
{kind=link}
{kind=link}
{kind=link}
水热因子对克氏针茅草原土壤呼吸及其土壤温度敏感性的影响
[谷蕊1, 2  , 潮洛濛
, 潮洛濛1, * , 张立欣1, 2 , 苏力德1, 2 , 万志强1, 2 , 闫玉龙1, 2 , 陈雅丽1, 2 , 高清竹2, * ]
, 潮洛濛, 张立欣]
|
|


作者简介:谷蕊(1990-),女,蒙古族,内蒙古赤峰人,在读硕士。E-mail:guruiwudan2008@163.com
作为内蒙古重要的草地资源,克氏针茅草原在我国畜牧业生产中占有重要的地位。土壤呼吸是大气与陆地生态系统碳循环的主要通量,是土壤碳输出的主要途径。试验区设置在内蒙古锡林浩特市东部毛登牧场内以克氏针茅建群的典型草原;在2011-2013年采用开顶式生长室(OTC)对试验样地进行模拟增温增雨处理,在生长季(5-9月份)使用开路式土壤碳通量测量系统(LI-8100, LI-COR,NE) 对试验样地土壤呼吸强度进行测定;同时监测试验样地气温、土壤温湿度等环境因子,采集土壤样品并室内分析土壤养分特征和微生物生物量,并对土壤呼吸与环境因子间进行相关性分析。克氏针茅草原土壤呼吸受空气温度和土壤含水量影响显著( P<0.05),分别呈现出二项函数和线性相关关系;不同处理下土壤呼吸在相同时间存在显著差异( P<0.05),2012年增温增雨与增温处理下土壤呼吸速率分别为2.60和1.96 μmol/(m2·s), 2013年增温增雨与增温处理下土壤呼吸速率分别为3.38和2.09 μmol/(m2·s);增温增雨处理下,克氏针茅草原空气温度的升高和土壤含水量的降低均对土壤呼吸产生了抑制。土壤呼吸温度敏感度(Q10)在2011-2012年中增温增雨处理明显高于对照处理和控温处理;2011-2013年控温处理都低于对照处理。本研究表明,土壤呼吸主要受水热条件影响,并且空气温度升高和土壤含水量的降低均对土壤呼吸有抑制作用,增温增雨处理下土壤呼吸速率要高于增温处理;土壤呼吸温度敏感系数与土壤温度、土壤含水量、土壤有机碳、全氮和微生物等环境因子之间的关系较为复杂,还有待深入研究。
, CHAO Luo-Meng, ZHANG Li-Xin
The grasslands of the Inner Mongolian Stipa krylovii steppe make an important contribution to animal production in China. Soil respiration is an important component of the carbon cycle on the steppe and the main source of soil carbon losses. In this study, typical grassland dominated by S. krylovii, located in Maodeng Ranch, Xilinhaote, Inner Mongolia was selected as the study area. An open top type growth chamber was used to simulate the response of soil respiration to temperature and precipitation from 2011 to 2013. The Automated Soil CO2 Flux System (LI-8100, LI-COR, NE) was used to observe the soil respiration in the study area during the growing season (May to September). Air temperature, soil temperature and soil moisture were measured using the EM50 Data Collection System (Decagon Devices, Inc., NE, USA). Soil samples were collected, soil nutrients and microbial numbers were determined and correlation analysis used to explore associations between soil respiration rate and environmental factors. Soil respiration was significantly affected by air temperature and soil water content ( P<0.01). Under warm and warm-moist conditions respectively soil respiration rates were 2.60 μmol/(m2·s) and 1.96 μmol/(m2·s) (2012) and 3.38 μmol/(m2·s) and 2.09 μmol/(m2·s) (2013); differences between years were significant ( P<0.05). Increased air temperature and decreased soil water content both inhibited soil respiration. During the 2013 growing season additional environmental factors including soil organic carbon, total nitrogen and soil microbial mass may also have affected soil respiration. The Q10 temperature coefficient was significantly increased by warm-moist conditions. It was concluded that soil respiration is mainly influenced by hydrothermal conditions; increased air temperature and reduced soil moisture content inhibit soil respiration. The relationship between environment factors and soil respiration on the Inner Mongolian steppe is complex and requires further research.
