

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
能源橡胶草 GGPPS基因启动子的克隆及瞬时表达研究
[李永梅 , 冯玉杰, 曹新文, 赵李靖, 祝建波, 闫洁
, 冯玉杰, 曹新文, 赵李靖, 祝建波, 闫洁* ]
, 冯玉杰, 曹新文, 赵李靖, 祝建波, 闫洁]
|
|


作者简介:李永梅(1988-),女,山西吕梁人,在读硕士。E-mail:liyongmeishzu@163.com
以多年生宿根型草本植物橡胶草为材料,根据已获得的橡胶草牻牛儿基牻牛儿基焦磷酸合酶(geranylgeranyl pyrophosphate synthase, GGPPS)基因序列设计引物,采用TAIL-PCR扩增到 GGPPS基因5'上游大小为1131 bp的序列。采用PlantCare和PLACE软件分析,表明该序列不仅具有启动子的基本元件,同时还有与多个器官特异表达元件以及与胁迫相关的顺式作用元件,命名为pTkGGPPS(GenBank:KT901796)。将该序列代替pCAMBIA1304质粒上的CaMV 35S启动子序列,以 GFP基因作为报告基因,构建pCAMBIA1304-pTkGGPPS-GFP植物表达载体,利用农杆菌介导法转化洋葱表皮细胞,结果表明,该启动子能够驱动 GFP基因表达,具有一定的活性。 TkGGPPS基因启动子克隆及瞬时表达为橡胶草中橡胶合成及组织特异性研究奠定了基础。
Based on the known sequence of GGPPS, which encodes geranylgeranyl pyrophosphate synthase, the GGPPS promoter sequence was amplified from the root of the perennial herb Taraxacum kok-saghyz by thermal asymmetric interlaced PCR (TAIL-PCR) using nested specific primers. Analyses of the full-length 1131-bp fragment by PlantCare and PLACE software showed that the promoter sequence contained basic cis-acting elements, multiple organ-specific expression elements, and a number of stress-related cis-acting elements. The promoter sequence was designated as pTkGGPPS (GenBank: KT901796). The plant expression vector pCAMBIA1304-pTkGGPPS-GFP was constructed using this sequence to replace the CaMV 35S promoter sequence in the plasmid of pCAMBIA1304 with GFP as the reporter gene. The construct was transformed into onion epidermal cells using Agrobacterium tumefaciens, and the promoter successfully drove expression of the GFP gene. The cloning and transient expression of the GGPPS gene promoter from T. kok-saghyz provides a reference for further studies on the mechanisms and tissue-specificity of rubber synthesis.
