



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
四个抗草甘膦基因的抗性比较
[刘健1, 2**, *  , 肖雅文
, 肖雅文2**, * , 芦佳1, 2**, * , 吴忠义2 , 张中保2 , 徐杰1*, * , 姚磊2*, * ]
, 肖雅文, 芦佳, 吴忠义, 姚磊]
|
|


作者简介:刘健(1988-),男,山西吕梁人,在读硕士。E-mail: 906047892@qq.com。肖雅文(1988-),女,四川成都人,硕士。E-mail: xyw_1020@126.com。芦佳(1987-),女,山西大同人,在读硕士。E-mail: 905696441@qq.com。
草甘膦抗性是当代农业重要的农艺性状。为了选择最佳的抗性基因用于植物筛选标记,通过选取 cp4 -epsps、 GR79 -epsps、 gat4621和 GAT四个草甘膦抗性基因进行比较,在拟南芥中验证各基因对草甘膦的抗性。4个基因的转化苗经4个浓度的草甘膦喷施检测,结果显示, GAT类基因的抗性明显高于 epsps类基因,基因间的差异达到了极显著水平。 gat4621基因对草甘膦的抗性最好,在各浓度下存活率均表现为最高值,且抗性稳定。国内发掘的 GAT基因对草甘膦也表现出良好的抗性。 GAT类基因的拟南芥转化苗在喷施高浓度草甘膦条件下出现严重抽苔抑制;但在停施草甘膦后抑制解除,且可以开花结实。转 epsps类基因可以获得高抗草甘膦植株,但在花期喷施草甘膦将造成败育。从基因大小及抗性强弱考虑, GAT类基因作为筛选标记基因应该更为适合。
, XIAO Ya-Wen, LU Jia, WU Zhong-Yi, YAO Lei
Glyphosate tolerance is an important agronomic trait in modern agriculture. In order to determine the best glyphosate tolerance gene for plant selectable markers we compared four glyphosate tolerance genes, cp4 -epsps, GR79 -epsps, gat4621 and GAT, in Arabidopsis. Four transgenic plants harboring each gene were treated with four different rates of glyphosate. The results showed that the resistance of plants with GAT gene was significantly higher than plants with the epsps gene. Plants with the gat4621 gene had the highest survival rate at all herbicide rates. The plants with the domestic GAT gene also had strong resistance. Reproductive development of Arabidopsis plants with the GAT gene was suppressed at higher glyphosate application rates.It was concluded that the GAT gene was more suitable as a selectable marker gene.