土壤呼吸是指在未经扰动的土壤中产生CO2的所有代谢作用, 包括根系的自养呼吸以及微生物和土壤动物的异氧呼吸, 占生态系统呼吸的60%~90%。作为大气与陆地生态系统碳循环主要通量, 土壤呼吸是土壤碳输出的主要途径[1, 2]。土壤呼吸主要受到温度和水分等环境因子的影响[3, 4, 5, 6]。土壤呼吸温度敏感系数(Q10)是评价土壤呼吸作用的重要指标之一, Q10值的微小变化可能对土壤呼吸评价产生重要影响; Q10值的错估将导致对生态系统C循环预测的重大误差。了解温度及其他因素对Q10的影响是预测未来气候变化下的土壤C平衡的关键[7]。
草原是我国北方分布最广泛的自然生态系统, 构成了我国北方强有力的生态安全屏障[8]。克氏针茅(Stipa krylovii)草原作为典型草原中生境比较干旱的类型, 是亚洲中部草原所特有的草原群系, 是典型草原的代表类型之一[9]。许多研究对克氏针茅草原的植物物候期及其与气象因子的关系[10]、植物群落特征[11]、温度对克氏针茅草原生长季的生态系统碳通量影响[12]等进行了详细的描述。同时也有学者对内蒙古地区的不同类型草原土壤呼吸及其生物和非生物影响因子进行了大量研究[5, 13, 14, 15, 16, 17]。陈全胜等[14]在水热条件对锡林河流域典型草原退化群落土壤呼吸的影响研究中指出土壤呼吸与气温, 土壤温度之间呈指数相关, 与土壤水分之间有显著线性相关关系。李凌浩等[18]的研究结果也指出锡林浩特流域羊草(Leymus chinensis)草原土壤呼吸速率受温度和水分因子共同影响。
在全球变暖的大趋势下, 温度升高、降水时空变化都将对土壤呼吸产生一定的影响, 从而进一步影响到全球碳循环。但环境水热因子对内蒙古克氏针茅草原土壤呼吸的影响和增温条件下土壤呼吸温度敏感性变化的研究较为少见。因此, 本文拟解决问题如下:1)在增温增雨条件下内蒙古克氏针茅草原土壤呼吸的变化及与环境因子之间的关系; 2)增温增雨条件下内蒙古克氏针茅草原土壤呼吸温度敏感系数的变化及与环境因子之间的关系。所以理清增温和降水变化条件下土壤呼吸动态及其影响因素, 对草原碳收支研究具有重要意义。
本试验研究区域为内蒙古自治区锡林浩特市毛登牧场(距锡林浩特市东郊40 km)内蒙古大学草地生态学研究基地(44° 9'49″ E, 116° 29'2.3″ N), 海拔1102 m, 其生态类型为以克氏针茅建群的典型草原, 该区域为温带半干旱大陆性气候。该区年平均气温2.6℃, 最热月(7月)平均气温21.4℃, 最冷月(1月)平均气温为-19.0℃, 年平均最高温为24.9℃, 年平均最低温为-23.8℃, 50年间的平均年降水量为271.42 mm, 降水主要集中在生长季5-9月, 占全年降水量的87.3%。研究区土壤类型为栗钙土。
