{kind=link}
不同玉米自交系耐深播性评价及遗传多样性分
[彭云玲*  , 赵小强, 闫慧萍, 武金欢]
, 赵小强, 闫慧萍, 武金欢]
, 赵小强, 闫慧萍, 武金欢]
|
|


作者简介:彭云玲(1978-),女,河南南阳人,副教授,博士 E-mail:pengyunlingpyl@163.com
为了筛选玉米耐深播鉴定指标,探讨耐深播综合评价方法并挖掘耐深播种质类群,本研究采用PVC管盆栽试验,在3,15和20 cm 三种深播处理下,测定各自交系的出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长,利用隶属函数法综合评价51份玉米自交系的耐深播性,并用70对SSR标记对其遗传多样性进行了分析。结果表明,随着播种深度的增加,各自交系的中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和大体呈增加趋势,而出苗率降低,苗长及根长先下降后增加。不同播深条件下,玉米自交系幼苗性状的方差分析表明,出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长与自交系间的差异极显著;同时5个性状(出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长)与播深以及5个性状与自交系和播深间相互作用的差异极显著;同时根长与播深之间的差异达到显著水平,但是根长与自交系和播深间相互作用的差异不显著。另外,不同播深处理下各性状的相关分析表明,在3 cm播深条件下的出苗率与中胚轴、中胚轴和胚芽鞘之和、苗长3个表型性状呈正相关;而在15和20 cm播深下,出苗率与中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长都极显著正相关。利用隶属函数法筛选出了6份强耐深播系,11份中等耐深播系。同时,利用70对SSR标记共检测出222个等位基因,平均3.17个,多态性信息量平均为0.579,幅度为0.265~0.801,遗传相似系数幅度为0.496~0.946,并将供试自交系划分成两大优势群或6个亚群,其中四平头亚群(SPT)、兰卡斯特亚群(Lan)和旅大红骨亚群(LRC)的耐深播性较强,出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长表现良好,含有较多的强或中等耐深播系,是重要的耐深播种质类群。
The aim of this study was to establish accurate indices to identify and evaluate the deep-sowing tolerance of a range of inbred lines from the maize germplasm. Maize seeds were planted at three depths (3, 15, and 20 cm) and the following phenotypic traits were evaluated: emergence rate (RAT), mesocotyl length (MES), coleoptile length (COL), mesocotyl and coleoptile total length (MES+COL), seedling length (SDL), and root length (RL). At the same time, simple sequence repeat (SSR) markers were used to evaluate the genetic diversity among 51 maize inbred lines. The results showed that as the sowing depth increased from 3 to 15 or 20 cm, the RAT gradually decreased, SDL and RL first increased and then decreased, and MES, COL, and MES+COL increased. There were strong and significant differences in RAT, MES, COL, MES+COL, SDL, and RL among the maize inbred lines. Five traits (RAT, COL, MES, MES+COL, SDL) were significantly affected by sowing depth and by the interaction between genotype and sowing depth. Root length differed significantly depending on sowing depth, but it was not affected by the interaction between genotype and sowing depth. There were strong and significant positive correlations between RAT at 15- and 20-cm sowing depths and MES, COL, MES+COL, SDL, and RL. Six highly deep-sowing tolerant inbred lines and 11 moderately deep-sowing tolerant inbred lines were identified by the membership function method based on their performance when sown at 15 and 20 cm depths. Seventy SSR markers produced 222 alleles with an average of 3.17 alleles per marker. The polymorphism information content for each marker ranged from 0.265 to 0.801 with an average 0.579. The 51 maize inbred lines were divided into two major groups and six sub-groups in a genetic diversity analysis. Relatively high membership values for RAT, MES, COL, MES+COL, SDL, and RL were obtained for the Sipingtou (SPT), Lancaster (Lan), and Luda Red Cob (LRC) lines, which were highly or moderately tolerant to deep sowing. These results showed that maize lines tolerant to deep sowing can be screened by planting seeds at 15 and 20 cm depths, and that RAT, MES, COL, MES+COL, SDL, and RL are important indices to identify deep-sowing tolerance in maize. We conclude that deep-sowing tolerance can be objectively and accurately evaluated by the membership function method in maize inbred lines, and that SPT, Lan, and LRC are important deep-sowing tolerant germplasm groups.
