


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
青海湖流域植被物候格局时空动态变化及其与植被退化的关系
[李广泳1 , 姜翠红2, *  , 程滔
, 程滔1 , 张浩然1 , 陈占涛3 ]
, 程滔|
|


作者简介:李广泳(1979-),男,山东东平人,工程师。E-mail: liguangyong@ngcc.cn
本研究利用2000-2014年期间MODIS 16 d合成EVI 数据,选用阈值法模型提取了青海湖流域逐年的植被关键物候期,分析了物候时空动态变化特征及其与植被退化的关系。结果表明,青海湖流域内植被陆续在4月中旬-6月中旬进入返青阶段,8月中旬-10月中旬进入枯黄期,生长季长度在100~150 d之间。植被陆续进入返青期的时间在流域整体空间上呈现由东南向西北延迟的水平地带性变化趋势,多高山分布区域的植被进入返青期时间表现出垂直非地带性特征。植被进入枯黄期的时间与进入返青期早晚的时间在空间格局上相反,生长季长短空间格局与植被进入返青时间早晚的空间格局一致。气候变化趋势下,流域植被物候时空动态变化存在空间异质性,返青期均值年际变化趋势不明显,枯黄期均值提前6.4 d,生长季均值缩短8.9 d。流域内植被物候除受气候因子影响外,与植被退化也有密切关系。植被生长趋于茂盛,返青期呈现延迟趋势;植被生长退化趋势,返青期呈现提前倾向。枯黄期与植被退化的关系较复杂,整体上未表现出明显一致的相关性。植被生长季与植被退化呈现强负相关,即植被生长呈现茂盛趋势的区域,生长季缩短。
This paper presents a threshold method to examine the vegetation phenology in a Qinghai Lake watershed based on a time-series analysis of a MODIS EVI dataset from 2000 to 2014 using 16-day intervals. The spatio-temporal variation of vegetation phenology and the relationship with vegetation degradation were also analyzed. The results showed that the beginning of the vegetation growing season (BGS) occurred from mid-April to mid-June and the end of growing season (EGS) from mid-August to mid-October; growing season (GS) was between 100 to 150 days. The BGS spatial pattern was embodied by a transition from southeast to northwest; vertical zonation in the mountainous region showed a contrary trend in EGS and GS. Spatio-temporal variation of vegetation phenology at the whole watershed level showed spatial heterogeneity under climate change, but mean BGS trends were not pronounced during the study period. EGS was advanced approximately 6.4 days and GS