本试验于2011年7月在试验样地布置12个植物生长均匀的3 m× 3 m的试验小区, 采用开顶式生长室(OTC), 试验设置3个处理:CK为对照样地; CT为增温处理, CTW为增温增雨处理(在增温基础上对OTC内进行增雨处理), 每个处理分别进行4次重复。试验选取近50年(1961-2010年)生长季平均月降水量作为标准, 50年间年降水量波动很大, 将降水幅度设定为月平均降水量的20%[17]。
土壤温湿度的测定用温湿度数据采集器(温度探头型号:DS18B20; 湿度探头型号:EC-5)记录地下10, 20, 30 cm 土层土壤温湿度以及距地表20 cm 以上的地表空气温度, 每个处理4 个重复。
土壤呼吸测定采用开路式土壤碳通量测量系统(LI-8100, LI-COR, NE), 并在每个处理永久设置1个聚氯乙烯土壤呼吸环(内径10 cm, 高度5 cm), 埋入土壤, 插入地下2 cm, 地面露出3 cm。为消除测定土壤呼吸时植物自养呼吸的影响, 在测定前一天将每个土壤呼吸环内的植物地上部分剪掉, 待受扰动土壤稳定24 h后开始土壤呼吸速率的测定。测定时间为2011-2013年7-9月, 上午9:00-10:00, 每月测定2次。
土壤样品的采集在2013年7月下旬, 在不同处理每个小区取地下0~10 cm土壤样品, 自然风干后, 选用ATC-006 原子吸收分光光度计、ATC-165凯氏定氮仪和ATC-001 紫外分光光度计进行土壤有机碳氮的测定。土壤微生物于2013年9月下旬取各处理地表0~10 cm土层土壤用保温箱低温保存, 实验室内测定土壤中微生物细菌和放线菌的数量, 采用平板计数法测定, 细菌使用牛肉膏蛋白胨培养基; 放线菌使用高氏1号琼脂培养基(淀粉琼脂培养基)。
经过3年的增温处理, 不同处理间生长季平均气温存在显著差异(P< 0.01), CT和CTW较CK分别升高了2.17和1.45℃。同时生长季平均土壤湿度显著降低(P< 0.01), CT和CTW的平均土壤含水量相比CK降低了1.60%和0.47%。CT和CTW处理中, 生长季平均土壤温度与CK无显著差异(P> 0.05)(表1)。
| 表1 3年生长季OTC增温效果 Table 1 The warming effect of OTC in growing season of three years |
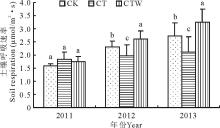
以2011-2013年5-9月测定的土壤呼吸进行年际变化分析结果表明(图1), 2011年生长季CT、CTW和CK分别为1.84, 1.76, 1.59 μ mol/(m2· s); 2012年生长季CK和CTW土壤呼吸速率均高于CT, 且CT土壤呼吸速率显著地低于CTW(P< 0.05), 生长季土壤呼吸平均值大小排序为CTW > CK > CT (2.60 μ mol/m2· s > 2.30 μ mol/m2· s > 1.96 μ mol/m2· s); 2013年生长季土壤呼吸速率的变化规律与2012年相同, CK高于CT而低于CTW, 并且CT的降低幅度显著(P< 0.05)。CK、CT、CTW的土壤呼吸速率分别为2.72, 2.09和3.38 μ mol/(m2· s)(图1)。
