

{kind=link}
{kind=link}
{kind=link}
{kind=link}
青藏高原高寒草甸花期物候和群落结构对氮、磷、钾添加的短期响应
[杨月娟1, 2, * , 张灏3, ** , 周华坤1, *  , 叶鑫
, 叶鑫2, 4 , 姚步青1 , 张春辉1 , 马真1 , 赵新全1 ]
, 叶鑫|
|


作者简介:杨月娟(1988-),女,山东青岛人,在读硕士。E-mail:yangyuejuan198907@163.com。张灏(1988-),男,山东潍坊人,在读硕士。E-mail:582545295@qq.com。
在中国科学院海北高寒草甸生态系统定位站的矮嵩草草甸水肥样地进行了氮、磷、钾施肥添加试验,研究了施肥对植物花期物候和群落结构的影响。结果表明,施肥使矮嵩草草甸有效积温增加,地下5 cm和地上10 cm有效积温的大小均为:氮>磷>钾>对照;施肥使垂穗披碱草、甘青剪股颖、冷地早熟禾、矮嵩草、黑褐薹草、双柱头藨草、美丽风毛菊的始花期、终花期显著推迟( P<0.05),而鹅绒委陵菜和麻花艽的始花期、终花期显著提前( P<0.05);施肥后禾草的花期持续期小于莎草和杂类草;与对照相比,垂穗披碱草-麻花艽、垂穗披碱草-美丽风毛菊、甘青剪股颖-麻花艽和冷地早熟禾-麻花艽的开花时间(始花期)上的差异变小,种间开花时间重叠增加;非度量多维尺度分析的检验结果为,对照、钾、磷和氮处理下其群落之间的相似程度逐渐降低,添加氮、磷促进了非相似性群落的形成。熵值法综合评价分析得出,对矮嵩草草甸添加氮、磷后,花期物候响应较为敏感的物种为:垂穗披碱草、矮嵩草、麻花艽和美丽风毛菊。
, YE XinFertilizers (N, P & K) were applied to a Kobresia humilis meadow at the Haibei Alpine Meadow Ecosystem Research Station to determine the effects of fertilizer on plant flowering phenology and community structure. Fertilizer application increased the effective accumulated soil temperature at 5 and 10 cm depth: nitrogen>phosphorus>potassium>control treatment. The first floweing day and end flowering day was delayed in a number of plant species including Elymus nutans, Agrostis hugoniana, Poa crymophila, Kobresia humilis, Carex atrofusca, Scirpus distigmaticus and Saussurea superb ( P<0.05), whereas in Potentilla anserine and Gentiana straminea flowering was advanced ( P<0.05). Flowering duration in Gramineae was lower than in Cyperaceae and Forbs following fertilizer application. The application of nitrogen and phosphorus promoted the formation of a heterogeneous community. Heterogeneity, assessed using nonmetric multidimensional scaling, was lowest in control fertilizer treatments followed by potassium, phosphorus, and finally nitrogen application.It was concluded that E. nutans, K. humilis, G. straminea and S. superb were sensitive to the application of nitrogen and phosphorus in K. humilis meadow.
植物的开花时间是植物生活史中最重要的阶段, 花期紧密地连接着营养生长和生殖生长, 植物花期的变化会影响到植物的表现以及适合度进而影响到群落结构和功能[1, 2]。草地施肥是维持草地生态系统养分平衡的有效措施[3], 是目前加速退化草地恢复重建的关键技术措施之一[4]。合理的草地施肥可以提高牧草的生物量和品质, 有助于草地生产力的恢复和提高[5, 6]。尽管已有研究表明, 环境变化会对植物物候产生较大影响[7, 8], 且有关环境变化对植物物候期影响的研究较为普遍[9, 10, 11], 但目前关于环境变化对花期物候的影响尚无明确定论[12, 13, 14]。近年来的研究表明, 土壤养分的增加会引起植物物候的变化[15], 草地群落研究中却很少涉及施肥对草地群落花期物候的影响。Cleland等[15]在草地群落的试验表明, 施氮使杂类草开花期轻度提前, 而禾草开花期显著推迟, 即增加土壤肥力, 禾草营养生长期显著延长而繁殖生长期相应推迟。李元恒[16]在内蒙古草原的研究表明, 单施氮肥或磷肥使禾草开花期显著提前, 但对其生殖生长持续期无显著影响, 同时施氮肥和磷肥使禾草开花期显著提前, 生殖生长持续期延长。高寒草甸由于其特殊地理和气候条件, 生态系统脆弱, 植物物候对环境因子的变化较为敏感, 研究高寒地区植物物候对环境变化的响应, 可以预测其他区域植物物候的变化情况, 具有重要的理论和实践意义。
本研究通过对围栏封育的矮嵩草(Kobresia humilis)草甸采用不同的施肥处理, 研究添加土壤速效养分对高寒草甸花期物候(始花期、终花期、花期持续期)和群落结构的影响, 拟回答3个问题:1)矮嵩草草甸植物花期物候对N、P、K添加的响应特征是什么?2)土壤养分环境变化对群落结构的影响程度如何?3)矮嵩草草甸植物群落有哪些植物种对土壤速效养分添加响应较为敏感?
