

{kind=link}
{kind=link}
{kind=link}
伞形科参棕亚科及牵环花亚科气孔的结构及其分类学价值
[王超群1, **  , 郭诗琦
, 郭诗琦1, ** , 刘玫1, * , 茹剑1 , 王靖茹1 , 程薪宇2 ]
, 郭诗琦, 刘玫, 茹剑|
|


作者简介:王超群(1994-),女,甘肃张掖人,本科。E-mail:wangchaoqun_723@163.com。郭诗琦(1994-),女,山东沂水人,本科。E-mail:guosq17@163.com。
本文首次深入研究了伞形科参棕亚科和牵环花亚科及相关类群34属(参棕亚科9属,牵环花亚科21属)48种植物叶片(茎或果实)的气孔结构,结果显示有3种类型气孔:无规则型,不等型及平列型。参棕亚科的气孔多为平列型,少为无规则型,而牵环花亚科多为无规则型,少为不等型( Gymnophyton除外)。 Bowlesia分支的细胞壁多为深波状,其他分支的细胞壁多平直。气孔结构的研究为分子系统学建立的参棕亚科和牵环花亚科,以及 Bowlesia分支提供了形态学依据,并支持分子系统学将 Apiopetalum, Mackinlaya及 Stilbocarpa从五加科分别移入参棕亚科和牵环花亚科,将 Homalosciadium, Platysace, Hermas及 Klotzschia移出两亚科。
The stomatal structures of the leaves (stems or fruits) of 48 species representing 34 genera of the Apiaceae subfamilies Mackinlayoideae (9 genera) and Azorelloideae (21 genera) and their relative taxa were studied in detail for the first time. Three types of stomata - anomocytic, anisocytic and paracytic - were observed. In Mackinlayoideae, the stomata are mostly paracytic and the minority are anomocytic. In Azorelloideae, stomata are mostly anomocytic and the minority are anisocytic. Cell walls in the Bowlesia clade are deeply waved while they are usually straight in the other clades of Azorelloideae. This stomatal study provides morphological evidence for the two subfamilies and the Bowlesia clade that have been established by molecular research. It also provides evidence to support moving Apiopetalum, Mackinlaya and Stilbocarpa from Araliaceae to Mackinlayoideae and Azorelloideae, and against placing Homalosciadium, Platysace, Hermas and Klotzschia in either of these subfamilies.
