




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
外源甜菜碱对低温胁迫下结缕草生理特性的影响
[梁小红1  , 安勐颍
, 安勐颍1 , 宋峥2 , 徐广臣1 , 濮阳雪华1, * ]
, 安勐颍]
|
|


作者简介:梁小红(1979-),女,辽宁本溪人,实验师。E-mail:liang81511@163.com
甜菜碱作为一种重要的渗透调节物质,与低温胁迫下植物的抗逆性有着密切的联系。为了探讨不同浓度外源甜菜碱对低温胁迫下结缕草相关生理指标的影响,本实验以结缕草‘青岛’品种为实验材料,分别采用0,50,100和150 mmol/L 的甜菜碱进行叶面喷施,在8/2℃(白天/夜间)的培养箱中连续处理28 d,以正常管理(28/24℃)作为对照。结果表明:外源甜菜碱能够有效缓解低温胁迫下结缕草坪观质量和叶绿素含量的下降,减少电解质渗透率和丙二醛含量的升高,显著提高SOD、POD、CAT和APX的活性,从而降低H2O2和
Glycine betaine (GB) is an important compatible solute that is associated with stress tolerance in plants under low-temperature stress. To investigate the effect of exogenous GB on the physiological indexes of Zoysia japonica under low-temperature stress, GB at 0, 50, 100, and 150 mmol/L was applied as a foliar spray to ‘Qingdao’, a Z. japonica cultivar, before transferring the plants to growth chambers. The plants were grown under average temperatures of 8℃/2℃ (day/night) for 28 days in the low-temperature treatment, and under