试验选择在中国科学院海北高寒草地生态系统国家野外科学观测研究站(简称海北站)附近区域进行。海北站地理坐标N 37° 37', E 101° 19'。地处青藏高原东北隅, 祁连山东段北支冷龙岭南麓, 大通河河谷的西北部, 位于青海省海北藏族自治州门源回族自治县境内的风匣口。该地区海拔3200~3600 m, 站区以低山、丘陵、滩地和河流阶地为主, 受高海拔条件的制约, 气候具有明显的高原大陆性特点, 无四季之分, 仅有冷暖二季之别, 干湿季分明, 年温差较小而日温差较大, 太阳辐射强烈[17]。青藏高原隆起过程所形成的特殊自然环境, 造就了适应寒冷湿中生的多年生草本植物种类, 定位站地区约有种子植物500余种, 其中以禾本科、菊科、龙胆科和毛茛科最多, 其次为莎草科、蔷薇科和豆科, 其中高寒草甸的优势种群为莎草科的嵩草属[18]。矮嵩草草甸是该地区主要草场类型之一, 禾本科牧草在此植物群落组成和生物量分配中的比例较大, 在高原草地畜牧业生产中具有重要的地位, 具有高蛋白、高脂肪、高无氮浸出物、高热值含量与低纤维等四高一低的特点, 是发展草地畜牧业的物质基础[19]。土壤特征主要表现为发育较年轻, 土层薄, pH值多在7以上, 土壤库内N、P、K储量丰富, 土壤潜在肥力高, 但营养物的矿化过程微弱, 所以养分有效率低, 尤其是速效性N、P含量[20]。
在围栏样地中按单因素设计试验, 试验采用国际养分网络(nutrient network)的施肥处理设计[21], 3个养分[氮(N)、磷(P)、钾(K)]添加处理, 共4个处理[N、P、K、CK(对照)], 每个处理重复6次, 共24个小区。每个小
区面积为6 m× 6 m, 各小区间有2 m 的缓冲带, 缓冲带不施肥, 各区的四角用木桩作标记。施肥时间选择在生长期中期的一个阴雨天或晴天的傍晚进行, 雨水或次日形成的露水使得肥料融化, 以保证植物对肥料的有效利用。施肥的日期分别为2009年7月15日、2010年7月5日, 施肥的方法是用手工均匀地撒在地表[22]。施肥量按照国际养分网络(nutrient network)上认可的标准进行配比[23], 每年的施肥量经折算后, 见表1。
| 表1 试验小区化肥施加量 Table 1 The amount of fertilizer applied at plot |
1.3.1 测定方法
在2010年, 选用HOBO温度自动记录仪对生长季各月的温度进行记录, 选用两个层次:地下5 cm和地上10 cm, 每两小时自动记录一次。物候观测从群落返青开始到群落中多数植物种凋落时结束。由于选择的植物包括禾本科、莎草科和杂类草, 为了便于统计和比较其物候期, 将禾本科植物的拔节、抽穗、开花期合并为开花期, 即物候的选择包括:返青营养期、花蕾开花期、结实期、果后营养期、枯黄期。实验采用定株标记的方法进行物候观测, 标记植物主要为矮嵩草草甸的优势种和常见种, 具有一定的代表性。其中, 禾本科植物3种, 分别是垂穗披碱草(Elymus nutans)、甘青剪股颖(Agrostis hugoniana)、冷地早熟禾(Poa crymophila); 莎草科植物3种, 分别是矮嵩草、黑褐薹草(Carex atrofusca)、双柱头藨草(Scirpus distigmaticus); 杂类草3种, 分别是麻花艽(Gentiana straminea)、美丽风毛菊(Saussurea superba)、鹅绒委陵菜(Potentilla anserina)。每个处理中的同种标记个数大于12株或3~5丛。对每个种选定个体时考虑种内个体差异、生境差异和分布位置差异, 以避免在同一个样方或相邻样方中集中定株。