启动子是位于结构基因5'端上游的DNA序列, 能够活化RNA聚合酶, 使之与模板DNA准确结合并具有转录起始特异性。启动子(promoters)就像“ 开关” , 决定转录的方向和效率, 控制基因表达的起始时间、空间和表达程度。目前, 大多数转基因植物所用的启动子是组成型启动子(如烟草花叶病毒35S启动子)。组成型启动子是基因工程中应用最早, 且应用最为广泛的一类启动子。它能够在组织中非特异性和高效表达, 外源基因的高效表达, 能够满足人类的需求; 但是, 外源基因的非特异性表达和异源基因的高效表达, 打破植物原有的代谢平衡, 影响植物的正常生长[1]。为了减少外源基因在组织中非特异性和高效表达所带来的不利影响, 寻求组织特异性启动子就显得尤为重要。
橡胶草(Taraxacum kok-saghyz)是一种多年生的产胶草本植物, 作为产胶代谢机理研究的生物反应器, 对橡胶草产胶相关功能基因的研究尤为重要。牻牛儿基牻牛儿基焦磷酸合酶是产胶代谢途径的一个上游基因, 它能够催化烯丙基焦磷酸合成GGPP。橡胶分子合成起始是一个十分缓慢的过程, 是橡胶生物合成的关键限速步骤; 增加橡胶产量的遗传控制战略应集中在橡胶分子合成的起始上[2]。有研究者报道, 在橡胶体外合成试验中, 二甲基烯丙基焦磷酸(dimethylallyl diphosphate, DMAPP)、香叶基焦磷酸(geranyl diphosphate, GPP)、法呢基焦磷(farnesyl diphosphate, FPP)和牻牛儿基牻牛儿基焦磷酸(geranylgeranyl pyrophosphate, GGPP)都可作为橡胶分子合成的前体物质且各起始物都以一个恒定的速度启动橡胶的合成。但是, 随着起始物碳链的增长, 橡胶合成速度也增加(起始物链长效应), 即DMAPP(C5)< GPP(C10)< FPP(C15)< GGPP(C20)。Cornish等[3]在研究不同烯丙基焦磷酸对银胶菊(Parthenium hysterophorus)橡胶生物合成的影响时, 也发现了起始物链长效应; 另外, 也有学者报道, 多年生的橡胶草根中含20%以上的橡胶(多年生的干重)[4]。综上所述, 研究GGPPS基因的特异性启动子对我国转基因橡胶草基因工程建立良好的技术平台具有重要的意义。
因此, 本研究通过热不对称性PCR(thermal asymmetric interlaced PCR, TAIL-PCR)技术, 克隆橡胶草中GGPPS基因的特异性启动子, 并对其进行活性研究; 根据启动子所含有的顺式调控元件, 预测该启动子的功能, 为提高橡胶草橡胶合成奠定理论基础。
2014年9月, 橡胶草采自新疆石河子市北泉镇附近。
1.2.1 橡胶草TkGGPPS基因启动子的克隆 根据已克隆的橡胶草TkGGPPS基因序列设计3个嵌套的特异性引物:sp1、sp2、sp3, 5个短的简并AD引物:AD1、AD2、AD3、AD4和AD5(表1)。用CTAB法提取橡胶草叶片基因组DNA, 3个特异引物和5个短的AD引物组合进行TAIL-PCR扩增。反应体系:模板(基因组DNA)1 μ L, 2.5 mmol/L dNTPs 2 μ L, 引物各0.5 μ L, 10× LA-Taq buffer 2.5 μ L, LA-Taq DNA polymerase(TaKaRa公司)0.5 μ L, 用ddH2O补足至25 μ L。第2轮和第3轮PCR分别以上一轮PCR的产物稀释100倍作为模板, 反应体系同上。扩增程序参考文献[5-9], 具体扩增程序见表1。用1.0%的琼脂糖凝胶电泳检测3轮PCR扩增产物, 用琼脂糖凝胶DNA回收试剂盒(Tiangen公司)回收第3轮PCR中单一条带, 连接到pMD19-T载体(TaKaRa公司)上, 连接产物转化Escherichia coli Top10感受态细胞, 阳性单克隆交由北京华大基因股份有限公司进行测序。
| 表1 TAIL-PCR的扩增程序 Table 1 Amplification program of TAIL-PCR |
根据TAIL-PCR所得序列设计特异引物:pTkGGPPS-F和pTkGGPPS-R。以橡胶草基因组DNA为模板, 以pTkGGPPS-F和pTkGGPPS-R为上下游引物, 扩增橡胶草TkGGPPS基因启动子序列, 扩增程序为94 ℃ 5 min; (94 ℃ 30 s, 57 ℃ 30 s, 72 ℃ 90 s)30个循环; 72 ℃ 10 min。用1.0%的琼脂糖凝胶电泳检测, 用DNA琼脂糖凝胶回收试剂盒回收PCR产物。将纯化产物与pMD19-T载体连接, 连接产物转化E.coli Top10感受态细胞, 将阳性单克隆进行测序(表2)。
| 表2 PCR扩增使用的引物 Table 2 Primers used in PCR amplification |