prolonged by 8.9 days. Vegetation phenology was also affected by vegetation degradation. BGS tended to be later when vegetation flourished and advanced when vegetation was degraded. The relationship between annual EGS date and vegetation degradation was relatively complex. EGS and vegetation degradation were significantly negatively correlated indicating EGS was earlier where the vegetation was in good condition.
植被物候研究周期性重复出现的植被自然现象及其与环境条件周期变化的相互关系, 反映了地球生物圈对地球气候和水循环季节、年际变化的响应[1], 被认为是植被响应区域气候变化最直接、敏感的特征指标之一; 另一方面, 植被物候通过很强的地表生物物理、化学过程控制当前和未来的气候模式以及陆地生态系统的碳循环[2]。因此, 科学的理解植被物候空间格局及时空动态变化特征, 不仅利于准确预测未来的气候变化趋势, 更利于掌握区域-全球尺度下受植被物候信息控制的碳收支空间格局及动态变化特征。
当前对植被物候的监测以野外定点观测和遥感监测为主[3], 研究内容多集中在全球变化趋势下植被物候变化特征及其与气候因子的关系, 尤其是关于植被返青期的相关研究较多[4]。在利用遥感技术研究植被物候过程中, 多数学者仍局限在通过选取植被样区(点)定点监测植被物候变化, 分析样区植被物候的年际变化规律及其与气候因子之间的关系[5, 6, 7, 8]。在植被样区尺度上研究植被物候变化的问题较简单, 而在空间尺度下, 植被物候还受其他自然和人为压力等潜在机制的影响, 如植被退化、放牧干扰等[9, 10, 11], 这些问题被认为是未来一段时间植被物候研究的新方向[2, 12]。进行植被物候时空动态变化格局研究是探究上述其他影响植被物候潜在因子的前提。长期以来, 气候变化被认为是影响干旱、半干旱区植被物候的关键因素, 忽略了植被退化对物候影响的重要性[13]。Chen等[14]认为, 青藏高原的草原、草甸植被退化会导致春季返青的延迟, 但没有进行相关具体的研究来进行证明。宋春桥等[8]分析了藏北高原典型植被样区物候变化、生长季最大增强植被指数(enhanced vegetation index, EVI)与气候因子之间的关系, 但没有对植被退化与植被物候之间的关系进行深入分析。李广泳等[11]虽然分析高寒草地植被物候与植被退化之间的关系, 但是研究仍局限在样点监测, 缺乏植被物候与植被退化空间关系的研究。在当今全球气候变化的背景下, 青藏高原已经被科学界公认为进行人类活动和生态环境冲突研究的重要试验基地[15]。尤其是近10年来, 对该区域植被物候的研究成为气候生态学领域在青藏高原研究的新热点。青藏高原在气候变化和人类活动的双重干扰下, 植被退化与茂盛趋势并存[16], 然而针对植被退化与物候时空动态变化之间的关系研究尚存在不足。
青海湖流域位于青藏高原和亚洲半干旱气候敏感带, 具有独特的高寒半干旱生态系统特征, 是一个综合性的自然、社会和经济复合生态圈。近30年来, 青海湖流域年平均气温增幅为0.28℃/10年[17], 明显超过同时期青藏高原地区乃至北半球同纬度其他地区的平均水平。在全球气候变化日益明显趋势下, 青海湖流域已经成为国际科学界研究青藏高原众多生态环境问题的代表区域之一[18]。本研究选取青海湖流域高寒植被为研究对象, 研究了近15年来植被物候时空动态变化特征, 重点分析了植被退化与植被物候时空动态变化之间的关系, 以期为气候变化趋势下国家实施的青海湖流域生态治理与保护工程提供理论支撑。
