才会出现以上结果。
综上所述, 苜蓿条纹单孢锈菌的成功侵染需要高湿和19℃左右的温度, 而我国北方秋季夜间的温湿度条件刚好可以满足苜蓿锈病病菌的侵染条件; 一旦侵染成功, 与寄主建立起寄生关系后, 25~30℃左右的温度即可使苜蓿锈病的潜育期达到最短, 在我国北方地区的秋季气温也可以达到这一范围。因此, 苜蓿条纹单孢锈菌的这一习性是使秋季成为苜蓿锈病的高发季节的主要原因。在气候条件特别适合的情况下, 病害可在同一个生长季中进行多次侵染, 严重时可造成病害的大发生。所以在苜蓿锈病防治之前, 应摸清该病的发生规律, 哪些是影响病害发生的主要因素, 可以更好地制定防治措施, 有效地控制病害。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|