草甘膦(glyphosate)是由美国孟山都公司开发的一种内吸传导型除草剂, 具有无选择性、无残留、灭生性的特点, 特别是对灭除多年生杂草非常有效。植物绿色部分均能很好地吸收草甘膦, 以叶片吸收为主。草甘膦通过与磷酸烯醇式丙酮酸(phosphoenolpyruvate, PEP)竞争而和5-烯醇式丙酮酸莽草酸-3-磷酸合成酶(5-enolpyruvylshikimate-3-phosphate synthase, EPSPS)、莽草酸-3-磷酸(shikimate-3-phosphate, S3P)形成稳定的复合体EPSPS-S3P-草甘膦, 使得芳香族氨基酸化合物合成受阻, 促使黄酮以及酚类化合物等激素和关键代谢物失调[1], 打乱了生物正常氮代谢而导致生物体的死亡[2]。
随着当前世界农业生产向着机械化和集约化方向迅速发展, 抗除草剂转基因作物成为一种普遍的种植需求。目前, 植物获得草甘膦抗性主要是通过两种不同的途径[3]。一种是通过诱变或向植物体内导入对草甘膦不敏感的EPSPS, 进而提高植物对草甘膦的耐受性。另一种是通过向植物体内导入草甘膦降解基因, 在草甘膦发挥作用前将其降解, 进而使植物对草甘膦产生抗性。EPSPS是草甘膦作用于植物产生抑制的关键性合成酶。在提高植物对草甘膦耐受性的研究中多数是致力于对草甘膦不敏感的EPSPS发掘。当前世界范围内应用最广的是来自根癌农杆菌(Agrobacterium tumefaciens)CP4菌株的EPSPS[4]。因其对PEP亲和性较好且高抗草甘膦, 转cp4-epsps基因的植物对草甘膦具有较高的抗性, 在抗草甘膦作物中应用最多[3]。草甘膦降解机制与牺牲EPSPS和PEP结合催化活性为代价而获得对草甘膦抗性不同, 是在草甘膦发生作用前将其降解以使植物产生抗性。在长期受草甘膦污染的土壤中存在许多可降解草甘膦的细菌[5]。多种基因已经被发现对草甘膦具有分解作用。草甘膦N-乙酰转移酶(glyphosate N-acetyltransferase, GAT)基因即为其中一种。研究发现, 草甘膦在GAT的作用下转化成对植物没有毒害的N-乙酰草甘膦(N-acetylglyphosate, NAG)[6]。现在先锋公司的转gat4621抗草甘膦作物已得到大面积商业种植[3]。目前商业化的抗草甘膦作物所利用的抗性基因均属国外专利。国内对草甘膦抗性基因的研究以中国农业科学院生物技术研究所获得抗性基因相对较多。林敏实验室分离到的GR79(或称AM79 aroA)[7]的草甘膦抗性表现较好, 被推测具有较大的商业应用价值[8]。此外, 该实验室还分离获得了国内唯一的草甘膦降解基因GAT, 转GAT的烟草(Nicotiana tabacum)获得较好的草甘膦抗性[9, 10]。
植物筛选标记基因是植物遗传转化体系中重要的组成部分。因植物材料的不同, 最佳筛选标记基因的选择有所不同[11]。最适的筛选标记基因对植物的遗传转化能起到事半功倍的作用。因cp4-epsps基因对草甘膦具有高度抗性, 被建议用作植物筛选标记基因[12]。本实验室已构建一套含多种筛选标记基因的植物表达双元载体[13], 寻找最佳的抗草甘膦基因用于构建载体的植物筛选标记是丰富现有载体的目标。虽然cp4-epsps和gat4621基因已经在商业生产中得以应用。但未见报道两类基因对草甘膦的抗性是否存在差异。本研究选择cp4-epsps、GR79-epsps、gat4621和GAT四个草甘膦抗性基因进行比较, 在拟南芥(Arabidopsis thaliana)中验证各基因对草甘膦的抗性, 以期获得最佳的抗性基因用于载体的构建。