青海湖流域位于青藏高原的东北部(36° 15'-38° 20' N, 97° 50'-101° 20' E)(图1), 面积为29661 km2, 海拔3194~5174 m, 属于典型的干旱半干旱气候区。流域内年平均温度范围为-0.8~1.1℃, 年降水量范围为324.5~412.8 mm, 呈现由东北向西南降低的趋势。地势西北高而东南低, 地貌类型复杂, 多高山分布, 山区面积占流域总量的68.6%。该区域内的青海湖是我国最大的内陆湖泊, 也是最大的咸水湖, 湖泊覆盖面积约占流域总面积的16.0%以上, 注入青海湖的主要河流有布哈河、沙柳河、哈尔盖河、黑马河和倒淌河, 其中布哈河是注入青海湖最大的河流, 占流域内年总径流量的48.8%[19]。流域内土地覆盖类型多样, 草地、灌丛、荒漠、水域及农田都有分布, 其中草地是流域主要覆盖类型, 约占流域面积的70%(图1)。
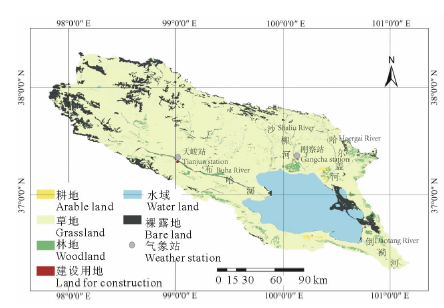 | 图1 青海湖流域植被覆盖(2012年)Fig.1 Land cover map of Qinghai Lake during 2012 |
遥感数据为来源于美国国家航空航天局(NASA)网站(http://ladsweb.nascom.nasa.gov/data/)搭载在Terra和Aqua卫星上的MODIS传感器获取的MOD13Q1 16 d 合成的植被指数产品, 空间分辨率为250 m。本研究选用的是增强植被指数(enhanced vegetation index, EVI), 时间跨度为2000-2014年, 共计684景。选用MODIS Reprojection Tool软件对所有影像进行投影转换批处理, 然后采用ERDAS 软件对同时相2景影像进行拼接、裁切, 得到研究需要的EVI数据。流域植被覆盖数据采用国土部门提供的2012年土地利用现状更新数据为基础, 结合同期Landsat卫星影像数据进行了修正。刚察、天峻气象站是流域内仅有的2个国家级气象站, 研究中涉及的2000-2014年的温度、降水量数据来源于2个气象站地面观测资料。
近20年来, 遥感技术已经成为在大尺度下监测植被物候的有效手段。本研究首先在ArcGIS环境下将ModisEVI数据进行标准化, 然后采用非对称高斯函数拟合方法对标准化后的EVI数据进行时间序列上的重建, 选用阈值法[20]分别提取研究期内历年植被物候期(返青期、枯黄期和生长季)。结合前期研究, 将青海湖流域植被返青期的阈值设置为0.1, 枯黄期阈值设置为0.5[11]。植被物候期计算以同年1月1日为起点进行标准化, 即1月1日为第1天, 1月2日为第2天……, 依此类推确定植被返青期、枯黄期, 返青期与枯黄期之间的自然天数即为生长季。阈值法模型如下:
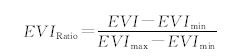 |
式中, EVIRatio为输出比值(值域为0~1), EVI为标准增强植被指数(enhanced vegetation index)值, EVImin指年内最小值, EVImax为年内最大EVI值。
一元回归趋势法是对随时间变化的变量进行回归分析的模型[21], 已经广泛应用到植被动态变化研究中, 取得了良好的结果。该模型应用ArcGIS栅格计算功能, 基于栅格单元来分析植被物候逐年的变化趋势, 从而反映植被物候的时空动态变化量。其模型如下:
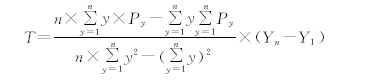 |
式中, n是研究期总年数; y为研究期中第y年; Py为第y年的物候数据; Yn为研究期末年; Y1为研究期第1年; T为物候期的变化量。T> 0, 说明物候期延迟(延长); T< 0, 说明物候期提前(缩短)。