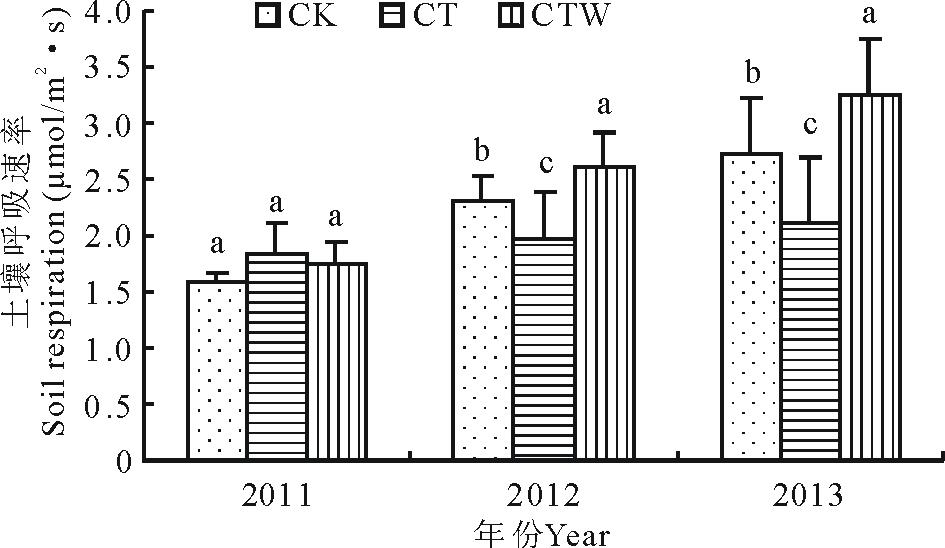 | 图1 不同处理间土壤呼吸年际之间的变化. 不同字母之间表示差异显著(P< 0.05)。Fig.1 Soil respiration change of different treatment and interannual. Significant between different letters (P< 0.05). |
不同处理下的土壤呼吸受到年际间影响差异不显著(P> 0.05), 试验样地3年的监测中, CTW对土壤呼吸速率的促进作用非常显著(P< 0.01), CT对土壤呼吸速率的抑制效果不显著(P> 0.05)。
2.3.1 Q10值与土壤温度、土壤含水量的关系 土壤呼吸温度敏感系数(Q10)表示土壤呼吸速率与温度的关系, 根据土壤呼吸与土壤温度间的关系计算(表2)。CT和CTW在开始增温(2011年)时CTW下最高, Q10值为6.49, 随着增温时间的增长(2012年)下降为3.04; 在2011年生长季CT的Q10值为6.48, 高于CK而低于CTW处理下; 但是在2012年生长季, Q10降低, 为2.36。2013年生长季, CT处理则低于CK处理, 两者的Q10值分别为4.49和3.70(表2)。
| 表2 不同处理的土壤呼吸温度敏感性(Q10)和土壤温湿度(T10、W10)及其相关性分析 Table 2 The temperature sensitivity of soil respiration (Q10) and soil temperature and humidity (T10, W10) under different processing and the correlation analysis between them |
将土壤温度和土壤含水量与Q10值做相关性分析, 发现Q10值与土壤温度和土壤含水量的关系不具有较好的相关性(表2)。
2.3.2 土壤碳氮含量与土壤微生物数量变化及与Q10关系 2013年生长季中, CT和CTW对土壤全氮和土壤有机碳含量的影响不显著(表3)。土壤有机碳含量分别为1.24%和1.23%, 土壤全氮含量分别为0.16%和0.15%。生长季末CT和CTW中土壤微生物生物量较CK减少(P< 0.05), CT中土壤微生物生物量降低51.9%, 微生物生物量为3.39× 106个; CTW的土壤微生物生物量的下降幅度为61.2%, 生物量为2.73× 106个。CT中土壤真菌和土壤放线菌数量显著降低(P< 0.05), 分别为1.88× 105个和1.17× 106个, 降低65.4%和12.5%; 且CTW对土壤真菌和放线菌数量的影响极为明显(P< 0.01), 生物量分别为1.48× 105个和9.89× 106个, 下降72.8%和26.1%。但是对于土壤细菌而言, CT和CTW对其影响不显著(P> 0.05), 分别为3.31× 106个和3.22× 106个。