average temperatures of 28℃/24℃ (day/night) in the control. Exogenous GB alleviated the loss of turf quality and chlorophyll content, and reduced the increase in electrolyte leakage and malondialdehyde content in Z. japonica under low-temperature stress. Exogenous GB increased the activities of superoxide dismutase, peroxidase, catalase, and ascorbate peroxidase, leading to lower H2O2 and
结缕草属(Zoysia)是一种隶属禾本科(Gramineae), 虎耳草亚科(Chloridoideae)的多年生C4型草本植物, 主要分布于亚洲东部地区, 在澳洲、非洲等比较温暖的地区也有分布, 也是我国常用的多年生暖季型草坪草之一[1]。目前研究表明, 结缕草属主要包括11个种, 若干个变种和变型, 其中在我国自然分布的有5个种和2个变种, 它们分别是结缕草(Zoysia japonica)、中华结缕草(Zoysia sinica)、沟叶结缕草(Zoysia matrella)、细叶结缕草(Zoysia tenuifolia)和大穗结缕草(Zoysia macrostachya)5个种以及青结缕草(Zoysia japonica var. pollida)和长花结缕草(Zoysia sinica var. nipponica)2个变种[2]。结缕草叶片厚硬而近革质, 适宜温暖湿润性气候, 耐高温、耐阴性及耐旱性较强, 此外, 结缕草还具有耐盐碱, 抗病虫害, 耐粗放管理, 耐践踏、耐贫瘠等优良特性, 因此, 结缕草具有很强的适应性和抗逆性, 在我国被广泛应用于城市园林绿化、水土保持、边坡修复等诸多领域[2, 3, 4]。但是, 结缕草也具有绿期短的缺点, 在冬季会枯黄休眠, 从而大大降低了其观赏价值, 限制了其在过渡带及北方地区的推广和使用[5]。因而, 如何提高结缕草对低温胁迫的抵抗性, 延长结缕草的绿期, 已经成为国内外众多学者关注的问题之一。