Pearson相关性系数是表达两个变量之间直线联系紧密程度的系数, 可以在ArcGIS运行环境下将两套时间序列相同的数据基于像元进行相关性分析[22]。相关系数越接近于1或-1, 相关度越强, 相关系数越接近于0, 相关度越弱。相关性系数为正值, 表示两变量之间存在正相关, 反之为负相关。通常情况下相关系数绝对值在0.8~1.0 为极强相关, 0.6~0.8为强相关, 0.4~0.6为中等程度相关, 0.2~0.4为弱相关, 0.0~0.2为极弱相关或无相关。皮尔逊相关性系数模型如下:
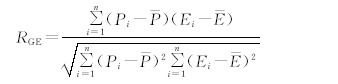 |
式中, RGE为植被物候与EVI的相关系数, Pi为第i年物候期, 为研究期内平均物候期, Ei为第i年EVI, 为研究期内平均EVI。
2.1.1 植被返青期空间格局 2000-2014年期间, 青海湖流域植被平均返青期为第144.6天, 植被最早进入返青的时间为4月中下旬(100~120 d)。截止到6月上旬(160 d), 流域内95.0%的植被进入返青期, 整个进入返青期历时约60 d(图2)。受流域地势变化的影响, 植被进入返青期时间在空间上呈现由东南向西北延迟的水平地带性变化趋势。青海湖南岸、倒淌河子流域及布哈河入湖口海拔较低的区域(3200~3300 m), 土壤湿润, 是流域内植被最早进入返青期的地区。5月上旬(120~130 d), 随着温度的升高, 植被返青开始向四周较高海拔区域扩展, 约占流域植被覆盖总面积的8.3%。尤其在北部多高山分布地区, 植被进入返青期时间表现出垂直非地带性, 海拔较低、水分条件较好的河流沟谷地带的植被陆续进入返青期; 5月中旬至6月上旬(130~160 d)是流域植被返青高峰期, 85.9%的植被返青集中在该时间段内。受高海拔低温的影响, 流域西北部海拔在3800 m以上的高寒荒漠植被, 一般集中在6月中旬(160~170 d)。青海湖北岸、南岸种植油菜(Brassicacampestris)的农田返青期集中在6月上旬(150~160 d), 主要是当地农民通常选择在5月下旬播种所决定。
2.1.2 植被枯黄期空间格局 流域内植被平均枯黄期为第268.7天, 大约在8月中旬植被开始陆续进入枯黄期, 10月中旬结束。流域植被全部进入枯黄期历时约60 d(图2)。流域内植被在先后进入枯黄期空间格局上呈现由西北向青海湖四周低海拔区域扩展的变化趋势, 与进入返青时间的空间格局相反。8月中旬(220 d)开始, 受高海拔地区降雪等因素的影响, 西北部高寒荒漠稀疏植被地区植被最早进入枯黄期。随着秋季来临, 大气温度降低, 降水量的减少, 9月份流域内植被进入枯黄期的面积开始增加。截止到9月中旬(260 d), 植被进入枯黄期的面积占流域植被覆盖总面积的15.1%, 集中分布在流域周边和中部高海拔区域。9月底(270 d), 流域内55.16%的植被进入枯黄期, 集中分布在海拔3600 m以上的区域。10月中旬(280~290 d), 流域内98.8%的植被进入枯黄期。流域北部山区的植被进入枯黄期的时间同样表现出了垂直非地带变化特征, 生长在高海拔区域植被进入枯黄期的时间较低海拔区域早。农田内种植的农作物进入枯黄期时间集中在9月中下旬(250~270 d), 较周边草地略提前。
2.1.3 植被生长季空间格局 植被返青期、枯黄期共同决定生长季的长短。流域内植被平均生长季为124.6 d, 其中生长季分布在100~150 d之间的植被占流域覆盖总面积的83.1%(图2)。研究期内青海湖流域植被平均生长季短于90 d的区域集中分布在西北部高海拔区域。主要由于该区域植被受高海拔低温影响, 植被返青较晚, 且进入8月份, 多降雪天气, 部分植物提前进入枯黄期导致其生长季较短。植被生长季超过130 d的区域集中分布在布哈河下游、倒淌河子流域及紧邻青海湖四周海拔低于3600 m的区域。尤其在青海湖周边和倒淌河子流域海拔低于3300 m的区域, 是流域内地表湿热条件最好的区域, 植被生长季超过150 d。农田内植被生长季短于流域植被平均生长季, 仅为117.7 d。
