


{kind=link}
{kind=link}
{kind=link}
{kind=link}
中国伞形科前胡属果实结构的系统学价值
[王淼媛 , 刘玫
, 刘玫* , 程薪宇, 王宇婷]
, 刘玫, 程薪宇, 王宇婷]
|
|


作者简介:王淼媛(1983-),女,黑龙江哈尔滨人,在读博士。E-mail: m.y.wang@163.com
本文深入研究了位于中国前胡属刺尖前胡组,多小苞片组,山亮蛇床组及近亮蛇床组的果实结构,结果显示前胡属果实多具单细胞毛状体。中脊及侧脊通常不明显,边脊形成翅。多数物种具小的伴生油管。油管分布在沟(1~4个)及结合面(2~14个),一些物种具短的或小的油管。结合面与分果同宽,具两个相对的心皮柄。胚乳腹面多平直。中果皮有球状晶体。前胡属不同组的果实结构相似,此属应是一个较自然的类群,属内是否需要分组值得进一步研究。石防风的果实与其他物种不同,具厚的边翅和大的维管束,支持分子系统学将其从前胡属分离出去。
The fruit structures of the sections Eleganitia, Bracteolata, Oreoselinum, and Selinoides in the genus Peucedanum were deeply studied in the paper. The results show that fruits of the genus usually have single cellular trichome. The median and lateral ribs are usually not prominent, but marginal ribs form wings. The small rib duct occurs in many taxa. Vittae are present in the furrow (1-4) and commisure (2-14), and short and drowf vittae are observed in some species. The commisure is as wide as the mericarp width and with two opposite carpophores. The endosperm is often flat at the commissural side. The spherical crystals occur in the mesocarp. The fruit structures are similar in different sections and the Peucedanum should be a more natural group, and the further study is needed to know if the genus should be divided into sections. The fruits of Peucedanum terebinthaceum differ from the other taxa, which have thick marginal wings and large vascular bundles, and it is supported here that the molecular research moved the Peucedanum terebinthaceum out from the genus.