甜菜碱(GB)是一种烷基烃类含氮化合物, 属于甘氨酸的季铵衍生物, 它可以和多种氨基酸结合形成新的复合物, 譬如甘氨酸甜菜碱、脯氨酸甜菜碱及组氨酸甜菜碱等, 其中甘氨酸甜菜碱是结构最为简单, 研究最为广泛的一类甜菜碱, 它广泛分布于动植物和微生物体内[6, 7]。植物受到逆境胁迫时, 甜菜碱可以作为渗透调节物质参与植物细胞的渗透调节, 亦可作为渗透保护物质来提高生物大分子结构和功能的稳定性[8]。因此, 在低温和盐渍等逆境胁迫下, 甘氨酸甜菜碱可以提高植物的抗逆性, 研究表明, 外源甜菜碱可以提高小麦(Triticum aestivum)、大麦(Hordeum vulgare)、烟草(Nicotiana tabacum)、黄瓜(Cucumis sativus)、玉米(Zea mays)等多种植物对低温胁迫的抵抗性[9, 10, 11, 12, 13]。然而, 目前关于外源甜菜碱对提高草坪草抗寒性的相关研究较少, 尤其是寒冷胁迫下, 外源甜菜碱对结缕草生理代谢影响的报道甚少。因此, 开展相关研究有利于进一步阐明甜菜碱对提高结缕草耐寒能力的生理机制, 从而延长结缕草的绿期, 为结缕草的进一步推广和应用奠定理论基础。
本研究选择结缕草‘ 青岛’ 为研究对象, 结缕草种子由青岛海源草坪有限责任公司提供。2013年7月将种子播种于直径16 cm, 高度15 cm的塑料花盆中, 播种量为20 g/m2, 花盆中的基质为草炭:蛭石:沙=4:2:1的混合物。播种后的植物材料在温室中培养4个月, 温室条件为光周期14 h, 光合有效辐射(PAR)400 μ mol/(m2· s), 温度白天28℃, 夜间24℃。在生长期间, 每3 d浇水1次, 每周采用半强度Hoagland营养液施肥1次[14], 成坪后每周修剪至约10 cm高。
在低温处理前3周, 所有的植物材料被转移至培养箱中, 培养箱的培养条件同温室条件。在2周的适应生长后, 即低温胁迫处理前1周, 处理组结缕草分别被喷施浓度为0, 50, 100, 150 mmol/L的甜菜碱。低温处理条件为光周期14 h, 光合有效辐射(PAR)400 μ mol/(m2· s), 温度白天8℃, 夜间2℃, 低温处理时间共28 d。以正常管理作为对照组。分别在低温处理后的第0, 7, 14, 21和28天取样, 液氮速冻后转移至-80℃冰箱进行保存。