2010年完成21轮物候观测, 每轮物候观测开始日期(月.日):4.27、5.3、5.8、5.15、5.22、5.30、6.7、6.15、6.21、6.30、7.8、7.17、7.25、7.30、8.7、8.15、8.21、8.28、9.10、9.16、9.27。第1次观察到物种某一物候期的出现日期记为该种该物候期的开始日, 而把最后1次观测到的日期记为该物候期的结束日, 某一物候期的持续时间为从开始日到结束日之间的间隔期[24]。
1.3.2 计算方法
根据逐日平均气温, 采用五日滑动平均法计算有效积温[25]。用儒略历(Julian calendar)计数法(1月1日为1年中第1天)进行物候期统计分析。N、P、K施肥处理下, 矮嵩草草甸植物种始花期(first flowering date, FFD)变化和终花期(end flowering date, EFD)变化分别表示为:FFDX-FFDCK和EFDX-EFDCK, FFDX和EFDX分别表示某一物种在N、P、K添加处理中的始花期和终花期, FFDCK和EFDCK分别表示某一物种在未施肥中的始花期和终花期, 差值为正值表示物种的始(终)花期推迟, 差值为负值表示始(终)花期提前。不同施肥处理下矮嵩草草甸禾草和杂类草开花时间差异表示为:|FFDG-FFDF|, FFDG和FFDF分别表示在CK、N、P、K处理下禾草和杂类草的始花期。
采用SAS 9.2分别对矮嵩草草甸植物始花期、终花期进行单因素四水平设计一元定量资料差异性检验。采用SAS 9.2实现非度量多维尺度分析(nonmetric multidimensional scaling, NMDS)检验土壤养分添加对花期物候加权的群落结构相似性的影响, 采用熵值法(entropy method)综合评价分析植物群落中对土壤养分添加响应较为敏感的物种。
从表2可以看出, 施肥使矮嵩草草甸≥ 10℃有效积温(以下简称有效积温)增加, 地下5 cm和地上10 cm有效积温的大小均为:N> P> K> CK。与CK相比, 地下5 cm时, N施肥处理有效积温增加的幅度最大(24.35%), K施肥处理有效积温增加的幅度最小(1.54%), 而地上10 cm时, 亦N施肥处理有效积温增加的幅度最大(20.36%), K施肥处理有效积温增加的幅度最小(2.81%)。
| 表2 不同施肥处理下矮嵩草草甸的有效积温 Table 2 Effective temperature of K. humilis meadow under different fertilizing treatments ℃ |
2.2.1 不同施肥处理对矮嵩草草甸始花期的影响
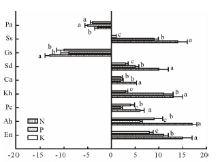
由表3和图1可以看出, 施肥使垂穗披碱草、甘青剪股颖、冷地早熟禾、矮嵩草、黑褐薹草、双柱头藨草、美丽风毛菊始花期显著推迟(P< 0.05), 而鹅绒委陵菜和麻花艽的始花期显著提前(P< 0.05)。在N、P、K添加处理下, 垂穗披碱草、甘青剪股颖、冷地早熟禾始花期比对照分别推迟15, 11, 8 d; 17, 6, 9 d; 6, 2, 4 d, 矮嵩草、黑褐薹草、双柱头藨草始花期分别推迟13, 11, 3 d; 5, 2, 2 d; 10, 5, 3 d, 而杂草类始花期对养分添加的响应不一致, 美丽风毛菊始花期推迟14, 9, 1 d, 麻花艽和鹅绒委陵菜始花期分别提前13, 9, 10 d; 3, 5, 4 d。在N添加处理下, 禾草(垂穗披碱草、甘青剪股颖和冷地早熟禾)和双柱头藨草始花期与P、K均有显著差异, 而美丽风毛菊N、P、K处理之间的始花期均有显著差异, 鹅绒委陵菜N、P、K处理之间的始花期均未达显著差异水平。
| 表3 不同施肥处理对矮嵩草草甸始花期的影响 Table 3 Effects of different fertilization treatments on first flowering day of K. humilis meadow d |