伞形科(Apiaceae)前胡属(Peucedanum)位于芹亚科(Apioideae)前胡族(Peucedaneae)[1], 全世界100~120种, 主要分布于欧洲, 亚洲, 非洲及北美洲[2], 我国约40种, 其中33种为特有, 广泛分布[3]。花粉形态学揭示这一属花粉为三棱长球形, 三孔沟, 边孔, 是一个较自然和进化的类群[4, 5, 6]。分子系统学研究显示前胡属与当归属等类群共同位于当归分支(Angelica clade)[7]。前胡属的石防风(Peucedanum terebinthaceum)是棱子芹族(Pleurospermeae)喜马拉雅棱子芹(Pleurospermum hookeri)的姊妹群[8, 9]。单人烨和佘孟兰根据萼片, 总苞及果实的特征将中国前胡属分为5个组:刺尖前胡组(Sect. Elegantia), 多小苞片组(Sect. Bracteolata), 山亮蛇床组(Sect. Oreoselinum), 近亮蛇床组(Sect. Selinoides)及前胡组(Sect. Peucedanum)[10]。Zhou等[11]指出前胡属的每个物种有其特有的叶表皮特征, 传统的分组可能是不自然的, 石防风与其他物种不同, 其保卫细胞略圆形, 表皮细胞垂周壁波浪状, 建议将其从前胡属分离出来。
伞形科的双悬果是高度特化的器官, 具有丰富的变异和结构稳定的特点, 是这一科分类的重要依据, 其果皮、翅及结晶等特征被用于亚科, 族及属的分类[1, 12]。近年来随着这一科果实研究的深入, 进一步揭示了果实微形态(如油管、微管束、结合面及心皮柄等)的重要性, 为分子系统学的研究提供了重要的形态学依据[13, 14, 15, 16]。已有的关于前胡属果实结构的研究侧重的是表面特征[17]。本文的目的是通过进一步深入研究我国前胡属果实微结构, 并与已研究的相关类群相比较, 揭示这一属是否为自然类群, 为分子系统学的研究提供形态学依据, 并为进一步研究世界前胡属及当归分支提供形态学基础。
前胡属19种及2变种果实, 物种名称及凭证标本信息见表1。材料分别取自于中国科学院成都生物研究所标本馆(CDBI-Herbarium, Chengdu Institute of Biology, Chinese Academy of Sciences), 中国科学院华南植物园标本馆(IBSC-South China Botanical Garden Herbarium), 中国科学院沈阳应用生态研究所标本馆(IFP-Institute of Applying Ecology, Chinese Academy of Sciences), 中国科学院昆明植物研究所标本馆(KUN-Herbarium, Kunming Institute of Botany, Chinese Academy of Sciences), 江苏省中国科学院植物研究所标本馆(NAS-Herbarium, Institute of Botany, Jiangsu Provence, Chinese Academy of Science)及中国科学院植物研究所标本馆(PE-Herbarium, Institute of Botany, Chinese Academy of Sciences)。这些材料覆盖了前胡属的4个组(刺尖前胡组, 多小苞片组, 山亮蛇床组和近亮蛇床组)。
| 表1 用于研究果实特征的中国前胡属物种的凭证标本信息及其物种的采集地 Table 1 Taxa of Peucedanum in China examined for fruit features, together with voucher specimen details and locality |
观察毛状体及油管:将干的果实放入热水(约90℃)中浸泡, 待材料完全吸水膨胀并沉入水底时(约为2 h), 将其取出, 用解剖镜(Olympus SP-350)观察油管的分布及结构并用数码相机(Olympus DP 70)照相。之后, 撕取外果皮, 将其放在载玻片上并加1~2滴50%甘油水溶液, 用显微镜(Olympus BX 51)观察毛状体结构。