坪观质量参照Turgeon[15]的方法, 采用1~9分制进行评分; 叶绿素含量采用95%乙醇提取测定[16]; 电解质渗透率采用Blum和Ebercon[17]的方法测定; 丙二醛含量采用Heath 和 Packer[18]的方法测定; 过氧化氢含量的测定采用文献[19]的方法; 超氧阴离子含量采用Jiang 和Zhang[20]的方法测定; 抗氧化酶活性的测定采用Zhang 和 Kirkham[21]的方法; 可溶性蛋白含量的测定采用Bradford[22]的方法; 脯氨酸含量采用李合生[23]的方法测定。
实验设置4个重复, 采用Excel 2013和SPSS 18.0进行统计分析, 并使用Excel 2013作图。
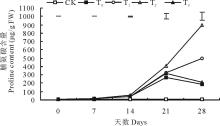
2.1.1 坪观质量和叶绿素含量 低温胁迫导致结缕草的坪观质量和叶绿素含量呈显著下降趋势(P< 0.05), 在第28天时, 结缕草坪观质量为4.7分, 处于不可接受状态, 叶绿素含量与对照相比下降了71.71%(图1)。与单独低温胁迫相比, 外施3种不同浓度的甜菜碱都在一定程度上缓解了坪观质量和叶绿素含量的下降, 其中, 外施100 mmol/L甜菜碱的缓解效果最为显著, 而外施150 mmol/L甜菜碱缓解作用较小, 尤其是对叶绿素含量的影响。在第28天时, 100 mmol/L甜菜碱预处理的结缕草坪观质量得分为6.7分, 依然处于可接受状态, 叶绿素含量相比单独低温胁迫升高了1.21倍。
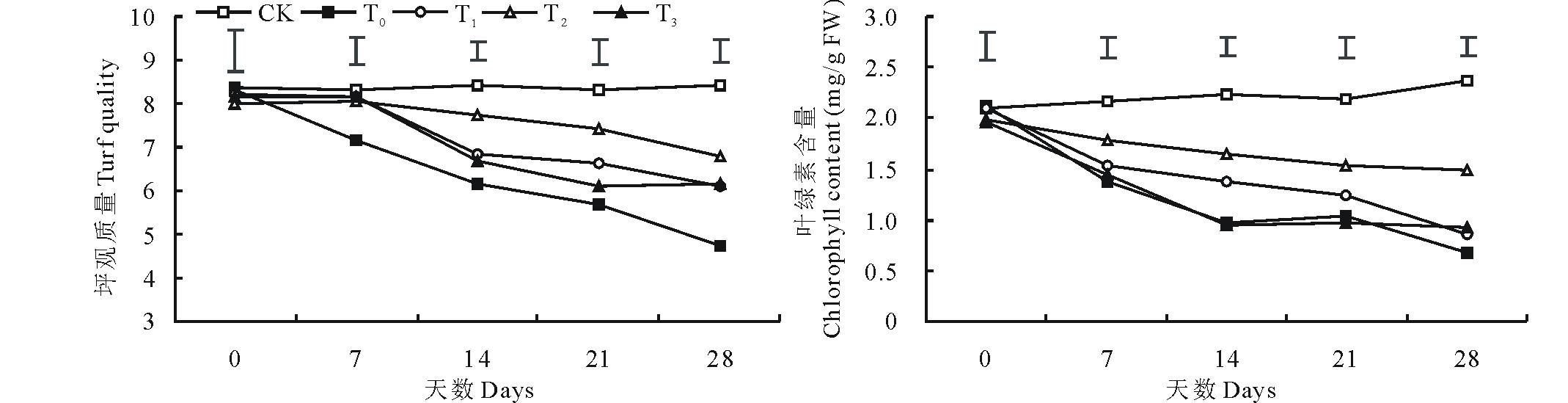 | 图1 外施不同浓度甜菜碱对低温胁迫下结缕草坪观质量和叶绿素含量的影响CK(对照组)=常温+0 mmol/L 甜菜碱; T0=低温胁迫+0 mmol/L甜菜碱; T1=低温胁迫+50 mmol/L甜菜碱; T2=低温胁迫+100 mmol/L甜菜碱; T3=低温胁迫+150 mmol/L甜菜碱。图中上方的垂直线条表示同一时间下不同处理之间的标准误(P< 0.05)。下同。