| 表3 不同处理土壤有机碳、土壤全氮含量及土壤微生物生物量 Table 3 Different treatments soil organic carbon, soil total nitrogen content and soil microbial biomass |
在2013年生长季对土壤温度敏感系数Q10和土壤有机碳和全氮以及土壤微生物总生物量进行相关分析, 结果表明Q10与土壤有机碳氮与微生物量间均没有显著相关关系(图2)。
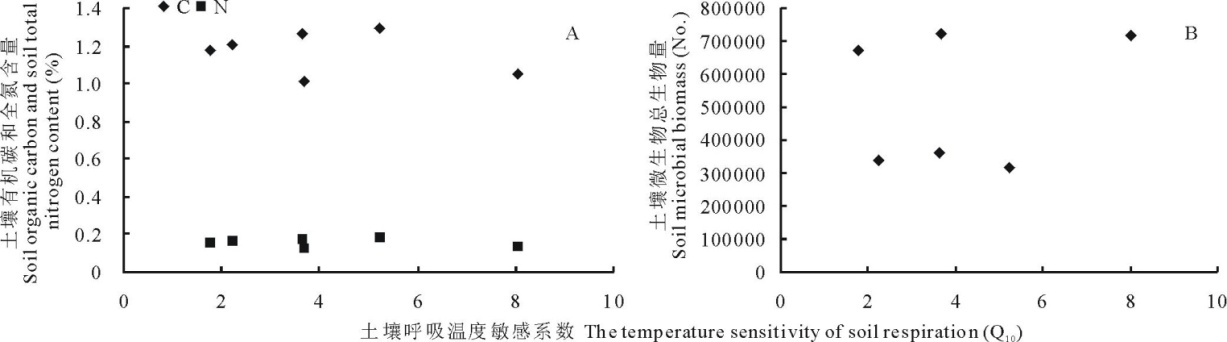 | 图2 土壤呼吸温度敏感系数和土壤有机碳、全氮以及土壤微生物之间的关系Fig.2 The relationship between soil organic carbon, soil total nitrogen content, soil microbial biomass and Q10 |
将不同处理样地内土壤呼吸分别与空气温度、土壤温度和土壤含水量进行相关分析, 结果如表4所示, 在不同环境因子中, 土壤呼吸与土壤温度之间表现出显著正相关关系(P< 0.05)。在CK、CT、CTW内, 土壤呼吸与土壤温度之间的相关系数分别为0.50, 0.40和0.11, 表明增温效果显著, 土壤呼吸与土壤温度之间的相关性逐渐增高。相关性分析表明, 土壤呼吸与土壤温度之间符合指数函数关系, 随着增温幅度的提高, 相关性系数R2表现出升高的趋势(表4)。
| 表4 不同处理下土壤呼吸与土壤温度、土壤含水量以及土壤呼吸与空气温度的拟合模式 Table 4 The fitting model of soil respiration with soil temperature, soil moisture content, soil respiration and the air temperature under different processing |
不同处理土壤呼吸与土壤含水量之间也存在显著正相关关系(P< 0.05)。相关性系数的大小依次为0.51< 0.62< 0.81(CTW< CK< CT)。土壤呼吸与土壤含水量之间随着温度的增高, 同样表现出逐渐增高的趋势。相关性分析表明土壤呼吸与土壤温度之间存在线性函数关系, 相关性系数R2也表现出增大的趋势。
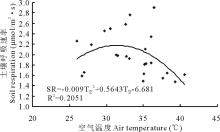
对内蒙古克氏针茅草原2011-2013年生长季土壤呼吸与空气温度的数据进行相关性分析表明, 不同处理土壤呼吸(SR)和空气温度(T空)符合二次项函数关系, 空气温度对土壤呼吸的影响呈现先升高后降低的趋势, 即土壤呼吸与空气温度之间存在一个阈值点(本试验中为31.4℃), 未达到之前土壤呼吸随空气温度的增加而增加, 超过阈值之后随空气温度的增加而降低(图3)。