伞形科(Apiaceae)包括天胡荽亚科(Hydrocotyloideae), 变豆菜亚科(Saniculoideae)及芹亚科(Apioideae)[1, 2]。Drude[1]主要根据果实结构将天胡荽亚科(果实具木质内果皮, 没有心皮柄, 油管无或只存在于主脊中)分为两个族, Hydrocotyleae(分果两侧压扁)及Mulineae(分果背腹压扁)。天胡荽亚科中国仅有3属(Centella, Dickinsia及Hydrocotyle)[3], 其他类群产于澳大利亚, 阿根廷及巴西等地[2]。20世纪末分子系统学研究表明伞形科, 特别是天胡荽亚科, 不是一个自然的类群[4, 5, 6, 7, 8, 9, 10, 11]。Plunkett[8]指出天胡荽亚科是多元的, 原位于Hydrocotyleae族的Hydrocotyle和Trachymene应放在五加科(Araliaceae), Homalosciadium和Platysace构成一个分支, 为其他伞形科的姊妹群, 而其余的天胡荽亚科类群分别位于两个新亚科, 参棕亚科(Mackinlaloideae)和牵环花亚科(Azorelloideae)。新苏格兰特有属Apiopetalum及生长在所罗门群岛的Mackinlaya属应从五加科移入参棕亚科。近期的分子系统学研究[12]进一步表明参棕亚科(其物种多位于Hydrocotyleae族)包括9属, 分为2个分支:Centella clade(Centella, Micropleura, Pentapeltis, Schoenolaena)和Xanthosia clade(Apiopetalum, Actinotus, Chlaenosciadium, Mackinlaya, Xanthosia); 牵环花亚科(其物种多位于Mulineae族)包括21属, 分为4个分支:Azorella clade(Azorella, Laretia, Mulinum, Dickinsia, Diplaspis, Huanaca, Schizeilema, Spananthe, Stilbocarpa), Asteriscium clade(Asteriscium, Domeykoa, Eremocharis, Gymnophyton, Oschatzia, Pozoa), Bowlesia clade(Bolax, Bowlesia, Dichosciadium, Drusa, Homalocarpus)和Diposis clade(Diposis)。牵环花亚科中的Dickinsia属原位于Hydrocotyleae族, 而Stilbocarpa属原位于五加科。原位于Mulineae族的Klotzschia属是牵环花亚科的姊妹群, 而另外两个属Hermas及Choritaenia靠近芹亚科[12]。
尽管大量研究证明果实形态特征(包括翅的形状, 分泌结构, 内果皮解剖特征及结晶的存在和分布)对伞形科分类是十分重要的[13, 14, 15, 16], 可以用于界定参棕亚科及牵环花亚科[17, 18], 并区分亚科内不同的分支[19], 但也有相当多的研究表明伞形科气孔的结构同样具有重要的分类学价值。芹亚科具有不同类型的气孔, 如无规则型(anomocytic), 不等型(anisocytic), 横列型(diacytic)及平列型(paracytic)[20, 21, 22, 23, 24]。然而迄今未见关于参棕亚科及牵环花亚科物种气孔结构的详细报道。本研究旨在深入研究参棕亚科和牵环花亚科所有属及其相关类群(Hermas, Homalosciadium, Klotzschia, Platysace)的气孔结构, 弄清气孔结构是否可以为分子系统学建立的两个亚科及分支提供形态学依据。
本文所用的研究材料为参棕亚科和牵环花亚科[11, 12]及相关类群(Hermas, Homalosciadium, Klotzschia, Platysace), 共34属48种植物的叶片, 个别材料因未取到叶片而用茎或果实。材料分别取自澳大利亚国家标本馆(CANB-Australian National Herbarium), 澳大利亚国家植物园标本馆(CBG-Herbarium, Australian National Botanical Garden), 美国哈佛大学标本馆(GH-Harverd University Herbaria, America), 中国科学院华南植物园标本馆(IBSC-South China Botanical Garden Herbarium), 南非约堡大学植物及植物生物技术系标本馆(JRAU-Herbarium, Department of