Fig.1 Effects of exogenous glycine betaine (GB) with different concentrations on turf quality and chlorophyll content of Z. japonica under low-temperature stressCK (control)=normal temperature+0 mmol/L GB; T0=low temperature+0 mmol/L GB; T1=low temperature+50 mmol/L GB; T2=low temperature+100 mmol/L GB; T3=low temperature+150 mmol/L GB. Vertical bars on the top indicate least significant difference values (P< 0.05) for treatment comparison at a given day of treatment. The same below. |
2.1.2 电解质渗透率和丙二醛含量 由图2可知, 低温胁迫导致结缕草的电解质渗透率和丙二醛含量呈显著上升趋势(P< 0.05), 到第28天时, 单独低温胁迫的结缕草电解质渗透率和丙二醛含量与对照相比分别增加了185.84%和99.47%。外施3种不同浓度的甜菜碱都在一定程度上缓解了低温胁迫引起的电解质渗透率和丙二醛含量的上升, 而且, 外施100 mmol/L甜菜碱的缓解效果最为显著。在第28天时, 与单独低温胁迫相比, 100 mmol/L甜菜碱预处理的结缕草电解质渗透率和丙二醛含量分别降低了44.9%和23.6%。
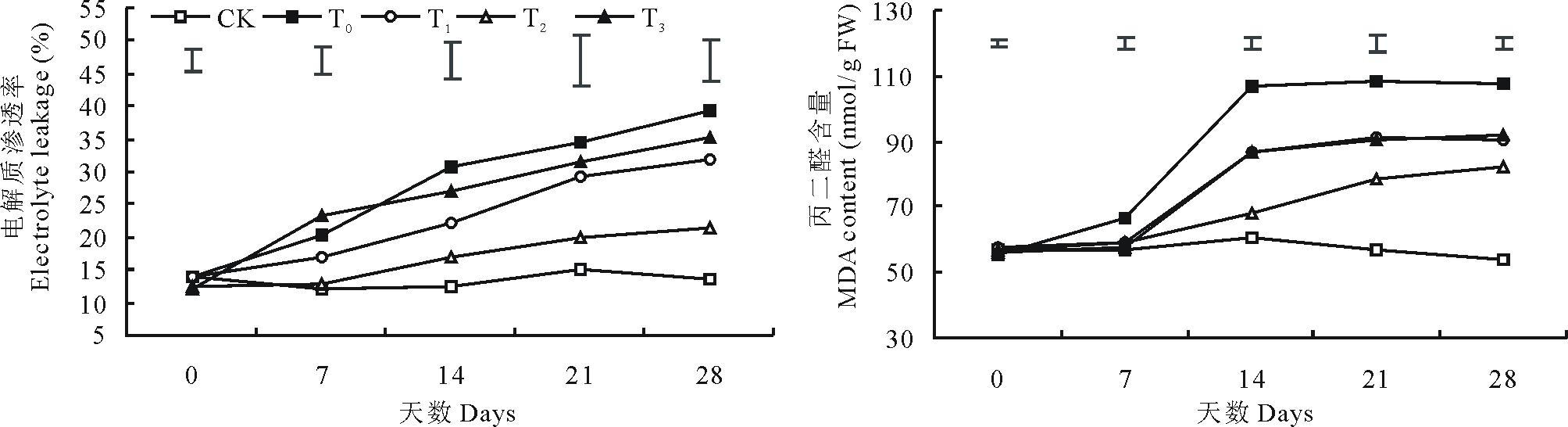 | 图2 外施不同浓度甜菜碱对低温胁迫下结缕草电解质渗透率和丙二醛含量的影响Fig.2 Effects of exogenous glycine betaine with different concentrations on electrolyte leakage and MDA content of Z. japonica under low-temperature stress |