1.2.2 pCAMBIA1304-pTkGGPPS-GFP植物表达载体构建 用EcoRⅠ 与NcoⅠ (TaKaRa公司)双酶切pMD19-Tector-pTkGGPPS重组质粒与pCAMBIA1304质粒, 分别回收小片段和大片段。将pTkGGPPS小片段与pCAMBIA1304载体大片段用T4-DNA连接酶(TaKaRa公司), 16 ℃连接过夜, 将重组质粒转化E.coli Top10感受态细胞, 经卡那霉素(Kan)筛选, 进行PCR和双酶切鉴定, 经鉴定正确的阳性重组子, 用电击转化法转入根癌农杆菌GV3101, 获得阳性转化子。命名为pCAMBIA1304-pTkGGPPS-GFP/GV3101。
1.2.3 根癌农杆菌介导的方法转化洋葱表皮细胞 根癌农杆菌介导转化洋葱表皮细胞的方法, 参照文献[10-12]。在超净工作台中, 将洋葱表皮用手术刀划成1 cm× 1 cm的方块, 用尖头镊子从一角轻轻撕下, 于MS培养基中预培养24 h, 农杆菌28 ℃振荡培养至OD260≈ 0.6, 收集菌液, 用等体积MS液体培养基重悬, 将洋葱表皮在重悬液中侵染30 min, 用滤纸吸去多余的菌液后, 重新接种到铺有一层滤纸的培养皿中, 共培养24 h后, 激光共聚焦显微镜检测荧光信号。
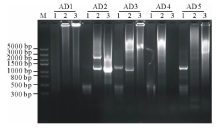
经电泳检测3轮PCR产物, AD2简并引物和3条特异引物组合在第3轮PCR产物中有清晰的扩增条带(图1), 经测序, 该片段长1185 bp。
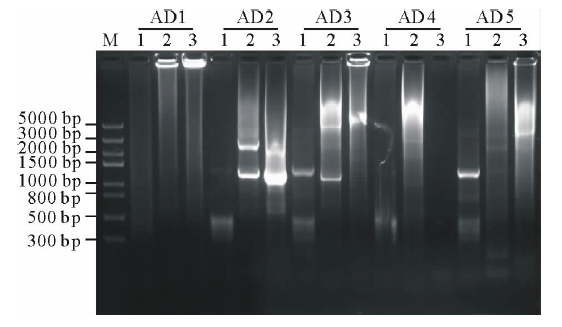 | 图1 TAIL-PCR扩增产物的电泳分析 M: Trans5K DNA Marker(TaRaKa公司); 1: 第1轮PCR产物; 2: 第2轮PCR产物; 3: 第3轮PCR产物; AD1、AD2、AD3、AD4和AD5: 在TAIL-PCR中, 与特异引物组合的5个简并引物。Fig.1 Analysis of TAIL-PCR products by agarose electrophoresis M: Trans5K DNA Marker; 1: Products of primary PCR; 2: Products of the secondary PCR; 3: Products of the tertiary PCR; AD1, AD2, AD3, AD4 and AD5: Five arbitrary degenerate primers in the TAIL-PCR. |
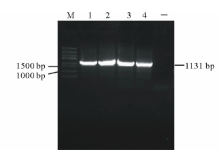
以橡胶草基因组DNA为模板, 以TAIL-PCR的测序结果设计特异引物pTkGGPPS-F和pTkGGPPS-R, 进行PCR扩增, 电泳检测发现单一的特异条带(图2), 与推测的理论值相符。测序发现该产物为橡胶草TkGGPPS基因的5'侧翼序列, 全长为1131 bp。命名为pTkGGPPS(GenBank登录号:KT901796)。
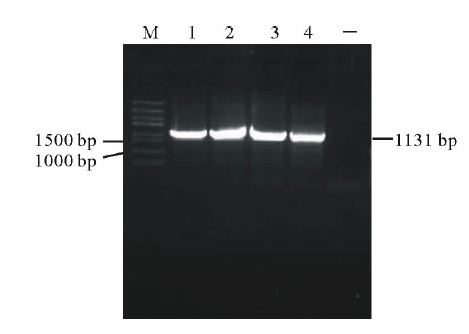 | 图2 橡胶草pTkGGPPS启动子序列的电泳分析Fig.2 Analysis of TkGGPPS promoter on agarose electrophoresis M: Trans5K DNA Marker; 1-4: TkGGPPS启动子的PCR产物PCR Product of TkGGPPS promoter; -: 阴性Negative. |
经启动子分析软件PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)和PLACE(https://sogo.dna.affrc.go.jp/cgi-bin/sogo.cgi?lang=en& pj=640& action=page& page=newplace)分析扩增得到pTkGGPPS启动子序列(图3)。