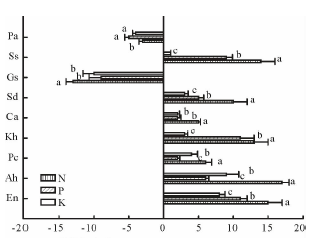 | 图1 不同施肥处理下矮嵩草草甸始花期的变化Fig.1 Changes of first flowering day of K. humilismeadow in different fertilization treatments En:垂穗披碱草E. nutans; Ah:甘青剪股颖A. hugoniana; Pc:冷地早熟禾P. crymophila; Kh: 矮蒿草 K. humilis; Ca:黑褐薹草 C. atrofusca; Sd:双柱头藨草 S. distigmaticus; Gs:麻花艽G. straminea; Ss:美丽风毛菊S. superba; Pa:鹅绒委陵菜P. anserine.不同小写字母表示同一植物不同处理间差异显著(P< 0.05)。 Different lowercase letters indicate significant difference at 0.05 level within different treatments of the same plant. 下同。The same below. |
2.2.2 不同施肥处理对矮嵩草草甸终花期的影响
由表4和图2可以看出, 施肥使垂穗披碱草、甘青剪股颖、冷地早熟禾、矮嵩草、黑褐薹草、双柱头藨草、美丽风毛菊终花期显著推迟(P< 0.05), 而鹅绒委陵菜和麻花艽的终花期显著提前(P< 0.05)。在N、P、K添加处理下, 垂穗披碱草、甘青剪股颖、冷地早熟禾终花期比对照分别推迟21, 13, 4 d; 5, 3, 2 d; 5, 2, 3 d, 矮嵩草、黑褐薹草、双柱头藨草终花期分别推迟19, 12, 4 d; 11, 4, 1 d; 7, 2, 4 d, 美丽风毛菊终花期推迟20, 11, 6 d, 麻花艽和鹅绒委陵菜终花期分别提前13, 8, 2 d; 8, 11, 10 d。在N添加处理下, 黑褐薹草终花期与P、K有显著差异, 而垂穗披碱草、矮嵩草、麻花艽和美丽风毛菊N、P、K处理之间的终花期均有显著差异, 鹅绒委陵菜N、P、K处理之间的终花期均未达显著差异水平。
| 表4 不同施肥处理对矮嵩草草甸终花期的影响 Table 4 Effects of different fertilization treatments on end flowering day of K. humilis meadow d |
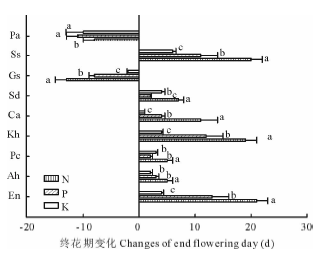 | 图2 不同施肥处理下矮嵩草草甸终花期的变化Fig.2 Changes of end flowering day of K. humilis meadow in different fertilization treatments |
2.2.3 不同施肥处理下矮嵩草草甸植物的花期持续期
由表5可得, 矮嵩草、黑褐薹草、美丽风毛菊在N、P、K添加下的花期持续期均大于CK, 分别增加了6, 1, 1 d; 6, 2, 1 d; 6, 2, 5 d; 甘青剪股颖、鹅绒委陵菜在N、P、K添加下的花期持续期均小于对照, 分别减少了12, 3, 9 d; 5, 6, 6 d; 而冷地早熟禾在P添加处理下的花期持续期等于CK, 在N、K中减少; 麻花艽在N添加处理下的花期持续期等于CK, 在P、K中增加; 双柱头藨草的花期持续期在N、P中减少, 在K中增加。总体上相比较而言, 施肥后禾草花期持续期小于莎草和杂类草的花期持续期。
| 表5 不同施肥处理下矮嵩草草甸植物的花期持续期 Table 5 Flowering lasting days of K. humilis meadow under different fertilizer treatments d |