Botany and Plant Biotechnology, University of Johannesburg, South Africa), 英国皇家植物园标本馆(K-Herbarium, Royal Botanical Garden, England), 美国密苏里植物园标本馆(MO-Herbarium, Missouri Botanical Garden, America), 美国纽约植物园标本馆(NY-New York Botanical Garden Herbarium, America), 中国科学院植物研究所标本馆(PE-Herbarium, Institute of Botany, CAS), 西澳大利亚保护和土地管理部植物标本馆(PERTH-Western Australian Herbarium, Department of Conservation and Land Management), 摩洛哥科学研究所标本馆(RAB-Herbarium, Institute Scientific, Moroco)及荷兰乌特勒州立大学系统植物研究所标本馆(U-Herbarium, Institute of Systematic Botany, State University of Utrecht)。研究的材料中仅Centella asiatica(积雪草)及Dickinsia hydrocotyloides(马蹄芹)产自中国。物种名称及凭证标本信息见表1。
| 表1 用于研究气孔结构的伞形科参棕亚科和牵环花亚科植物的凭证标本信息及采集地 Table 1 Apiaceae subfamilies Mackinlayoideae and Azorelloideae examined for stomatal structures, together with voucher specimen details and locality |
所有材料浸泡在约90 ℃热水中约15~30 min, 待材料充分吸水膨胀后将材料取出, 根据Zarinkamar[25]的观察(伞形科植物叶片的气孔密度, 下表皮大于上表皮), 撕取植物叶片的下表皮或茎及果实的表皮, 将其放在载玻片上, 并在放盖玻片之前加1~2滴50%甘油水溶液, 用具成像系统(Olympus DP 70)的显微镜(Olympus BX 51)观察气孔结构并照相。每个样品观察100个气孔并统计不同类型气孔数量, 根据照片用Photoshop CS 2.0软件绘制气孔图。
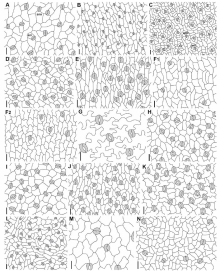
根据前人[20, 21, 22, 23, 26]对气孔类型的描述, 我们确认所研究的物种均有3种类型气孔, 无规则型, 不等型及平列型(图1, 2)。每个物种具有的气孔类型及所占的比例见表2。在研究的物种中, 部分物种仅具一种类型气孔(占观察气孔数目的100%), 为平列型(如Actinotus minor, Centella asiatica及Chlaenosciadium gardneri, 图1A, C, D)或无规则型(如Bowlesia flabilis, Dickinsia hydrocotyloides及Drusa oppositifolia, 图2G, I, M)。一些物种具两种类型气孔:多数无规则型(80%~90%), 少数为不等型(10%~20%)(如Asteriscium chilense, Azorella compacta及Oschatzia saxifraga, 图2A, C, S); 多数无规则型(70%), 少数为平列型(30%)(Hermas quercifolia, 图2Q); 或多数为平列型(70%~90%), 少数为无规则型(10%~30%)(如Apiopetalum glabratum及Micropleura flabellifolia, 图1B, I)。