分析表明, TkGGPPS基因启动子序列不仅具有植物启动子基本的顺式作用元件(CAAT-box和TATA-box), 还有与光、植物激素、逆境胁迫等相关的顺式作用元件, 比如I-box(光调节响应的顺式作用元件(cis-acting elements involved in light-regulated responsiveness)、GARE(赤霉素应答相关的元件GA-responsive element)、GT-1 box(病原菌和盐胁迫响应的顺式作用元件cis-acting elements Plays a role in pathogen- and salt- induced responsiveness)、W-box(水杨酸诱导响应的WRKY结合位点WRKY binding wite involved in salicylic acid-induced responsiveness)、ACGT-motif(黄化诱导相关的顺式作用元件cis-acting elements involved in etiolation-induced responsiveness)和LTRE(低温应答元件low temperature responsive element)等。各个元件所具有的生物学功能如表3所示。
 | 图3 橡胶草GGPPS启动子序列Fig.3 The promoter sequence of GGPPS in T. kok-saghyz |
| 表3 TkGGPPS启动子顺式作用元件的生物学功能分析 Table 3 Biological function analysis of cis-acting element of TkGGPPS promoter |
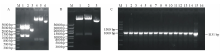
用EcoRⅠ 与NcoⅠ 双酶切pMD19-Tector-pTkGGPPS重组质粒和pCAMBIA1304质粒, 结果如图4A所示。分别回收目的小片段和载体大片段, 将pTkGGPPS小片段与pCAMBIA1304载体大片段用T4-DNA连接酶, 16 ℃, 连接2 d, 将重组质粒转化E.coli Top10感受态细胞, 涂布到含Kan抗性的LB筛选平板上, 过夜培养。经PCR检测和双酶切双重鉴定, 结果如图4B所示。用EcoRⅠ 与NcoⅠ 双酶切, 有一个约1131 bp的目的片段和一个大片段, 说明 pCAMBIA1304-pTkGGPPS-GFP载体构建成功。将鉴定成功的重组质粒, 转化根癌农杆菌GV3101, PCR检测结果如图4C所示。
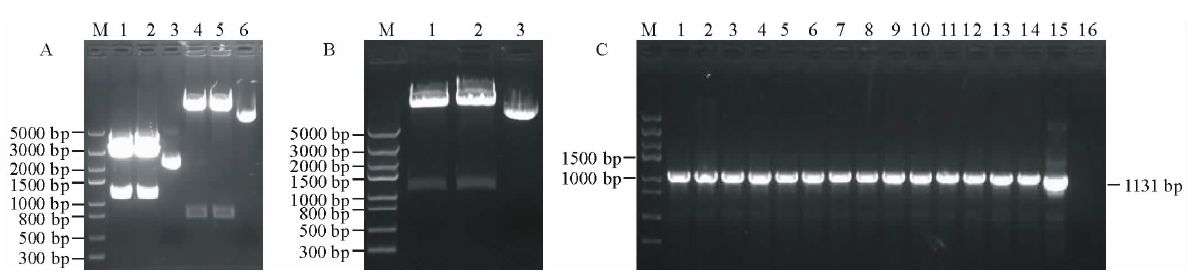 | 图4 pCAMBIA1304-pTkGGPPS-GFP/GV3101植物表达载体的构建 A: pMD19-T-pTkGGPPS质粒和pCAMBIA1304质粒双酶切电泳图 (1~2: pMD19-T-pTkGGPPS质粒双酶切结果; 3: 阳性质粒对; 4~5: pCAMBIA1304质粒双酶切结果; 6: 阳性质粒对照); B: pCAMBIA1304-pTkGGPPS-GFP质粒双酶切电泳图 (1~2: pCAMBIA1304-pTkGGPPS-GFP质粒双酶切结果; 3: 阳性质粒对照); C: pCAMBIA1304-pTkGGPPS-GFP/GV3101菌液PCR鉴定电泳图 (1~14: pCAMBIA1304-pTkGGPPS-GFP/GV3101; 15: 阳性对照; 16: 阴性对照)。Fig.4 Construction of prokaryotic expression vector of pCAMBIA1304-pTkGGPPS-GFP/GV3101 M: Trans5K DNA Marker; A: The electrophoregram of pMD19-T-pTkGGPPS plamid digested (1-2: pMD19-T-pTkGGPPS plasmid through double enzyme cut; 3: The positive plamid; 4-5: pCAMBIA1304 plasmid through double enzyme cut; 6: The positive plamid); B: The electrophoregram of pCAMBIA1304-pTkGGPPS-GFP plamid digested (1-2: pCAMBIA1304-pTkGGPPS-GFP plasmid through double enzyme cut; 3: The positive plamid); C: PCR electrophoregram of pCAMBIA1304-pTkGGPPS-GFP/GV3101(1-14: pCAMBIA1304-pTkGGPPS-GFP/GV3101; 15: Positive control; 16: Negative control). |