GMA(glycolmethacrylate:乙二醇甲基丙烯酸酯)切片:根据Feder和O’ Brien[18]的方法, 将经吸水膨胀的果实(每个物种取2个)放入FAA(formalin-acetic acid-alcohol: 福尔马林-冰醋酸-酒精)固定, 时间不少于24 h。材料经50%酒精(4~6 h), 100%酒精(4~6 h), 异丙醇(6 h)和正丁醇(6 h)脱水, 每步各重复1次。接着材料进入GMA渗透, 共3次, 第1和2次各为1 d, 第3次不少于5 d。之后将材料及GMA装入无色胶囊, 并将其置于60℃温箱聚合(24 h)。用Leica Ultracut R 切片机切片, 厚度2~3 μ m。同样根据Feder和O’ Brien[18]的方法, 将切片用甲苯胺蓝染色, 中性树胶封片[18]。用具数码相机(Olympus DP 26)的显微镜(Olympus BX 51)观察果实结构并照相。
本文详细地描述了中国前胡属19种及2变种果实结构, 包括毛状体, 果皮, 脊, 翅, 维管束, 油管, 结合面, 胚乳, 心皮柄及结晶, 详细特征见图1~图4, 主要分类特征见表2。
毛状体:前胡属一些物种果实表面具单细胞毛状体, 如刺尖前胡(刺尖前胡组), 滨海前胡(多小苞片组), 毛前胡(山亮蛇床组), 华北前胡(近亮蛇床组), 其中华北前胡的果皮还偶见2细胞毛状体(图3A1, 2)。有些物种的果皮光滑, 无毛状体, 如会泽前胡(刺尖前胡组), 竹节前胡(多小苞片组), 北京前胡(山亮蛇床组), 南川前胡和石防风(近亮蛇床组)(图3C1, 2)。脊及翅:果实均为背腹压扁(图3A1~C1, 图4A~U), 分果的边脊(即靠近结合面的脊)形成翅, 即边翅(图1A~S, 图3A1~C1, 图4A~U)。石防风和宽叶石防风的边翅厚实, 达分果厚度的1/2(图3C1, 图4R, 图4S), 其他物种的边翅较薄, 如华北前胡(图3A1, 图4H), 会泽前胡(图4A), 滨海前胡(图4J)及毛前胡(图3B1, 图4O)。多数物种果实的中脊及侧脊无明显向外突起, 如会泽前胡, 红前胡(山亮蛇床组)及兴安前胡(近亮蛇床组)(图4A, 图4B, 图4P)。少数物种果实的中脊及侧脊明显向外突起, 即脊处果皮的厚度为沟处的1.5~2.0倍, 包括南岭前胡(多小苞片组)(图4K), 华北前胡(图4H), 细裂前胡(山亮蛇床组)(图4L)及毛前胡(图4O)。边翅横切面多平直, 如华北前胡及毛前胡(图3A1, 图3B1), 但石防风及宽叶石防风边翅呈弧形(图3C1, 图4R, 图4S)。
维管束:存在于分果的5个脊(即中脊, 侧脊及边脊)中(图4B), 多数物种每个脊内具1个维管束, 如会泽前胡(图4A), 南岭前胡(图4K), 华中前胡(近亮蛇床组)(图4M), 泰山前胡(山亮蛇床组)(图4U)。但少数物种的在边脊(或边翅)中可见2个维管束, 如北京前胡(图4C), 南川前胡(图4F), 草原前胡(多小苞片组)(图4Q)。维管束通常较小, 约占果皮厚度的1/2, 如会泽前胡(图4A), 但石防风边翅的维管束明显较大, 约为中脊及侧脊中维管束的5倍(图4R)。
伴生油管:伴生油管为与维管束相伴的油管, 存在于分果的脊(或翅)内维管束的外侧(图3B2, 图4B)。多数前胡物种具伴生油管, 如滇西前胡(图4D), 少毛前胡(近亮蛇床组)(图4I), 毛前胡(图3C2, 图4O)及前胡(多小苞片组)(图4N)。多数伴生油管的官腔小(如图4A, 图4C), 有时不易观察到。少数物种无伴生油管, 他们分别位于刺尖前胡组等4个组, 如竹节前胡(图4E), 刺尖前胡(图4G), 细裂前胡(图4L), 华中前胡(图4M)。