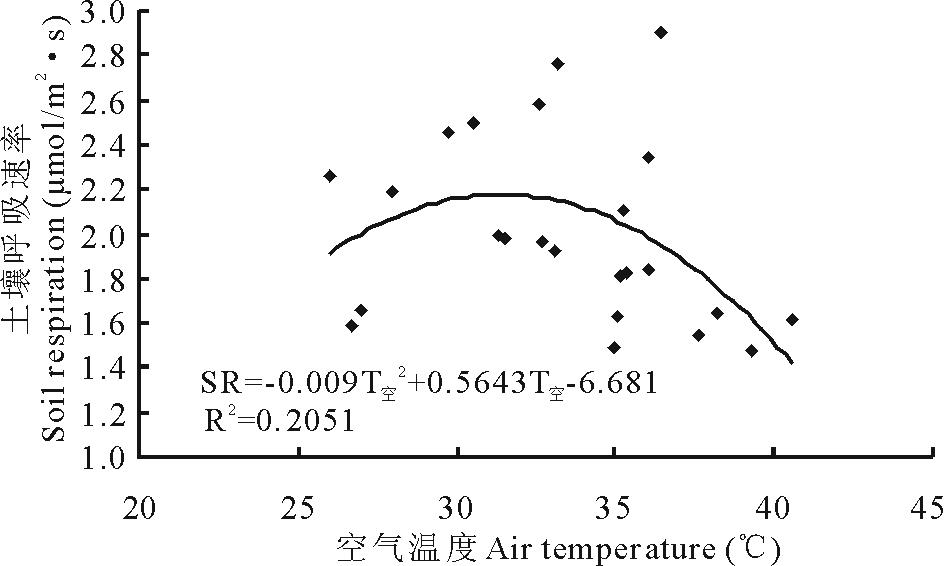 | 图3 土壤呼吸与空气温度的关系Fig.3 The relationship between the air temperature and soil respiration |
土壤水分是温带半干旱典型草原生态系统中非常重要的土壤物理特性之一, 也是土壤发育中最为活跃的影响因素。土壤中的水分对生态系统的物质循环和能量流动、植物生长发育均具有重要的作用。土壤含水量对土壤呼吸有很大的影响, 尤其在干旱或半干旱地区水分会取代温度成为土壤呼吸的最主要限制因子[21, 22]。本研究中, 不同处理下土壤呼吸与土壤含水量均呈显著正相关关系。在对照处理下, 土壤呼吸与土壤含水量的相关性系数低于增温处理而高于增温增雨处理, 说明在温度增高的条件下, 土壤含水量对土壤呼吸有较大的影响。根据土壤呼吸水分限制的理论[23], 土壤水分过低, 对植物根呼吸产生一定影响, 会抑制土壤呼吸。本研究结果表明不同处理下土壤含水量差异显著(P< 0.01), 对照、增温和增温增雨处理土壤含水量依次为10.73%, 8.96%, 10.13%, 土壤含水量随气温的升高而降低。原因是气温升高加剧植物蒸腾作用和土壤水分的蒸散, 使得土壤含水量降低, 进而导致土壤呼吸降低。
内蒙古克氏针茅典型草原为温带半干旱大陆性气候, 受海拔等因素影响, 日间光照强烈, 夜间降温迅速, 因此气温成为该地区生态系统能量流动和物质循环的主要影响因子。研究显示, 温度与土壤呼吸的关系因研究对象和研究地点不同而有所差异, 但大部分可以用指数模型来表示[3, 6, 22]。土壤呼吸受多个环境因子共同作用影响, 本研究中, 土壤呼吸与气温之间存在显著二项函数关系。在一定阈值内气温的升高会导致土壤呼吸的增加, 但是超过阈值上限后, 土壤呼吸会随着气温的升高而减弱。在阈值之前, 气温作为主要影响因子, 促进植物蒸腾作用和土壤水分的蒸散和植物新陈代谢, 以及土壤中活性碳库向钝性或缓性碳库的转移, 从而使得土壤呼吸增强; 但超过阈值后, 土壤含水量成为主要限制因子, 对植物产生水分胁迫, 植物根呼吸速率减弱, 从而使得土壤呼吸降低。这与Holthausen和Caldwell[24]对滨藜属植物根呼吸季节动态研究结果中温度与根呼吸的关系相似。
土壤环境主要的研究对象包括土壤有机化合物的类型及其含量、微生物种群及活性、植物的元素背景值等[25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35]。研究表明土壤呼吸与土壤有机碳、全氮和土壤微生物之间有显著相关关系, 对土壤呼吸或促进[26, 27]、或无影响[28]。Niu等[29]在对温带草原氮素影响研究中指出氮素的添加对生态系统呼吸有促进作用, 鲍芳和周广胜[34]在中国草原土壤呼吸研究进展中也指出土壤有机碳和全氮是影响我国温带草原土壤呼吸的主要因素之一。