1.1.1 植物材料与菌株 野生型拟南芥Columbia(Col-0), 大肠杆菌菌株DH5α 和根癌农杆菌GV3101:pMP90都为北京市农林科学院农业生物技术研究中心保存。
1.1.2 载体、酶和试剂 本实验使用的各种限制性内切酶、M-MLV反转录酶、T4连接酶、去磷酸化酶购自TaKaRa、Promega和NEB公司。高保真酶Phusion购自Thermo公司。Easy Taq购自TransGen Biotech公司。PCR纯化试剂盒、胶回收试剂盒、质粒小提试剂盒购自博迈德有限公司。引物合成及测序由中美泰和生物技术(北京)有限公司完成。
1.2.1 草甘膦抗性基因的获得 根据孟山都公司的pMON植物转化载体序列[14], 设计引物AtPT-5'和CP4-3'(本试验所用引物序列见表1, 下划线部分为酶切位点), 从抗草甘膦玉米基因组中扩增cp4-epsps基因。该cp4-epsps基因序列5'端含有1段225 bp的拟南芥epsps基因的引导肽CPT序列。
根据中国专利ZL200710177090.1中GR79序列, 由Invitrogen公司根据植物偏爱密码子优化碱基序列并合成。为保持与上述cp4-epsps基因相同, 通过融合PCR方法在GR79的5'端添加CPT引导肽序列。方法是用引物AtPT-5'和AtPTGR79-R以cp4-epsps基因为模板扩增CPT引导肽序列; 用引物AtPTGR79-F和GR79-3'扩增GR79基因。琼脂糖凝胶电泳纯化PCR产物后, 以纯化产物为模板, 将CPT和GR79基因混合, 再用引物AtPT-5'和GR79-3'扩增, 获得5'端添加CPT引导肽序列的GR79-epsps基因。
根据美国专利US 7973218-B2中gat4621序列, 基因由中美泰和生物技术(北京)有限公司合成, 并克隆至pBluescript SK载体SacⅠ (基因5'端)和KpnⅠ (基因3'端)位点。
根据中国专利200510086626.X中GAT序列, 并选用单子叶植物偏爱密码子优化碱基序列后命名GATm。基因GATm由中美泰和生物技术(北京)有限公司合成, 并克隆至pBluescript SK载体SacⅠ (基因5'端)和KpnⅠ (基因3'端)位点。
| 表1 试验中所用PCR引物 Table 1 PCR primers used in the experiments |
1.2.2 植物转化双元载体的构建 用SacⅠ 和KpnⅠ 分别双酶切cp4-epsps和GR79-epsps含CPT的PCR产物及含gat4621和GATm的pBSK载体, 琼脂糖凝胶电泳纯化后备用。用SacⅠ 和KpnⅠ 双酶切植物表达载体pYBA1305[15](pYBA100衍生载体, 植物筛选标记基因为bar, 含CaMV 35SΩ 启动子和CaMV polyA终止子的植物表达框), 琼脂糖凝胶电泳纯化后分别与上述各基因片段连接。连接产物转化大肠杆菌DH5a, 挑选克隆提取质粒DNA酶切验证并测序, 得到4个基因用于植物转化的双元载体, 各载体的T-DNA区段见图1。