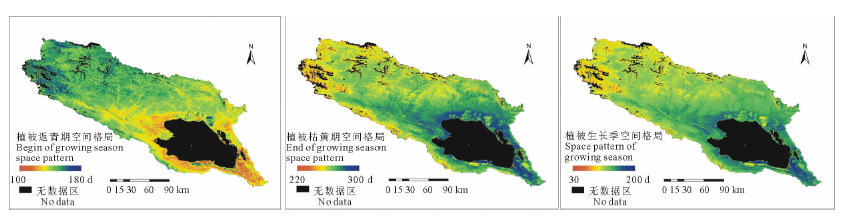 | 图2 青海湖流域植被物候空间格局(2000-2014年)Fig.2 Spatial pattern of vegetation phenology in Qinghai Lake watershed during 2000to2014 |
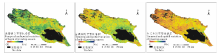
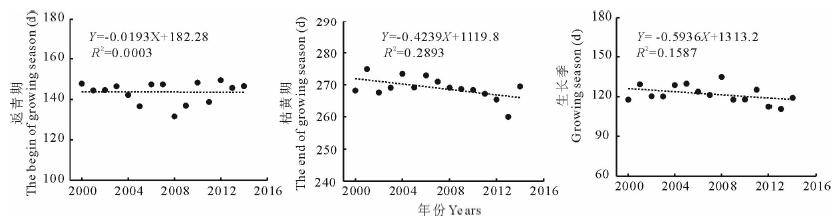
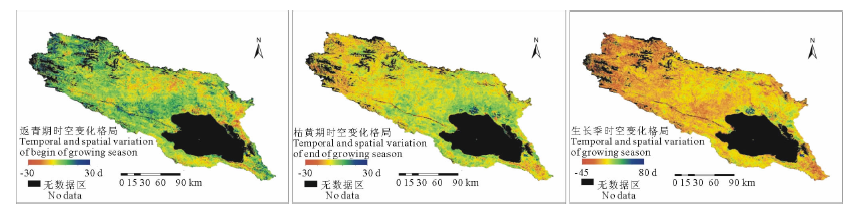
2.2.1 植被返青期时空变化特征 根据图3, 气候变化趋势下, 2000-2014年期间青海湖流域植被平均返青期只表现出自然的波动, 但在地理空间上却表现出变化趋势的明显差异性(图4)。流域西北部、青海湖西侧及倒淌河子流域东部植被表现出明显的返青期延迟趋势, 超过10 d以上, 约占流域植被覆盖总面积的16.0%。流域中北部区域、青海湖东北部及南部区域却呈现植被返青提前趋势, 提前6 d以上, 面积约占流域植被面积的25.1%。受土地利用变化的影响, 实施退耕还林、还草的农田区植被物候明显提前, 大约10~20 d。
2.2.2 植被枯黄期时空变化特征 研究期流域内历年平均枯黄期提前6.4 d, 速率为4.2 d/10年(R2=0.2893)(图3)。在空间上植被枯黄期年际变化趋势并不一致(图4)。植被枯黄期延迟超过2 d的区域集中在紧邻青海湖北岸、南岸及布哈河中、下游地区也有零星分布, 约占流域植被覆盖总面积的17.3%。尤其在紧邻青海湖北岸、东岸实施退耕还林的区域, 枯黄期延迟超过10 d以上。植被枯黄期提前超过2 d的区域在流域内分布广泛, 占流域植被覆盖总面积的64.2%。流域西北部、布哈河子流域是枯黄期提前趋势最明显的区域, 其中枯黄期提前超过10 d的区域占流域植被覆盖总面积的21.8%。
 | 图3 青海湖流域植被物候变化趋势(2000-2014年)Fig.3 The vegetation phenology trends in Qinghai Lake watershed during 2000 to 2014 |
 | 图4 青海湖流域植被物候时空动态变化空间格局(2000-2014年)Fig 4 Spatio-temporal variation of vegetation phenology in Qinghai Lake watershed during 2000 to 2014 |
2.2.3 植被生长季时空变化特征 植被返青期、生长季时空变化的空间差异性决定植被生长季变化趋势在空间上同样具有明显的差异。15年内青海湖流域植被平均生长季缩短8.9 d, 速率为5.9 d/10年(R2=0.1587)。生长季延长超过2 d的区域以流域中北部、湖北岸和南岸植被为主, 占流域植被覆盖面积的30.4%, 其中部分区域生长季超过6 d, 占20.3%。退耕还林、还草区是生长季延长趋势最明显的区域, 延长时间超过20 d以上。生长季缩短的区域集中分布在流域中、西部和东部倒淌河子流域, 缩短超过2 d的区域占流域植被覆盖面积的57.5%以上, 其中44.9%的区域植被生长季缩短超过6 d以上, 集中分布在流域西北高寒荒漠植被覆盖区、湖西北岸及倒淌河流域内。
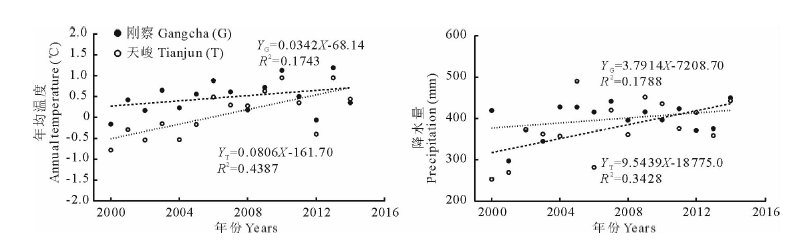