2.2.4 不同施肥处理对矮嵩草草甸禾草和杂类草开花时间差异的影响
自然群落中该研究区域的禾本科类植物(垂穗披碱草、甘青剪股颖和冷地早熟禾)的开花时间明显地早于杂类草(麻花艽、美丽风毛菊、鹅绒委陵菜)的开花时间, 但是在N、P、K添加处理下(图3), 垂穗披碱草-麻花艽、垂穗披碱草-美丽凤毛菊、甘青剪股颖-麻花艽和冷地早熟禾-麻花艽的开花时间(始花期)上的差异均显著小于各自CK(P< 0.05), 增加了开花时间上的重叠, 潜在的增加了群落中植物对资源的竞争, 可能会引起植物在繁殖及相互竞争、共存上的新格局。
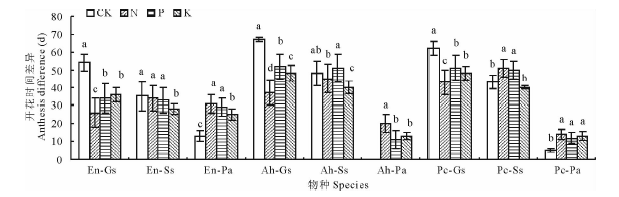 | 图3 不同施肥处理对矮嵩草草甸禾草和杂类草开花时间差异的影响Fig.3 Effects of different fertilization treatments on anthesis difference of Gramineae and Forbs in K. humilis meadow |
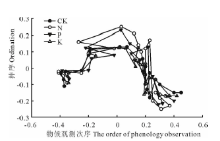
非度量多维尺度分析排序检验了CK、N、P、K添加对群落结构的影响(图4), 4条不同的曲线分别表示了非度量多维尺度分析排序呈现出的4个养分添加处理(CK、N、P、K)下群落之间在花期的群落结构相似性分离程度, 应力=0.039, 说明非度量多维尺度分析拟合矮嵩草草甸群落结构相似性较好。结果表明, 在不同养分添加下, 群落结构有显著差异, CK、K、P和N群落之间的相似程度逐渐降低, CK群落与K群落的相似程度大于P群落、N群落, 群落每次物候观测之间的相似性也变低。这说明N、P添加后矮嵩草草甸群落的物候和群落物种组成的变化较为剧烈, 群落物候偏移, 促进非相似性群落的形成。
 | 图4 矮嵩草草甸群落结构相似性在N、P、K添加下的非度量多维尺度分析排序(应力=0.039)Fig.4 Nonmetric multidimensional scaling ordination ofK. humilis meadow community similarity among nitrogen, phosphorus and potassium addition gradient(stress value=0.039) 同一曲线上从左至右的21个点表示群落内物候观测次序, 从2010年4月27日— 2010年9月27日共21次观测。21 points from left to right of the same curve indicate the order of phenology observation from April 27th to September 27th of 2010, 21 observation times in all. |
由于K在青藏高原高寒地区并不是限制因子, 所以仅筛选出对添加N、P响应较为敏感的植物种, 研究植物物候对环境条件变化的响应特征, 为以后对其作进一步研究提供依据。采用熵值法确定各指标(始花期、终花期和花期持续期)的权重, 综合评价分析对N、P添加敏感的物种。物候作为一种物种性状反映了植物生活史对策差异, 矮嵩草草甸植物花期物候响应N肥的敏感性大小分别为:垂穗披碱草> 美丽风毛菊> 矮嵩草> 麻花艽> 甘青剪股颖> 鹅绒委陵菜> 冷地早熟禾> 双柱头藨草> 黑褐薹草, 矮嵩草草甸植物花期物候响应P肥的敏感性大小分别为:矮嵩草> 麻花艽> 美丽风毛菊> 垂穗披碱草> 冷地早熟禾> 甘青剪股颖> 鹅绒委陵菜> 黑褐薹草> 双柱头藨草(表6)。所以, 对N、P施肥处理下矮嵩草草甸群落花期物候响应较为敏感的物种为:垂穗披碱草、矮嵩草、麻花艽和美丽风毛菊。