将经PCR鉴定成功的阳性单克隆, 利用根癌农杆菌介导的方法, 对洋葱表皮细胞进行瞬时转化(图5B), 经质量分数为40%蔗糖溶液浸泡30 min发生质壁分离(图5C)。结果表明, TkGGPPS基因的启动子能够驱动GFP基因的表达, 表明该启动子具有活性。
 | 图5 pTkGGPPS-GFP融合蛋白的活性检测 A: 正常洋葱表皮细胞; B: 转染pTkGGPPS-GFP融合基因的洋葱表皮细胞; C: 转染pTkGGPPS-GFP融合基因, 经质量分数为40%蔗糖溶液质壁分离后的洋葱表皮细胞。Fig.5 Activity detection of pTkGGPPS-GFP fusion protein A: CK; B: Onion epidermal cells of transfectional pTkGGPPS-GFP fusion gene; C: transfectional pTkGGPPS-GFP fusion gene, after a 40% sucrose solution plasmolysis onion epidermal cells. |
在分子生物学的研究中, 基因克隆和分子杂交探针的制备等操作常需分离与已知DNA序列相近的未知序列; 而TAIL-PCR技术作为一种用来分离已知序列旁侧的未知序列的一种方法, 该技术简单易行, 反应高效灵敏, 产物的特异性高, 重复性好, 能够在较短的时间内获得目的片段, 已经成为分子生物学研究中一项有用的技术。TAIL-PCR是Liu等[8]1995年设计并对其进行不断发展完善。Liu等[6, 8]用这种方法成功分离了YAC、P1和BAC载体的插入序列和拟南芥T-DNA插入的侧翼序列。梁成真等[13]利用优化TAIL-PCR方法, 成功地克隆棉花抗逆相关转录因子编码基因。仇艳光等[14]对TAIL-PCR进行改良, 并成功地分离小麦基因启动子。启动子作为转录调控中心一直是研究的热点, 获得具有功能的启动子是研究的基础。本研究随机选取5对简并引物, 利用改良的TAIL-PCR方法, 成功地扩增得到橡胶草GGPPS基因5'侧翼序列, 其中, 引物AD2的特异性较高。
通过分析启动子序列, 发现该启动子除含有植物启动子所具有CAAT-box和TATA-box等基本的顺式作用元件外, 还包括I-box、GARE、W-box、GT-1 box和LTRE等多个与光、植物激素、逆境胁迫等相关的顺式作用元件。最为重要的是, 该启动子具有根、叶肉和胚乳特异性的顺式作用元件, 这与GGPPS基因主要在根、叶和种子的表达的结果相一致[15]。另外, Hefner等[16]研究表明, 经茉莉酸甲酯诱导后的红豆杉细胞GGPPS的RNA水平要明显高于非诱导的对照细胞; 化文平等[17]研究表明, SmGGPPS3基因在丹参不同发育时期不同器官中表达差异显著, 同时受茉莉酸甲酯和病原菌的诱导。这说明该启动子区域应该有茉莉酸甲酯响应的顺式作用元件或相关的转录因子, 但是该启动子的生物信息学的预测并没有发现, 原因需要做进一步的研究。
以绿色荧光蛋白为报告基因, 利用根癌农杆菌介导的瞬时转化法对该启动子进行活性检测, 结果表明本研究克隆获得的启动子具有转录活性。GGPPS基因在橡胶草中表达具有组织特异性, 该启动子是否是组织特异性启动子, 还需进一步验证。
本研究利用TALL-PCR成功克隆到橡胶草GGPPS基因5'侧翼序列, 大小为1131 bp。利用启动子分析软件分析, 表明该序列不仅含有植物启动子基本的顺式作用元件, 还包含相应逆境、植物激素等顺式作用元件。以绿色荧光蛋白为报告基因, 构建融合表达载体, 瞬时转化洋葱表皮细胞, 发现该启动子能够驱动GFP基因的表达, 说明该启动子具有生物活性。橡胶草中橡胶主要在根部合成, 利用该启动子驱动橡胶合成相关功能基因在橡胶草根中过表达, 以提高橡胶合成量, 为橡胶草产业化生产橡胶提供有力的工具。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [1] |
|
| [7] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [17] |
|