非伴生油管:非伴生油管简称油管, 不与维管束相伴, 存在于在果实的沟里, 即两个脊之间的中果皮(图1A~S)及结合面(图2A~S)。少数物种果实的沟里油管为1个, 如兴安前胡(图1B), 刺尖前胡(图1G), 北京前胡(图4C), 兴安前胡(图1B)。有些物种沟里的油管多达4个, 如滇西前胡(图4D), 华中前胡(图4M), 前胡(图4N), 毛前胡(图4O)。结合面油管在少数物种为2个, 如兴安前胡(图2B), 刺尖前胡(图2G), 草原前胡(图2P), 石防风(图2Q)。而多数物种结合面油管数目较多, 可达10~14个, 滇西前胡(图2D), 滨海前胡(图2I)及华中前胡(图2L)。一些物种具短油管或小油管, 如滇西前胡(图2D), 南岭前胡(图2J), 长前胡(图2R)。
 | 图1 前胡属分果外形, 示中脊, 侧脊, 边翅, 及沟里的油管Fig.1 The mericarps shapes of Peucedanum showing median rib, lateral rib, marginal wing and vitta |
 | 图2 前胡属分果结合面, 示短油管及小油管Fig.2 The commisure sides of mericarps of Peucedanum showing regular and irregular vittae |
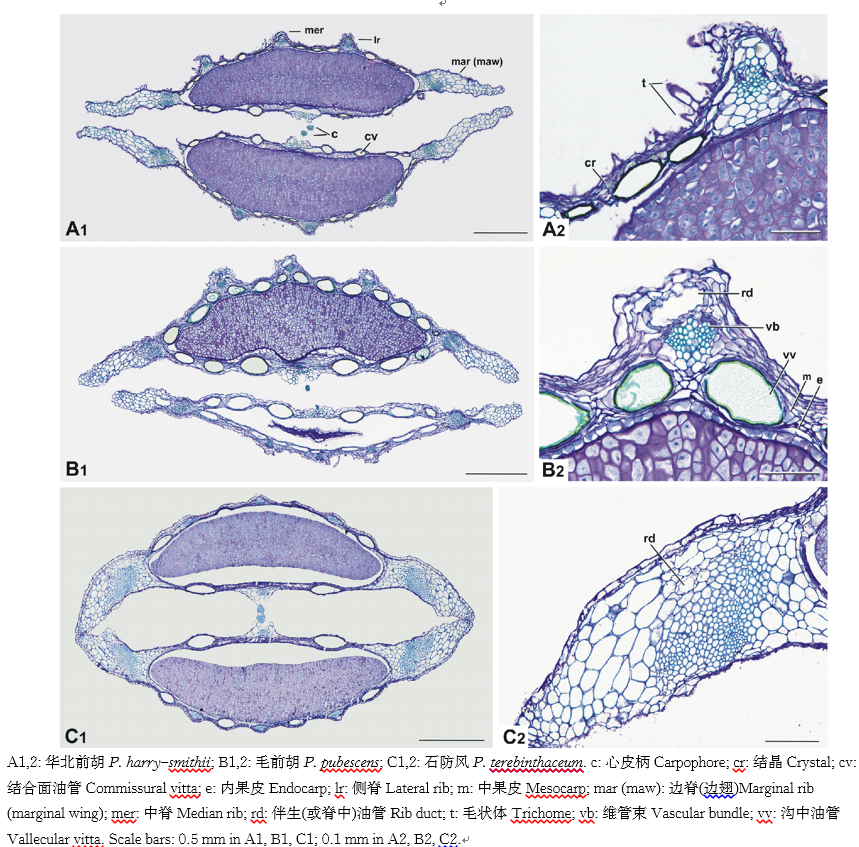 | 图3 前胡属果实横切面, 示分果形状, 毛状体, 脊, 翅, 维管束, 伴生油管, 沟里及结合面油管, 胚乳, 结合面宽及心皮柄特征Fig.3 Transverse sections of fruits of Peucedanum showing variation in mericarp shape, trichome, ribs, marginal wing, vascular bundle, rib duct, vittae in furrow and commisure, endosperm, commissural width and features of the carpophore |
中果皮, 内果皮及胚乳:中果皮由6~10层细胞构成, 围绕种子的中果皮通常为薄壁细胞, 但翅中的细胞轻微木化(图3B2)。内果皮通常为1层薄壁细胞(图3B2)。胚乳在结合面多为平直, 如会泽前胡(图4A), 兴安前胡(图4B), 滨海前胡(图4J)。但北京前胡(图4C), 竹节前胡(图4E)及南岭前胡(图4K)的胚乳微凸。
结合面及心皮柄:结合面为2分果的结合处(图4B)。前胡属所有物种均具有非常宽的结合面, 其宽度同分果的宽度相同(图4A~U)。两个较细的心皮柄彼此相对, 存在于所有研究的物种(图3A1~C1, 4A~U)。
结晶:球状的晶体存在于前胡属多数物种的外果皮及中果皮, 如会泽前胡, 竹节前胡, 北京前胡, 华北前胡(图3A2), 有些物种未见球晶, 如毛前胡(图3B2)。
 | 图4 前胡属果实横切面, 示分果形状, 脊, 边翅, 维管束, 伴生油管, 沟里及结合面油管, 胚乳, 结合面宽及心皮柄特征Fig.4 Transverse sections of the fruits of Peucedanum showing variation in mericarp shape, rib, marginal wing, vascular bundle, rib duct, vittae in furrow and commisure, endosperm, commissural width and features of the carpophore |
本研究涉及单人烨和佘孟兰前胡属分类系统中5个组的4个组[10], 即刺尖前胡组, 多小苞片组, 山亮蛇床组和近亮蛇床组。果实结构在不同组内的变化以及物种之间的关系讨论如下。