 | 图1 伞形科参棕亚科及其相关类群植物叶表皮气孔结构缩写 Abbreviations: ano (anomocytic)-无规则型气孔, pa (paracytic)-平列型气孔. (A) Actinotus minor, (B) Apiopetalum glabratum, (C) 积雪草Centella asiatica, (D) Chlaenosciadium gardneri, (E) Homalosciadium homalocarpum, (F) Mackinlaya celebica, (G) Mackinlaya macrosciadea, (H) Micropleura renifolia, (I) Pentapeltis peltigera, (J) Platysace maxwelli, (K) Schoenolaena juncea, (L) Xanthosia atkinsoniana. 标尺 Scale bar=20 μ m.Fig.1 The leaf epidermis of Apiaceae subfamily Mackinlayoideae and related genera showing the stomatal types |
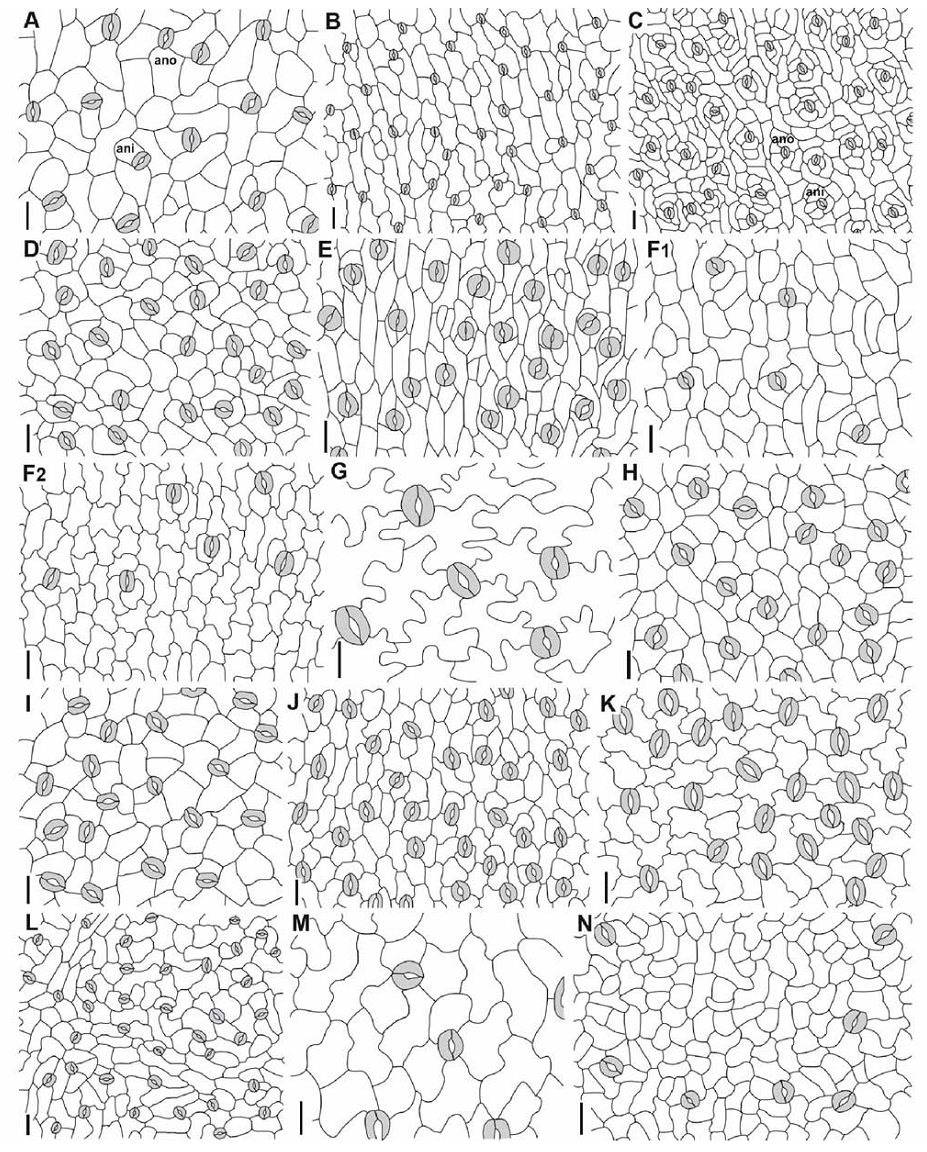 | 图2 伞形科牵环花亚科及其相关类群植物叶(茎或果实)表皮气孔结构示无规则型, 不等型及平列型 Including anomocytic (ano), anisocytic (ani), and paracytic (pa). (A) Asteriscium chilense, (B) A. closii, (C) Azorella compacta, (D) A. crenata, (E) A. sessiliflora, (F1, 2) Bolax gummifera (1 from middle of leaf balde and 2 from leaf margin), (G) Bowlesia flabilis, (H) Dichosciadium ranunculaceum, (I) 马蹄芹Dickinsia hydrocotyloides, (J) Diplaspis nivis, (K) Diposis saniculifolia, (L) Domeykoa amplexicaulis, (M) Drusa oppositifolia, (N) Eremocharis longiramea.标尺 Scale bar=20 μ m.Fig.2 The leaf (stem or fruit) epidermis of Apiaceae subfamily Azorelloideae and related taxa showing the stomatal types |
| 表2 伞形科参棕亚科及牵环花亚科植物叶(茎或果实)表皮气孔类型 Table 2 Summary of stomata of leaf (stem or fruit) epidermis in Apiaceae subfamilies Mackinlayoideae and Azorelloideae |
 | 续图2 伞形科牵环花亚科及其相关类群植物叶(茎或果实)表皮气孔结构示无规则型, 不等型及平列型Including anisocytic (ani), anomocytic (ano), and paracytic (pa). (O) Gymnophyton isatidicarpum (from fruits), (P) G. polycephalum (from stems), (Q) Hermas quercifolia, (R) Homalocarpus dissectus, (S) Huanaca andina, (T) Klotzschia rhizophylla, (U) Laretia acaulis, (V) Mulinum microphyllum, (W) M. spinosum, (X) Oschatzia cuneifolia, (Y) Pozoa coriacea, (Z) Schizeilema fragoseum, (AA) Spananthe paniculata, (BB) Stilbocarpa polaris. 标尺 Scale bar=20 μ m.Continued Fig.2 The leaf (stem or fruit) epidermis of Apiaceae subfamily Azorelloideae and related taxa showing the stomatal types |