1.2.3 拟南芥的遗传转化及阳性苗的筛选 2013年, 利用电击法将各载体转入农杆菌GV3101:pMP90中, 采用农杆菌介导的花序浸渍法侵染拟南芥[16], 每个基因各侵染32株野生型拟南芥。将收获的各基因转化T0代种子分别均匀播撒在3盘盛有营养土的培养托盘中, 置于22℃(昼)/18℃(夜), 光照周期为16 h(光)/8 h(暗)的人工气候室。待幼苗长出两片子叶后喷洒2~3次浓度为0.2%(体积比)的商业Basta除草剂筛选阳性苗。存活植株用小量快速法[17]提取植物基因组DNA进行PCR扩增检测。用引物AtPT-5'和CP4-3'检测cp4-epsps转化苗; 扩增出1.8 kb目的条带为阳性苗。用引物AtPTGR79-F和GR79-3'检测GR79-epsps转化苗; 扩增出1.5 kb目的条带的为阳性苗。用引物GAT4621-F/R检测gat4621转化苗; 扩增出约269 bp目的条带为阳性苗。用引物GATm-F/R检测GATm转化苗; 扩增出约245 bp目的条带为阳性苗。
1.2.4 草甘膦抗性的鉴定 待喷洒Basta除草剂获得阳性苗后, 对4个基因的转基因苗按照浓度从低至高逐步喷施不同浓度的孟山都公司“ 农达” 除草剂(Roundup, 41%草甘膦异丙胺盐水剂)稀释液。稀释浓度梯度为0.2%, 0.4%, 0.6%, 0.8%(体积比), 每隔3 d喷1次, 每个梯度喷2次。统计4个基因的转化株在每个草甘膦浓度下的存活率, 存活率=存活株数/Basta抗性株。每个基因3盘苗, 为3次重复进行统计。
1.2.5 基因表达量的定量PCR检测 在0.4%浓度“ 农达” 处理下, 4个基因的转基因苗对草甘膦的抗性差异已经非常明显。随机选取各基因的高抗与弱抗苗各1株, 用TRIzol法提取总RNA, 用M-MLV翻转成cDNA。以拟南芥SAND基因(AT2G28390)为内参进行基因表达量的荧光相对定量检测。基因检测引物为CP4-F/R、GR79-F/R、GAT4621-F/R、GATm-F/R和SAND-F/R。
各基因在不同草甘膦浓度下的抗性差异, 用SPSS软件进行差异显著性方差分析。
用引物AtPT-5'和CP4-3'以抗草甘膦玉米基因组DNA为模板, 扩增获得cp4-epsps基因。经GenBank的Blast比对, 该cp4-epsps基因序列5'端含1段与拟南芥epsps基因(AT2G45300)前225 bp同源性100%的引导肽序列; 其cp4-epsps基因区段与抗草甘膦大豆中cp4-epsps基因(AF464188)核酸序列同源性最高, 为99.64%, 氨基酸同源性为100%。该cp4-epsps基因PCR产物经SacⅠ 和KpnⅠ 双酶切后连入植物表达载体pYBA1305; 酶切及测序验证正确后命名1305-cp4-epsps。
由Invitrogen公司根据植物偏爱密码子合成的GR79序列与原始序列核酸的同源性为72.48%, 氨基酸同源性为100%。通过融合PCR添加CPT引导肽后, 该GR79-epsps基因PCR产物经SacⅠ 和KpnⅠ 双酶切后连入植物表达载体pYBA1305; 酶切及测序验证正确后命名1305-GR79-epsps。
基因gat4621合成后用SacⅠ 和KpnⅠ 双酶切相应载体, 纯化后的基因连入植物表达载体pYBA1305; 酶切及测序验证正确后命名1305-gat4621。
根据单子叶植物偏爱密码子优化后, GATm与原始序列核酸的同源性为82.17%, 氨基酸同源性为100%。基因GATm合成后用SacⅠ 和KpnⅠ 双酶切相应载体, 纯化后的基因连入植物表达载体pYBA1305; 酶切及测序验证正确后命名1305-GATm。