根据图5所示, 2000-2014年期间, 刚察、天峻2个气象站年均温度、降水量都呈明显增加趋势, 且天峻气象站观测的气候变化趋势较刚察气象站更显著, 变化速率均值分别为0.57℃/10年和66.68 mm/10年, 气候变化暖湿趋势明显, 与整个青藏高原气候变化趋势一致[23]。在暖湿的气候变化趋势下, 流域植被平均返青期变化趋势并不明显, 且仅与年降水量具有弱负相关性, 相关性系数分别为0.1303和0.2140, 与气象站年均温度无相关性, 说明青海湖流域年均温度升高背景下流域内植被平均返青期并未表现出提前趋势。流域植被平均枯黄期与天峻年均温度具有负弱相关性(R2=0.3091), 与刚察(R2=0.1626)、天峻年降水量(R2=0.2382)具有负相关性, 说明年降水量在一定程度上影响着流域植被枯黄期。植被平均生长季与天峻气象站温度(R2=0.2280)、降水量(R2=0.1381)具有弱负相关性, 相关性明显优于刚察。
 | 图5 刚察、天峻年均温度、降水量变化趋势(2000-2014年)Fig.5 The average annual temperature and precipitation trends of Gangcha and Tianjun station during 2000 to 2014 |
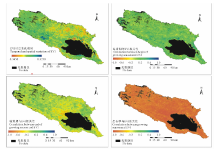
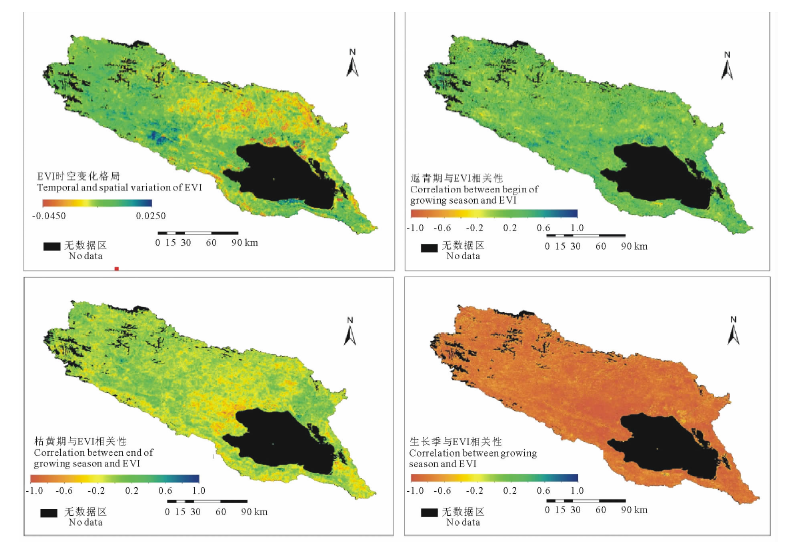
研究中选用2000-2014年历年年内最大EVI的变化趋势来反映青海湖流域植被退化趋势。根据图6中年内最大EVI时空变化格局显示, 在气候变化、人类活动等双重因子的干扰下, 青海湖流域植被动态变化空间异质性较大。西部区域植被的EVI呈现较明显的增加趋势, 说明该区域植被生长趋于茂盛, 尤其在布哈河中游, 植被茂盛趋势尤其显著。流域中北部地区, 植被的EVI出现大面积降低趋势, 说明该地区部分区域植被退化趋势较明显。紧邻青海湖北岸、东岸的退耕还草、还林区, EVI降低趋势显著, 主要是由于原种植的油菜等农作物被草地、稀疏灌丛植被替代, 导致植被地表覆盖度降低。
根据图6, 从整个流域来看, 植被返青期与EVI时空年际变化整体上呈现较好的正相关性, 28.0%的植被返青期与EVI年际变化存在弱正相关性(0.2< RGE< 0.4), 21.21%的植被返青期与EVI呈现中度正相关性(0.4< RGE< 0.6), 仅有8.39%的植被返青与EVI年际变化负相关指数达到0.2以上, 呈点状散布在流域内。这说明青海湖流域内植被退化趋势影响着植被返青期的变化, 植被退化趋势会导致返青期提前, 植被茂盛趋势, 返青期延迟。植被枯黄期与EVI时空动态变化趋势上的关系空间异质性较大。布哈河下游、倒淌河子流域及沙柳河中游小部分区域植被枯黄期与EVI时空变化呈现弱负相关性(0.2< RGE< 0.4), 占流域植被覆盖面积的21.54%; 11.88%的植被枯黄期与EVI呈现弱正相关性(0.2< RGE< 0.4), 集中分布在流域东北、西北部区域; 其余49.00%的植被枯黄期未表现出与植被退化的相关性。流域内植被生长季表现与EVI年际变化呈现较强的负相关性, 其中24.76%植被生长季与EVI呈现强负相关性(0.6< RGE< 0.8), 68.08%表现出极强的负相关性(0.8< RGE< 1), 说明气候变化趋势下, 青海湖流域植被茂盛与生长季之间具有紧密的联系, 植被退化趋势, 生长季呈现延长, 反之依然。
 | 图6 青海湖流域植被EVI时空变化格局及与植被物候的相关性(2000-2014年)Fig.6 Spatio-temporal variation of EVI and its relevance to vegetation phenology in Qinghai Lake watershed during 2000 to 2014 |