温度是影响植物物候的最重要因素, 植物的各个生长发育期都有一定的积温要求, 达到了一定的积温要求后, 物候现象才会出现[26]。温度是高寒地区植物生长的限制性因子, 地下5 cm和地上10 cm有效积温的大小均为:N> P> K> CK(表2)。这主要是由于施N、P后, 消除了矮嵩草草甸N和P素的限制作用, 促进了N、P利用率高的物种生长, 植被长势和盖度较高, 对地表起到小幅增温的作用, 能够满足植物对热量的要求[27], 而K在青藏高原高寒地区并不是限制因子, 施K对矮嵩草草甸群落影响并不显著, 所以地下5 cm和地上10 cm有效积温均小于N、P。
各物种在始花期、花期持续期和终花期上的不同反映了植物的时间生态位有一定程度上的分离[28]。对草地群落共存机制的研究表明, 植物时间上资源利用差异是维持群落物种多样性的重要机制[29], 相对优势种而言, 伴生种在生长季节内占有边缘生态位, 与优势种的时间生态位分离, 以减缓种间竞争。在本研究中, 禾草(垂穗披碱草、甘青剪股颖和冷地早熟禾)、莎草(矮嵩草、黑褐薹草和双柱头藨草)和美丽风毛菊的始花期、终花期均推迟, 而杂类草(麻花艽和鹅绒委陵菜)的始花期和终花期均提前(表3、图1, 表4、图2)。麻花艽和美丽风毛菊的花期提前可能是由于避免与禾草在资源上的竞争, 提前开花可以有效地确保繁殖部分的生长, 保证其未来在群落中的竞争力和生存。禾草和莎草是矮嵩草草甸优势种植物, 对土壤养分的吸收利用转化能力较临近的杂类草强[30, 31], 禾草和莎草花期推迟意味着植物延迟了从营养生长到繁殖生长转化的时间, 即植物个体营养部分增大或者克隆生长增加[15], 禾草和莎草在群落中具有更大的竞争优势。但是与莎草和杂类草相比, 禾草类花期推迟并没有使花期持续期也相应延长(表5), 这可能是由于在8月中旬后秋季寒冷天气到来之前高寒草甸群落中禾草植物种选择停止开花。我们认为在群落种间开花期分离可能是高寒草甸群落种间为降低种间资源和传粉竞争压力的进化分离响应机制。N、P、K添加后, 减小了垂穗披碱草-麻花艽、垂穗披碱草-美丽风毛菊、甘青剪股颖-麻花艽和冷地早熟禾-麻花艽在开花时间上的差异(图3), 降低了物候互补, 植物生长繁殖速率趋于同步性[28, 31]。
与添加K相比, 矮嵩草草甸添加N、P后群落物候变化较为剧烈, 促进了非相似性群落的形成(图4), 施肥后花期物候偏移所导致的群落生态位变化, 增强了植物间对限制性资源的竞争, 群落中出现生态位空洞, 为物种入侵、群落结构变化提供生态位间隙[14, 32, 33]。在花期物候时, 熵值法综合评价出垂穗披碱草、矮嵩草、麻花艽和美丽风毛菊是对N、P施肥响应较敏感的物种(表6), 需要对以后持续的肥料添加效应进行监测研究。
| 表6 熵值法综合评价结果 Table 6 The results of comprehensive evaluationby entropy method |
综上, 植物物候差异是植物生活史对策变化的体现, 在矮嵩草草甸上添加N、P、K后, 禾草(垂穗披碱草、甘青剪股颖和冷地早熟禾)、莎草(矮嵩草、黑褐薹草和双柱头藨草)和美丽风毛菊的始花期和终花期显著推迟, 而麻花艽和鹅绒委陵菜均明显提前, 但禾草的花期持续期却未明显延长, 矮嵩草草甸群落物候对添加N、P响应较为敏感, 对长期施肥的响应模式需要进一步研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [7] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [22] |
|
| [24] |
|
| [25] |
|
| [27] |
|
| [28] |
|
| [30] |
|
| [32] |
|
| [33] |
|