 | 图1 载体T-DNA区段示意图Fig.1 Structure of the T-DNA region of vectors A: 载体1305-cp4-epsps Vector 1305-cp4-epsps; B: 载体1305-GR79-epsps Vector 1305-GR79-epsps; C: 载体1305-gat4621 Vector 1305-gat4621; D: 载体1305-GATm Vector 1305-GATm. |
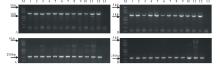
利用电击法将4个载体分别转入农杆菌GV3101:pMP90中, 采用农杆菌介导的花序浸渍法侵染拟南芥[16]。将每个载体收获的拟南芥T0代种子播种于3盘盛有营养土的托盘中置于人工气候室培养。因pYBA1305载体的植物筛选标记基因为Bar, 转基因苗首先应具有草胺磷抗性。幼苗经喷洒2~3次Basta除草剂筛选后, 能够继续生长且长势良好的植株为抗性株。从各载体的抗性苗中随机抽取12株提取植物基因组DNA进行PCR验证, 结果全部为阳性(图2), 证明目的基因已整合到拟南芥植物基因组中。经统计4个载体1305-cp4-epsps、1305-GR79-epsps、1305-gat4621和1305-GATm分别平均每盘各获得转化苗(185.7± 18.6), (192.0± 23.5), (145.3± 43.5)和(119.0± 8.5)棵; 平均每转化1株拟南芥获得11~18株T1代转化苗。
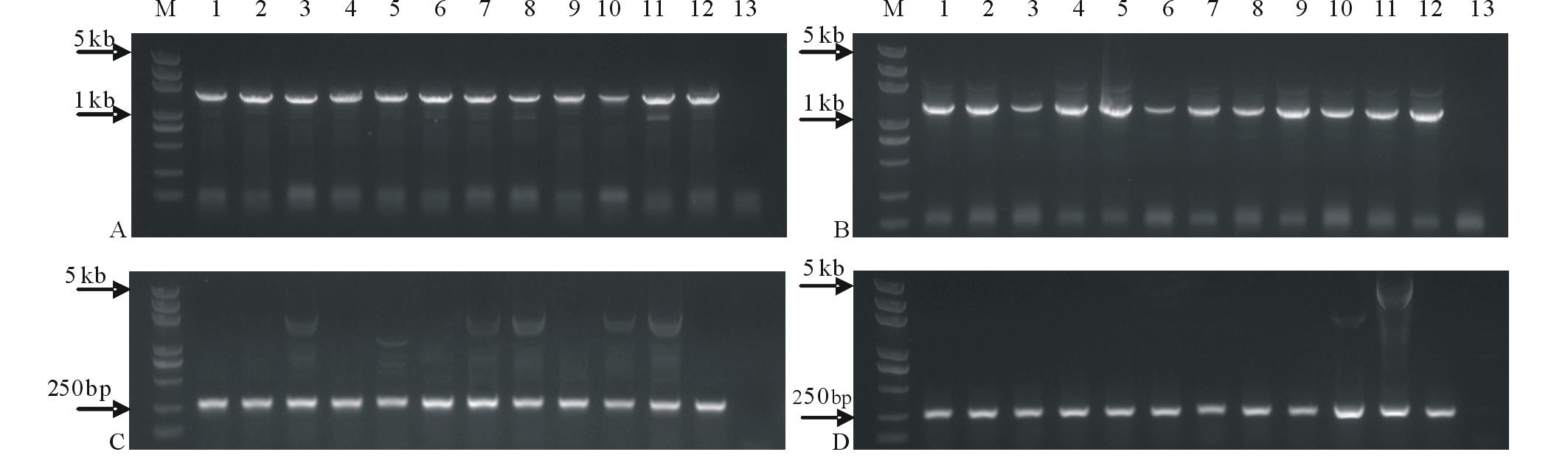 | 图2 转基因阳性苗4种抗性基因的PCR检测Fig.2 The PCR detections of four glyphosate-resistant genes from transgenic plants A:cp4-epsps; B:GR79-epsps; C:gat4621; D:GATm; M: DNA分子量标记DNA ladder; 1~12:不同转化植株Different transgenic plants; 13:空白对照Blank control. |
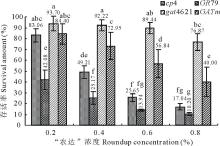
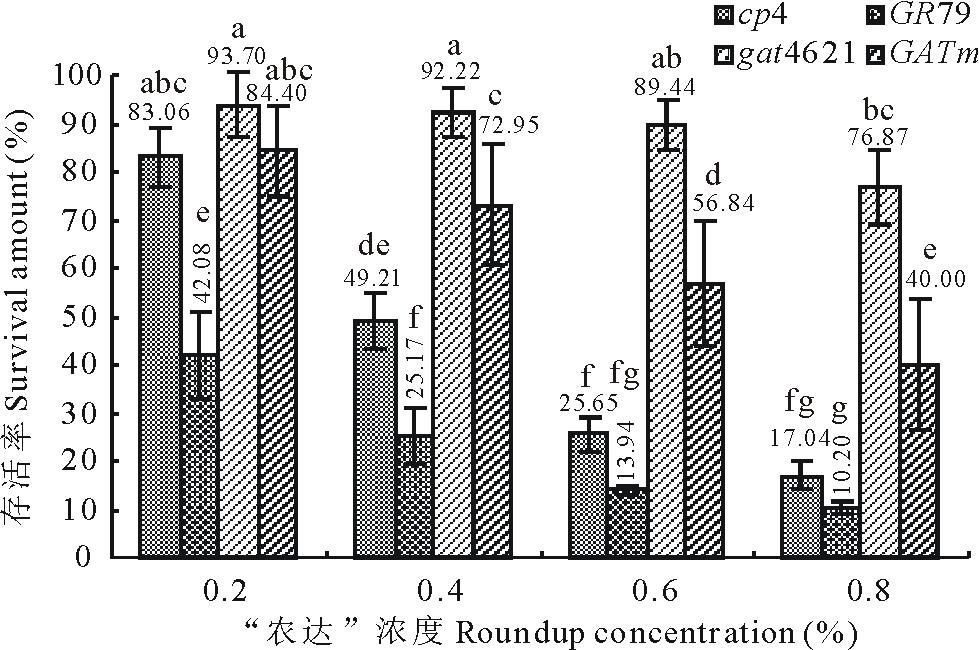 | 图3 不同草甘膦浓度梯度下各基因转化苗的存活率 不同字母表示差异显著(P< 0.05)。Fig.3 Transgenic plants survival amount on different concentrations of glyphosate The different letters mean significantly different (P< 0.05). |
在先经除草剂Basta筛选的基础上, 分别对4个载体的转化苗逐步喷施了0.2%~0.8%四个浓度梯度的“ 农达” 。转化苗喷施草甘膦后的存活率统计结果见图3, 生长情况见图4。由图3和图4可以看到各浓度下GAT类(gat4621和GATm)转化苗的存活率都较EPSPS类(cp4-epsps和GR79-epsps)转化苗的存活率高。结果显示浓度间和基因间的差异都达到了极显著的水平(P< 0.05)。相比低浓度的0.2%, 当浓度提升至0.6%时差异开始显著。4个基因间的差异都达到了显著性水平(P< 0.05); 其中抗性最好的gat4621与其他3个基因的差异达到极显著水平。