2.2.1 过氧化氢和超氧阴离子含量 低温胁迫导致结缕草的H2O2含量和
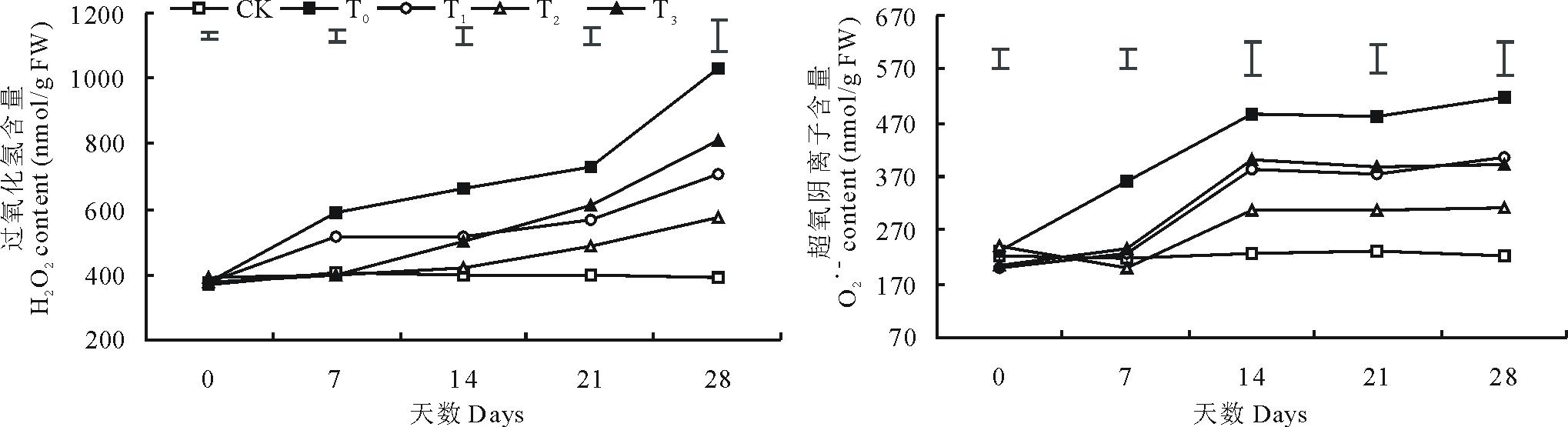 | 图3 外施不同浓度甜菜碱对低温胁迫下结缕草H2O2含量和 |
2.2.2 抗氧化酶活性 由图4可知, 低温胁迫显著提高了结缕草4种抗氧化酶的活性(P< 0.05)。低温胁迫处理条件下, 超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)和抗坏血酸过氧化物酶(APX)的活性分别在第14, 28, 21和28天时达到峰值, 与对照相比其活性分别提高了151.09%, 67.18%, 110.99%和72.22%。与单独低温胁迫处理相比, 外施3种不同浓度的甜菜碱均在不同程度上提高了4种抗氧化酶的活性, 其中100 mmol/L 的外源甜菜碱作用效果最为显著, 而150 mmol/L的外源甜菜碱作用效果较小, 尤其是对CAT和APX活性的影响。相比单独低温胁迫, 100 mmol/L甜菜碱预处理的结缕草SOD、POD、CAT和APX的活性分别在第14, 28, 21和28天时增加了82.5%, 47.4%, 77.2%和68.5%。
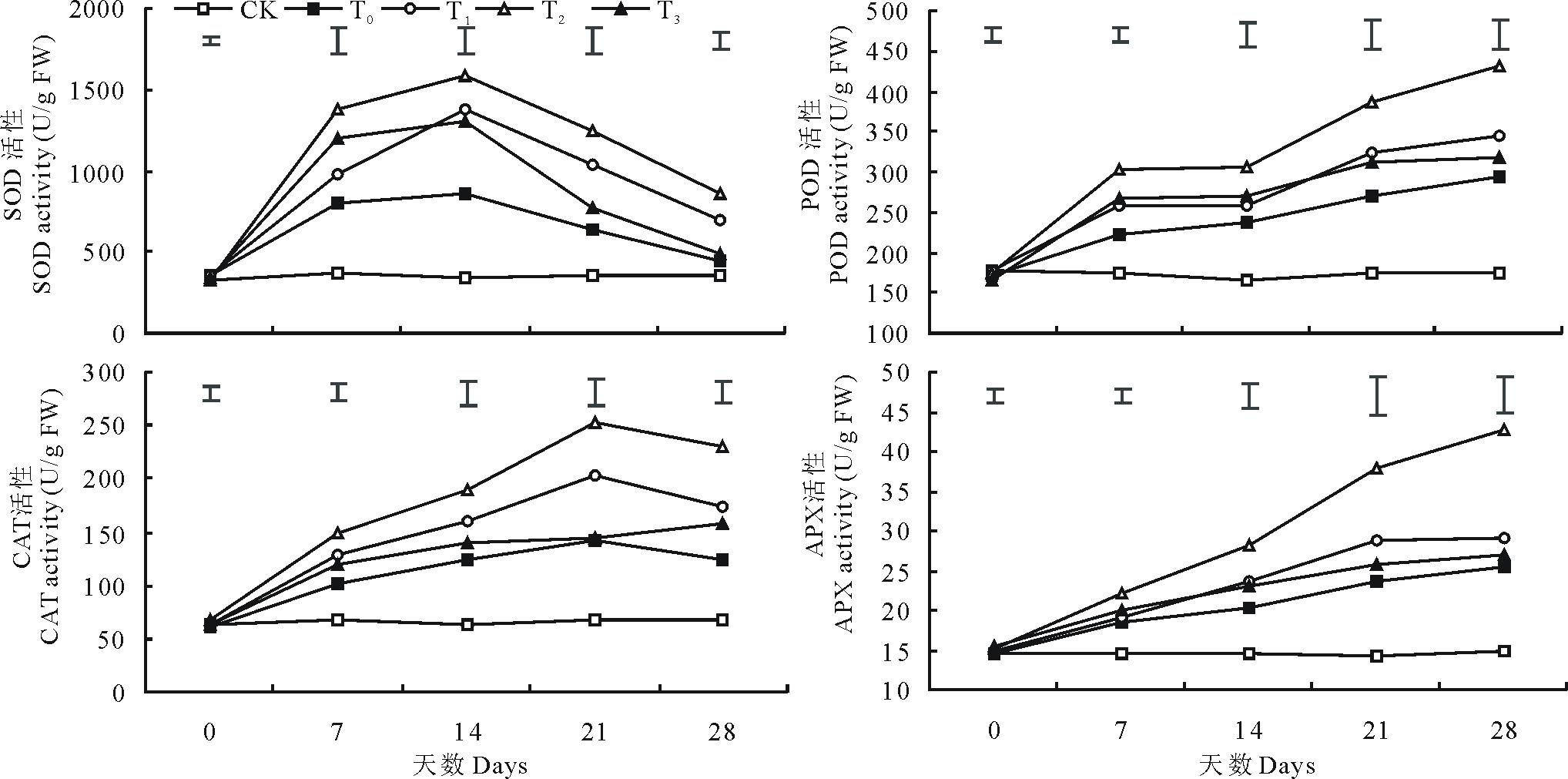 | 图4 外施不同浓度甜菜碱对低温胁迫下结缕草抗氧化酶活性的影响Fig.4 Effects of exogenous glycine betaine with different concentrations on antioxidant enzyme activity of Z. japonica under low-temperature stress |
2.3.1 可溶性蛋白含量 低温胁迫下结缕草的可溶性蛋白含量呈现先增加后下降的趋势, 其含量在第14天时达到峰值, 与对照相比增加了94.74%(图5)。同单独低温处理相比, 外施100 mmol/L甜菜碱显著提高了结缕草的可溶性蛋白含量, 在第14天时增加了72.7%, 而外施50和150 mmol/L甜菜碱对低温胁迫下结缕草可溶性蛋白含量的影响并不显著。