植被作为联结土壤、水体和大气之间物质和能量交换的关键环节, 其物候时空格局直接反映环境条件季节和年际变化, 并随着气候纬度、经度和海拔变化的规律性而表现出推移性的特点[24], 尤其是青藏高原高寒草地物候与海拔具有密切关系[25]。青海湖流域地势整体呈现西北高而东南低, 决定了植被物候格局在流域整体空间表现出水平地带性变化规律; 局部区域多高山分布, 高差较大, 气候的垂直变化特征决定了植被物候期的垂直非地带性。2000-2014年期间流域内植被物候期格局与前人研究的2010年植被物候格局特征基本一致[11], 部分区域表现出差异, 其主要原因是本研究植被物候期空间格局是15年植被物候期的平均值, 而2010年植被物候只反映了本年度的植被物候空间格局特征, 年内气候因子的不确定性和土地利用方式的变化等都会改变该年度的植被物候, 表现出与历年均值的差异。
2000-2014年期间青海湖流域植被历年返青期均值整体变化趋势并不明显, 与Shen等[26]采用多源遥感影像数据、物候模型得出的2000-2011年期间整个青藏高原平均返青期变化趋势并不显著的结论一致, 但与前人基于气象站观测的植被物候数据得出的青海湖流域乃至整个青藏高原植被返青期的研究结果迥异[11, 16, 27, 28], 其原因主要由于植被物候及其变化是多个环境因子综合作用的结果[24], 而气象站及其周边的环境因子并不能代表整个青海湖流域的环境状况。因此, 气象站选取样区(点)观测的植被物候年际动态变化并不能代表不同空间尺度下植被物候的整体动态变化特征。近15年来流域内历年平均枯黄期呈现提前趋势, 生长季缩短, 与丁明军等[25]基于AVHRR 数据分析的1999-2009年青藏高原植被物候变化趋势不同。研究结论的差异可能与数据源、时间[29]和空间尺度差异有关, 需要进一步的研究证实。
长时间序列下植被活动的减弱将导致植被退化, 可通过植被指数或物候指标的动态变化反映出来[30]。当前人们越来越关注气候变化对植物物候的影响, 忽略了全球变化中植被退化与植被物候之间的关系。青海湖流域作为青海省重要的牧业生产区, 草地退化是流域生态环境恶化的主要表现形式之一[31]。草地退化会对植被物候产生直接或间接的负面影响, 尤其是受土壤水分限制的植被生态系统[13, 32]。草地退化后植被高度的降低、地上生物量的减少会通过改变近地表小气候间接影响植被物候[33]。Chen等[14]认为草地退化后植被覆盖度的降低将通过增加地表反照率降低地表温度, 最终表现为植被返青延迟, 本研究中植被退化与返青期呈现正相关的研究与此结论相悖。其原因主要由于Chen等[14]仅从植被退化会导致近地表小气候的变化推理了植被退化与返青的关系, 忽略了不同草地退化梯度下土壤养分含量的变化[34]也会改变植被物候。通常在高寒草地生态系统中, 随着植被退化程度加重, 土壤中有机碳、全磷、速效氮等养分含量降低[35, 36]。相关研究证实土壤中氮、磷含量的增加将导致莎草科、禾本科、杂类草主要代表植物的返青期和枯黄期推迟[37], 即植被退化将导致植被返青期和枯黄期提前, 与本研究中植被退化将导致返青期延迟的结论一致。青海湖流域内植被退化与枯黄期的关系较复杂, 可能与在研究中没有考虑到流域内季节性轮牧和家畜牧食行为会直接、间接的影响植被物候有关[38, 39]。在适宜的牧草生育阶段进行合理的放牧干扰会直接加速营养物质循环、改善冠层辐射状况、促进资源再分配[40], 给牧草提供良好的补偿或超补偿生长环境, 直接加快牧草再生长速度[41], 延迟其进入枯黄期的时间[42]。另一方面, 放牧干扰通过控制地表枯落物减缓地表径流, 促进降水渗透、减少土壤水分蒸散量, 最终改善近地表土壤微环境, 从而影响植被物候期[43], 该部分内容将是下一步有待研究解决的关键问题。