Bazzaz和Williams[30]在对大气CO2浓度对混交林幼苗生长中的研究中指出, 当土壤中的有机质含量、根系生物量、微生物活性增加时, 其土壤呼吸速度显著增加。土壤微生物是土壤生态系统中养分来源的原动力, 在动植物残体的降解和转化, 养分的释放和循环及改善土壤理化性质中起着重要作用[36]。本研究发现增温和增温增雨条件下土壤微生物数量降低, 但土壤有机碳和全氮含量没有显著变化, 土壤呼吸与土壤有机碳和全氮含量以及土壤微生物数量没有显著相关关系。原因可能是土壤水分是本文研究中影响土壤呼吸的最主要限制因子, 其他环境因子对于土壤呼吸的影响在不同水分条件下差异不明显。
研究表明, 影响Q10值的因素有温度、水分、呼吸底物、土壤微生物、土壤矿物学性质和土壤质地等[11]。谭炯锐等[31]发现北京大兴杨树人工林土壤呼吸Q10值与土壤温度呈负相关关系; 在土壤含水量为8%~10%的条件下Q10达到最大值, 土壤含水量继续增大时Q10开始降低, 呈现先增加后降低的趋势。王小国等[32]对四川盆地3种土地利用方式的研究中发现Q10值与土壤温度呈现负相关, 但是与土壤含水量呈现正相关关系。张金波等[33]在三江平原不同土地利用方式的土壤呼吸敏感性差异及其影响因素分析中指出Q10值与土壤有机碳、水溶性有机碳和微生物量呈显著的线性关系。本研究结果表明, Q10与土壤温度和土壤含水量、土壤有机碳、全氮及土壤微生物之间无显著关系。本实验结果中, 2011年Q10值在模拟增温处理中高于对照处理, 而在2012和2013年模拟增温处理下Q10值反而低于对照处理, 模拟增温增雨下高于对照处理。土壤有机碳和全氮含量以及土壤微生物数量与Q10值之间的关系有待于进一步验证。土壤呼吸温度敏感性系数是生态系统综合作用的结果, 单独的温度、湿度或者土壤有机碳、全氮等不能对其产生一定的影响, 因此对土壤呼吸敏感性在增温条件的响应机制还有待于进一步研究。
克氏针茅草原土壤呼吸受水热条件影响。本研究表明, 土壤呼吸主要受水热条件影响, 并且空气温度升高和土壤含水量的降低均对土壤呼吸有抑制作用, 增温增雨处理下土壤呼吸速率要高于增温处理, 2012年增温增雨与增温处理下土壤呼吸速率分别为2.60和1.96 μ mol/(m2· s), 2013年增温增雨与增温处理下土壤呼吸速率分别为3.38和2.09 μ mol/(m2· s), 且差异显著(P< 0.05)。土壤呼吸与土壤含水量、土壤温度间具有较好的相关性, 本研究中增温增雨处理、对照样地、增温处理土壤含水量与土壤呼吸的相关性系数R2分别为0.51, 0.62, 0.81; 土壤呼吸与土壤温度的相关性系数R2 分别为0.11, 0.40和0.50。土壤呼吸温度敏感系数与土壤温度、土壤含水量、土壤有机碳、全氮和微生物等环境因子之间的关系不确定。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [22] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|