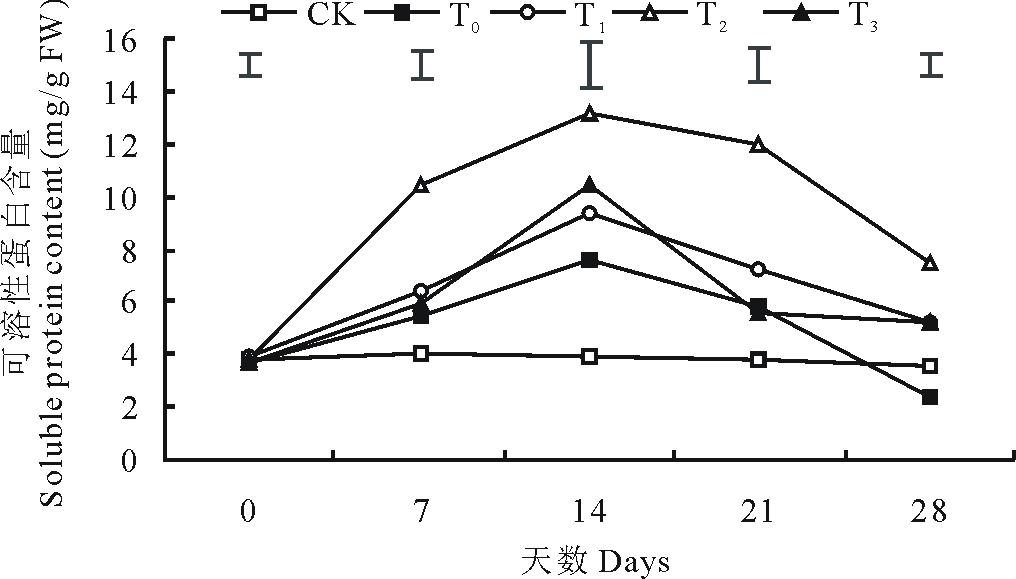 | 图5 外施不同浓度甜菜碱对低温胁迫下结缕草可溶性蛋白含量的影响Fig.5 Effects of exogenous glycine betaine with different concentrations on soluble protein content of Z. japonica under low-temperature stress |
2.3.2 脯氨酸含量 从第14天开始, 低温胁迫显著提高了结缕草的脯氨酸含量(P< 0.05), 在第21天时, 单独低温胁迫的结缕草脯氨酸含量达到峰值, 与对照相比提高了22.46倍, 此后其含量开始呈现下降趋势(图6)。在第28天时, 与单独低温胁迫处理相比, 外施50和100 mmol/L的甜菜碱分别使脯氨酸含量提高了1.65和3.73倍, 而外施150 mmol/L的甜菜碱与单独低温胁迫处理相比没有显著差异。
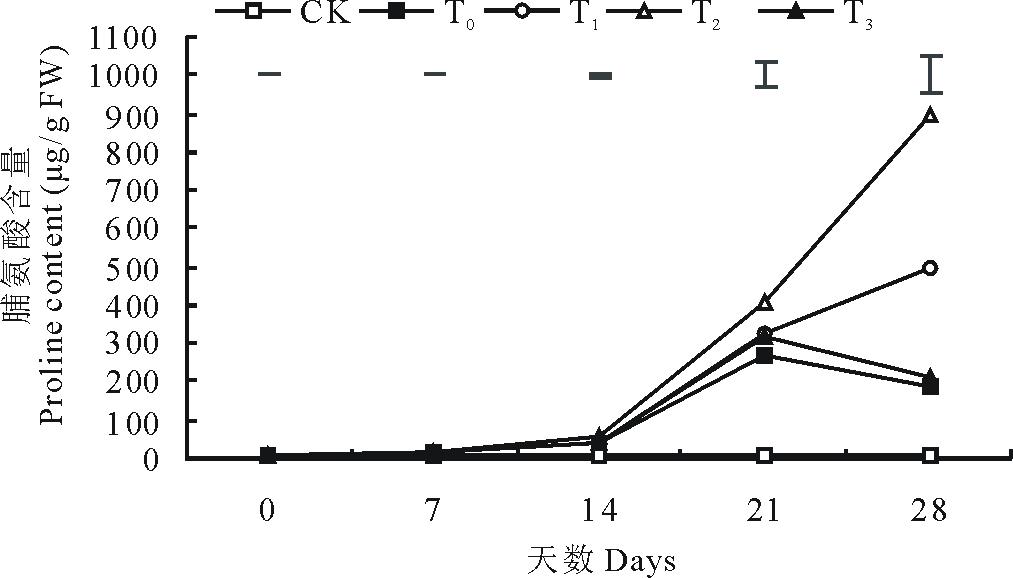 | 图6 外施不同浓度甜菜碱对低温胁迫下结缕草脯氨酸含量的影响Fig.6 Effects of exogenous glycine betaine with different concentrations on proline content of Z. japonica under low-temperature stress |
坪观质量是评价草坪质量好坏最直接的指标, 更是评价草坪优劣的综合性指标[24]。从本研究的结果可以看出, 低温胁迫会显著降低结缕草的坪观质量, 而外施甜菜碱则可有效的缓解低温胁迫对坪观质量的影响, 使结缕草保持较高的坪观质量。叶绿素是植物光合作用中不可缺少的重要物质, 更是反应叶片受伤程度最为直接的指标[25, 26]。本实验中, 低温胁迫显著降低了结缕草的叶绿素含量, 这与前人的研究结果相一致[27, 28]。这可能是由于低温胁迫不仅破坏了叶绿体的超微结构, 导致光合色素降解, 而且抑制了叶绿素的合成, 从而降低了叶绿素的含量。外施甜菜碱则显著缓解了低温胁迫下结缕草叶绿素含量的降低, 说明外施甜菜碱可以保护低温胁迫下结缕草的叶绿体结构, 从而使结缕草的叶绿素含量保持较高水平。