对4个“ 农达” 浓度和4个基因组成的全部16个处理的存活率进行单因子方差分析(图3)。gat4621基因对草甘膦的抗性最好, 在各浓度下存活率均表现为最高值, 且抗性稳定, 只在0.8%浓度时才有所下降, 差异达到显著性水平。GATm基因对草甘膦也表现出良好的抗性, 但随着浓度的提高抗性逐渐下降, 浓度达到0.6%时基因内差异达到了显著水平; 其与gat4621基因的抗性在0.2%的浓度时无差异。
 | 图4 不同草甘膦浓度梯度下各基因转化苗的存活情况Fig.4 The growth situation of transgenic plants of different genes on the different concentrations of glyphosate |
 | 图5 喷施草甘膦后GAT类和epsps类基因转化苗的抽苔情况Fig.5 The bolting of transgenic plants of GAT or epsps genes A: GATm转化苗 The transgenic plants of GATm; B: cp4-epsps转化苗The transgenic plants of cp4-epsps. |
epsps基因转化苗对草甘膦相对较敏感。在喷施0.2%的“ 农达” 后, GR79-epsps基因转化苗的存活率急剧下降至42.08%。cp4-epsps基因转化苗的存活率虽然达到83.06%, 但其中许多苗出现心叶变黄不良症状(图4)。当“ 农达” 浓度达到0.4%时, cp4-epsps基因转化苗中前期表现不良症状的苗死亡, 存活率降到49.21%。当浓度提高到0.8%时, cp4-epsps基因转化苗的存活率下降到17.04%。各浓度下GR79-epsps基因转化苗的存活率最低, 且均约为cp4-epsps基因转化苗存活率的50%, 在低浓度时差异达显著水平。
由于在长日照条件下拟南芥Col-0生态型生长3~4周后开始抽苔。当用0.2%浓度的“ 农达” 处理转化苗时生长时期已经接近3周, 部分苗已经开始抽苔。随着施用“ 农达” 浓度的提高和生长期的变化, epsps高抗转化苗陆续抽苔开花, 但GAT转化苗表现出抽苔受到抑制, 叶片颜色变暗(图4)。但在停止继续喷施草甘膦后, 生长受到抑制的转化苗叶片逐渐转绿, 并陆续抽苔结实(图5A)。epsps存活的高抗转化苗在喷施草甘膦后抽苔没有受到影响, 叶片颜色较绿(图5B); 但在种子收获时发现所有的果荚都没有结实(图6A), 没有收到1粒种子。在体式显微镜下拨开epsps转化苗的果荚可以看到所有的胚珠都在早期停止了发育, 而GAT转化苗的果荚内胚珠发育成正常的种子(图6B)。将GAT转化苗收获的种子播种于人工气候室, 经喷洒除草剂Basta后, 存活的抗性苗在不喷施除草剂“ 农达” 的条件下可以正常抽苔结实。
 | 图6 epsps类和GAT类基因转化苗的果荚结实情况Fig.6 The siliques of transgenic plants of GAT or epsps genes A: cp4-epsps转化苗 The transgenic plant of cp4-epsps; B: GATm转化苗 The transgenic plant of GATm. |
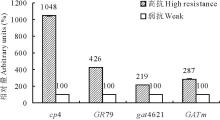
当“ 农达” 浓度提升到0.4%时, 4个基因的转化苗都出现生长受到抑制幼苗。各基因转化苗中随机选取高抗和弱抗幼苗各1株, 提取叶片总RNA并反转录成cDNA。在检测RNA无基因组DNA污染后, 用此cDNA为模板, 以SAND为内源参照基因进行抗性基因表达的荧光相对定量分析(SAND基因引物见表1)。结果如图7所示, 4个基因分别在高抗株中的表达量远高于弱抗株。为了明确高抗株的高抗性是否源于拷贝数的增加而引起。分别对gat4621和GATm各16个高抗转化株T2代幼苗进行了抗除草剂Basta分离比统计。gat4621和GATm各有75.0%和43.8%的转化株系符合3∶ 1分离比, 这些株系可以初步认为是单拷贝插入。