| 表2 中国前胡属果实的重要分类特征 Table 2 Summary of taxonomically important characters of the fruits in various taxa of Peucedanum of China |
刺尖前胡组:刺尖前胡组叶末回裂片线形, 顶端刺尖状[10]。这个组包括4种, 本文研究了3种。惠红等[19]根据会泽前胡植物极其矮化和地理分布特点, 建议将其从刺尖前胡组独立出来, 并拟与木里前胡(Peucedanum muliense)、矮前胡(Peucedanum nanum)和滇西前胡等西南高山矮化种类成立一新组。Pimenov[20]将刺尖前胡移入Kitagawa, 改名为K. komarovii。然而研究显示会泽前胡与滇西前胡的果实结构非常相似, 均具伴生油管, 沟里和结合面的油管均为多个(图4A, D)。尽管刺尖前胡的油管数量较少, 沟里1及结合面2(图4G), 但此种同会泽前胡一样, 中果皮具结晶。沟里和结合面油管数量少(分别具1和2)的特征存也在于其他组的物种, 如兴安前胡(图2B, 图4B)和草原前胡(图2P, 图4Q)。Kitagawia baicalensis的种皮由3~5层细胞构成(未发表), 这一特征不存在于前胡属所有研究的物种。
多小苞片组:多小苞片组叶末回裂片顶端不呈刺尖状[10]。这个组包括10种, 本文研究了6种。滨海前胡的茎曲折, 常呈蜿蜒状, 生长于滨海滩地或近海山地[10]。刘启新和惠红[21]指出滨海前胡与该组成员不同, 可以认为是一个在特定区域和生态环境下独立演化而成的类群, 在分类上似应把它从原来的组中分出, 单独成一组。然而果实结构表明滨海前胡结合面油管与滇西前胡的油管均具多条油管(图2I, 2D)。此外, 滨海前胡的毛状体及结晶等特征与同组的其他物种相同(如前胡)。Zhou等[11]的叶表皮结构也显示滨海前胡很难与本组的其他物种区分。
山亮蛇床组:山亮蛇床组果实侧棱狭窄而厚[10]。这个组包括6种, 本文研究了5种。刘启新和惠红[21]指出红前胡和刺尖前胡有相似的生态环境和形态特征, 而且酶谱特征亦相似。本研究显示红前胡(图4P)及泰山前胡的边翅较窄(图4U)。然而同组的其他物种, 如北京前胡(图4C)及细裂前胡(图4L)具有较宽的边翅, 同位于其他组的刺尖前胡(图4G)及南川前胡(图4F)的边翅也较宽。而且山亮蛇床组果实的其他特征(如维管束, 伴生油管, 油管及结晶等)也存在于其他组的物种。
近亮蛇床组:近亮蛇床组果实侧棱较宽[10]。这个组有10种, 本文研究了5种。石防风被转移到Kitagawia, 作为这个属的模式种, 即K. terebinthacea[20]。血清分类学研究表明石防风应自成一类, 可建一新组[22]。石防风与前胡属不同这一结论曾得到分子系统学及叶表皮结构[11]的证实[9], 刘启新等[22]指出宽叶石防风是从石防风演化而来, 本研究显示这两个物种果实边翅厚, 背面呈弧状(图4R, S), 而其他物种边翅薄而且背面平直(图4A~Q, T, U), 而且石防风翅中维管束明显较大(图4R)。这些特点均说明石防风及宽叶石防风应从前胡属分出来。
果实结构显示前胡属是一个比较自然的类群, 不同组物种果实结构相似, 因而属下分组可能是不合适的。石防风及宽叶石防风果实不同于前胡属其他物种, 支持分子系统学将此种从前胡属分离出去[9]。此外前胡属果实的毛状体、边翅、中果皮内具结晶、内果皮为薄壁细胞构成等特征同当归属的果实相似, 为分子系统学揭示的二者有较近的亲缘关系提供了形态学依据[7]。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [10] |
|
| [17] |
|
| [19] |
|
| [21] |
|
| [22] |
|