细胞膜系统是植物感受低温最敏感的部位, 也是植物抵抗低温伤害的关键结构。电解质渗透率和丙二醛含量通常可以有效地反映出细胞膜系统的伤害程度和膜脂的过氧化程度[29, 30]。本研究表明, 低温胁迫显著增加了结缕草细胞膜的电解质渗透率和丙二醛含量, 说明低温胁迫导致了膜脂过氧化反应, 破坏了结缕草的细胞膜系统。研究表明, 甜菜碱能够防止细胞膜的热变性, 降低原生质膜从液晶相转变为凝胶相的温度, 从而对低温胁迫下的细胞膜的完整性起到保护作用[31]。也有研究指出, 甜菜碱可以稳定低温胁迫下的细胞膜结构是由于甜菜碱的季铵化阳离子特性可以结合细胞膜上的羟基阴离子[32]。本实验中, 外施甜菜碱显著降低了低温胁迫下结缕草的电解质渗透率和丙二醛含量, 说明外施甜菜碱能够缓解低温胁迫下结缕草的膜脂过氧化反应, 对结缕草细胞膜系统的完整性起到了一定的保护作用, 进而提高了结缕草对低温胁迫的抵御能力。
低温胁迫下植物体内积聚的活性氧是导致膜脂过氧化反应, 细胞膜结构遭到破坏的重要原因, 其中H2O2和
可溶性蛋白和脯氨酸是植物体内的2种重要的渗透调节剂。可溶性蛋白和脯氨酸本身都具有很强的亲水性, 其含量的增加会提高细胞内束缚水的含量, 进而保护细胞膜免受脱水的伤害, 来达到提高植物抵御低温胁迫能力的目的[38]。本研究表明, 低温胁迫显著增加了结缕草的可溶性蛋白和脯氨酸含量, 并且外施甜菜碱进一步增加了其含量。相关研究指出, 增加的可溶性蛋白很可能是一些植物抵御低温胁迫相关的或有专一性的功能蛋白酶[39]。也有研究表明, 甜菜碱可能在加速低温胁迫下受到损伤的蛋白质的从头合成过程中起到一定作用[40]。因此, 外施甜菜碱能够通过可溶性蛋白和脯氨酸的渗透调节作用, 进一步提高结缕草对低温胁迫的抵抗性。
综上所述, 外源甜菜碱能够有效缓解低温胁迫下结缕草坪观质量和叶绿素含量的降低, 减少电解质渗透率和丙二醛含量的升高, 一方面是通过提高结缕草的抗氧化代谢能力, 增强了SOD、POD、CAT和APX的活性, 从而清除了多余的H2O2和
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [12] |
|
| [13] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [33] |
|
| [34] |
|
| [38] |
|
| [39] |
|