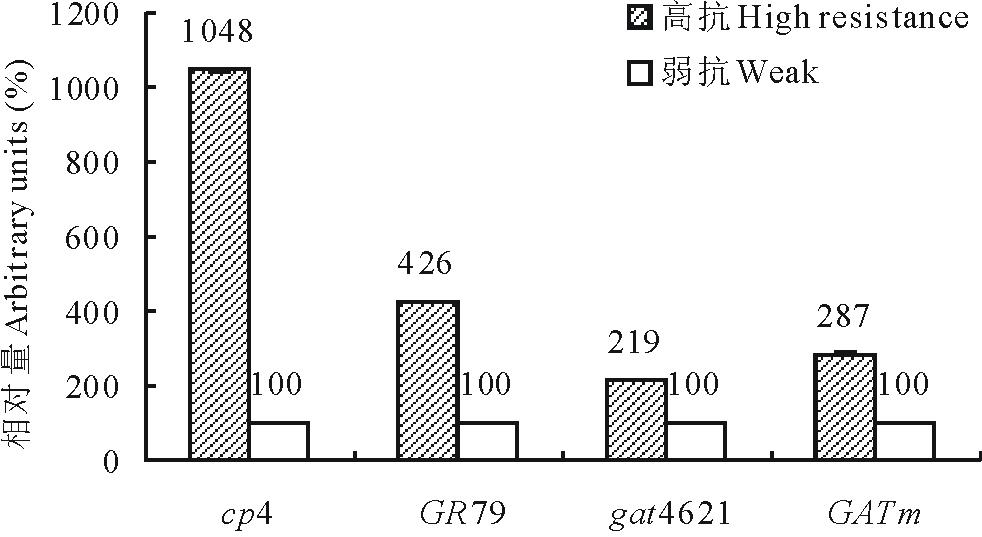 | 图7 转基因苗的基因表达量分析Fig.7 The analysis of genes expression of transgenic plants |
草甘膦抗性是重要的农艺性状。本研究选取两类4个草甘膦抗性基因构建于同一载体的相同克隆位点, 且epsps类基因都含有相同的CPT引导肽序列, 相对客观地体现了该4个基因对草甘膦的真实抗性。从T1代转化苗喷施“ 农达” 后的存活率来看, GAT类基因的抗性明显高于epsps类基因。先锋公司的gat4621是由地衣芽孢杆菌(Bacillus licheniformis)3个菌株的GAT基因经体外重组(DNA shuffling)后大量筛选获得。此外, 为了在植物中更好表达该蛋白, 一个由GCT编码的丙氨酸被插在蛋白质氨基酸序列的第二位[18]。国内发掘的GAT蛋白氨基酸序列第二位不含有插入的丙氨酸, 也表现出对草甘膦的良好抗性, 且在施用低浓度草甘膦时抗性已经接近gat4621, 其在将来的农业生产上应具有广阔的应用前景。GAT类基因的转化苗在喷施高浓度“ 农达” 条件下存活率高, 但严重抑制抽苔。由于在不施用草甘膦时GAT转基因苗能够正常抽苔, 这种抽苔抑制可能是由于中间产物NAG不能迅速降解而积累所致。这一现象以前未见相关报道, 具体原因有待以后进一步深入研究。在实际应用中注意草甘膦的施用浓度, 将有助于防止或缓解药害的发生。
采用过量表达epsps或引入突变的epsps使植物获得对草甘膦耐性方法易造成植物体内草甘膦的累积, 使植物出现败育、授粉率降低、不耐胁迫、农艺性状改变等问题[19]。本研究结果再次证实尽管转epsps基因可以获得高抗草甘膦植株, 但在花期喷施草甘膦将造成败育, 这就要求在草甘膦的实际使用中注意作物适用生育期。
研究结果显示高抗转基因植株是由于转入基因的转录水平(或表达量)高; 而从分离比看这又不是因为多拷贝引起; 更主要的原因应该是由不同的插入位点所致。最近的研究发现由于草甘膦的多年使用, 在选择压下自然界中的杂草有的对草甘膦产生了自然抗性。美国佐治亚州的抗草甘膦长芒苋(Amaranthus palmeri)中含有5~160倍拷贝数的epsps基因, 且分布于每条染色体[20], 表明多拷贝超剂量的表达抗性基因可增加植物的草甘膦抗性。虽然多拷贝的转入外源基因有可能造成转入的基因沉默和抑制, 但在适当的情况下这未尝不是一种可用的手段。此外, 将epsps基因和草甘膦降解基因联合使用将有可能做到优势互补。
本研究只是利用十字花科模式植物进行的验证。由于不同的物种自身对草甘膦的抗性表现不同, 在单子叶大田作物中抗性基因的抗性和表现可能会有差异。在目前的大田作物应用中cp4-epsps基因仍是最主要的草甘膦抗性基因[3]。从基因大小和抗性强弱考虑, GAT类基因作为筛选标记基因应该更为适合, 但在具体实施中应注意草甘膦的施用浓度。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [3] |
|
| [7] |
|
| [9] |
|
| [10] |
|
| [15] |
|
| [17] |
|