早期的研究表明伞形科植物的气孔多为无规则型及不等型, 少见平列型及横列型[21, 22, 23, 26]。所研究的参棕亚科物种仅有或多为平列型气孔, 无规则型气孔的数量较少。在传统的五加科分类系统[27, 28, 29, 30, 31]中, 因Apiopetalum及Mackinlaya两属的花瓣具爪被放在Mackinlayeae族。Eyde和Tseng[32]及Rodrí guez[33]认为Apiopetalum和Mackinlaya兼具五加科(木本及肉质果实)及伞形科(花瓣具爪)的特征, Mackinlaya还具有伞形科特有的两心皮果实及鞘状叶柄基部, 二属可能为介于两科之间的类群。果实形态学研究[18]显示Apiopetalum和Mackinlaya的腹束结构同伞形科的参棕亚科物种的腹束结构十分相似, 支持分子系统学[4, 5, 9, 12, 34]将二属从五加科移入伞形科参棕亚科。我们的研究表明Apiopetalum和Mackinlaya(图1B, F, G)同参棕亚科其他物种(图1A, C, D, H-L)一样, 主要为平列型气孔, 进一步为二属应位于参棕亚科提供了形态学依据。
花及果实形态学研究显示原位于五加科, 生长在澳洲麦加里岛的Stilbocarpa属的形态与伞形科物种较为相似[32, 35, 36]。Mitchell等[37]指出Stilbocarpa同天胡荽亚科的Schizeilema及Azorella两属亲缘关系较近, 建议将其移入天胡荽亚科。分子系统学研究[12]将Stilbocarpa置于牵环花亚科。本研究显示Stilbocarpa的气孔主要为无规则型(图2BB), 同牵环花亚科其他物种(如Dichosciadium ranunculaceum及Dickinsia hydrocotyloides, 图2H, I)的气孔结构极为相似, 支持Stilbocarpa属分类地位的改变。Pozoa, Eremocharis, Domeykoa, Asteriscium及Gymnophyton属被认为是彼此亲缘关系密切的类群[38]。Henwood等[19]指出这些属的染色体基数为x=5。这种较近亲缘关系得到了分子系统学的证实[12]。本研究显示无规则型气孔是这些属的主要类型(Gymnophyton除外, 需要进一步研究此属叶片的气孔)。许多形态学研究[17, 19, 39, 40]揭示Bowlesia, Dichosciadium, Homalocarpus, Drusa及Bolax属均具星状毛, 分子系统学研究表明他们构成一个分支[11, 12]。我们注意到这些属多具有深波状细胞壁(Dichosciadium及Bolax除外)。这种深波状细胞壁还存在于Gymnophyton isatidicarpum的果皮, 但其他研究的物种的细胞壁平直。
Magee等[41]指出产于非洲的Hermas属应位于牵环花亚科, 但分子系统学研究表明此属应靠近芹亚科[12], Hermas的叶片具单列多细胞毛状体, 另一产于巴西的Klotzschia(牵环花亚科的姊妹群)具有长柄的星状毛, 这两属植物的毛状体均与牵环花亚科植物的不同(未发表)。Hermas及Klotzschia多具无规则型气孔(图2Q, T), Hermas具有的平列型气孔仅存在于参棕亚科。无规则型气孔类型普遍存在于变豆菜亚科(Saniculoideae)(如Actinolema及Sanicula)及芹亚科[22, 23, 26], 而平列型气孔在芹亚科很少见[22]。笔者认为Hermas的分类地位需要进一步研究。此外本研究还显示Homalosciadium及Platysace两属的气孔类型(图1E, J)完全不同于参棕亚科, 支持分子系统学将二者移出胡荽亚科[12]。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [3] |
|
| [15] |
|
| [16] |
|