通常情况下, 植被生长季延长促进植物生长发育, 与生物量、净生态系统生产力的关系表现为正相关, 然而在特定的区域环境条件下也存在植被生长季与生态系统生产力呈现负相关的情况[44]。宋春桥等[8]通过研究得出藏北高原地区高寒草甸、高寒草原、高寒荒漠草原植被生长季内最大EVI都呈现降低趋势, 生长季呈现延长的现象, 该结论验证了本研究得出的青海湖流域植被退化趋势下, 生长季趋于延长的结果。植被返青期和枯黄期共同决定植被生长季的长度, 因此, 流域内植被退化与生长季之间呈现负相关的原因仍然与植被退化通过改变近地表小气候、土壤养分含量等影响了植被返青期和枯黄期有关。
植被退化不仅表现为植被生物量、覆盖度的降低, 植被群落优势物种的改变也是植被退化的表现形式之一[45]。长期以来, 过度的放牧利用致使流域内退化草地植物群落结构发生变化, 物种组成产生退化演替[46]。高寒草地生态系统内不同的植物物种之间不仅存在物候特征的差异, 其对气候变化的响应也不同[47]。流域内植被动态变化与物候之间关系异常的区域是否与植被退化演替有关仍需进一步的研究证明。
本研究利用青海湖流域2000-2014年MODIS 16 d EVI合成数据, 选用阈值法提取了青海湖流域植被物候空间格局及时空动态变化, 并在分析整个流域尺度下植被平均物候期年际变化与年均温度、降水量之间关系的基础上, 进一步探究了植被退化与物候时空动态变化之间的关系, 主要结论如下:
1)2000-2014年期间, 青海湖流域内植被陆续进入返青阶段集中在4月中旬-6月中旬, 历时约60 d。植被返青时间在流域整体空间上呈现由东南向西北延迟的水平地带性变化格局, 多高山分布区域的植被进入返青期时间表现出垂直非地带性。植被陆续进入枯黄阶段在8月中旬-10月中旬, 历时约60 d, 进入枯黄期时间在空间格局上与返青空间格局扩展趋势相反。流域内植被平均生长季为124.6 d, 生长季长度集中分布在100~150 d之间, 空间格局与返青期、枯黄期空间格局一致。
2)研究期内的流域植被返青期均值变化趋势并不明显, 但在地理空间格局上却表现出变化趋势的明显差异性; 虽然流域内植被枯黄期均值提前6.4 d, 速率为4.2 d/10年, 但在空间上枯黄期变化趋势存在异质性。流域西北部、布哈河子流域是枯黄期提前趋势最明显的区域; 流域内植被生长季均值缩短8.9 d, 速率为5.9 d/10年, 空间上大面积植被生长季整体缩短趋势较明显。青海湖北岸、东岸退耕还林、还草区是生长季延长趋势最明显的区域, 延长时间超过40 d以上。
3)气候变化趋势下, 虽然青海湖流域植被平均物候期变化与温度、降水量具有一定的相关性, 但并不能完全解释流域植被物候时空动态变化空间异质性的原因。研究表明植被退化趋势对植被物候的时空动态变化也有影响, 即植被年际生长趋于茂盛, 返青期呈现延迟趋势; 反之, 植被年际生长退化趋势, 返青期呈现提前倾向。枯黄期与植被年际生长状态的关系较复杂, 整体上未表现出较明显的关系。植被年际生长动态变化趋势与生长季呈现强负相关, 即植被退化趋势的区域, 植被生长季趋于延长, 反之亦然。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [3] |
|
| [6] |
|
| [8] |
|
| [11] |
|
| [16] |
|
| [17] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [27] |
|
| [28] |
|
| [31] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [41] |
|
| [46] |
|
| [47] |
|