玉米(Zea mays)作为我国的第一大粮食作物, 其在经济、能源方面发挥着重要作用, 但是其对干旱比较敏感。干旱缺水不仅阻碍玉米生长发育, 干扰生理过程, 而且严重影响籽粒产量[1]。种子萌发阶段是作物能否在干旱条件下完成生育周期的关键时期之一[2], 苗期遇干旱时会使作物出苗能力下降, 保苗率降低, 严重影响田间苗数, 从而导致减产[3]。深播是玉米苗期避旱的重要途径之一, 研究玉米种质资源的耐深播特性对干旱地区玉米生产和耐深播育种具有重要意义。Zhao和Wang[4]从印第安蓝粒玉米中筛选出耐深播自交系368-4。董存吉等[5]利用长根茎特性的印第安蓝粒玉米种质已育成SN系列自交系和杂交种“ 抗42” 。山西农业科学院以自选系太160-2为母本, 耐深播长根茎玉米自选系LR1921为父本, 培育出抗倒、抗旱、高产的旱玉5号玉米品种[6]。作物的耐深播性研究中, 众多学者认为出苗率是耐深播种质鉴定的重要指标, 出苗率越高, 耐深播性越强[7, 8, 9, 10]。赵光武等[11]和马殿荣等[12]认为玉米耐深播的主要原因是其中胚轴显著伸长。Hoshikaw[13]和Zhang等[14]报道中胚轴长度是决定玉米耐深播能力的主要因素, 而在小麦(Triticum aestivum)的研究中人们发现胚芽鞘长是决定小麦耐深播性的主要因素[15]。张磊等[16]研究表明, 中胚轴与胚芽鞘协同作用把玉米幼苗送出地表。吴海燕等[17]报道玉米的出苗率、苗长、中胚轴长、胚芽鞘长与播深显著相关。作物的耐深播性是一个非常复杂的遗传性状, 易受环境的影响[18], 因此筛选出可靠的耐深播性鉴定指标, 并且科学地进行耐深播性评价, 对耐深播种质筛选具有重要意义, 是耐深播育种的前提。分子标记作为一种新兴的技术, 目前主要用于遗传多样性分析、分子遗传图谱构建、基因定位及分子标记辅助育种等领域[19, 20], 利用SSR分子标记可以挖掘耐深播种质类群及有利基因, 有效地提高育种目标。目前对玉米的耐深播性虽有报道, 但是对玉米耐深播种质鉴定指标的筛选、耐深播性综合评价及耐深播种质类群发掘罕见报道。因此, 本研究拟筛选出玉米耐深播性鉴定参考指标, 建立玉米自交系耐深播性综合评价方法, 并通过SSR标记研究其自交系的遗传多样性, 挖掘耐深播种质类群, 以期为玉米耐深播种质的筛选与改良提供参考依据, 同时为寻找耐深播有利基因提供重要材料。
本试验选用的51份自交系均由甘肃省种质资源库提供(表1)。选用均匀分布于玉米全基因组的230对SSR标记, 从中筛选出扩增条带清晰稳定、多态性好的70对SSR标记(表6), 这些SSR标记的引物序列均来自玉米基因组数据库(Maize Genome Database)网站(http: //www. MaizeGDB.org), 由上海生工生物工程有限公司合成。
| 表1 供试玉米自交系及其系谱来源 Table 1 Pedigrees or origins of 51 maize inbred lines in this study |
试验于2013年4月采用盆栽法在甘肃农业大学日光温室进行。将蛭石分层装入尼龙网封底的PVC管(高40 cm, 直径15 cm)中, 然后选取饱满一致的自交系种子30粒播于蛭石上, 再分别盖3, 15和20 cm蛭石使管内蛭石高度达40 cm, 每一处理3次重复, 播种前统一配土、装钵、浇水。在室内条件下萌发10 d后统计出苗率(出苗数与播种数的百分数), 每一处理选取整体一致的幼苗6株测定其胚芽鞘长(从胚芽鞘节到胚芽鞘顶端的长度)、中胚轴长(种子到胚芽鞘节之间的长度)、中胚轴与胚芽鞘之和(种子到胚芽鞘顶端的长度)、根长(最长根的长度)、苗长(中胚轴和胚芽鞘接合点以上部分的长度)。借鉴郝德荣等[21]方法, 利用隶属函数法对51份玉米自交系进行耐深播性综合评价, 其公式为:
Uij=(Xij-Xjmin)/(Xjmax-Xjmin) (1)
Uij=1-(Xij-Xjmin)/(Xjmax-Xjmin) (2)
式中, Uij表示i材料j指标的耐深播隶属值; Xij表示i材料j指标的测定值; Xjmin表示所有材料j指标的最小值; Xjmax表示所有材料j指标的最大值。若所测指标与材料的耐深播性呈正相关, 则采用(1)式计算隶属值, 反之则用(2)式。累加各指标的具体隶属值, 并求出平均值后进行比较, 平均值越大, 自交系的耐深播性越强。
采用CTAB法提取玉米基因组DNA[22], 用1%琼脂糖凝胶电泳检测DNA质量, 用德国IMPLEN微量分光光度仪检测DNA浓度。PCR反应在Biometra-T1 PCR仪上进行, 采用10 μ L反应体系, 包括2× Taq PCR Master Mix(Tiangen 公司)5.0 μ L, 1 mmol/L正、反SSR引物各0.3 μ L, ddH2O 3.7 μ L, 50 ng/μ L DNA 0.7 μ L。PCR反应程序为:95 ℃预变性5 min, 1个循环; 94 ℃变性30 s, 50~59 ℃退火30 s, 72 ℃延伸30 s, 共36个循环; 最后72 ℃延伸10 min, 4 ℃结束并保存。PCR扩增产物用8%非变性聚丙烯酰胺凝胶电泳进行分析。
试验数据使用统计软件SPSS 16.0的相关程序进行各性状的方差分析、相关分析和描述统计。SSR扩增产物以0、1统计建立数据库, 在相同迁移率位置上, 有带记为“ 1” , 无带记为“ 0” , 缺失记为“ 2” 。以简单配对参数(simple matching coefficient)估计基因频率; 以NTSYS-pc 2.1计算遗传相似系数(genetic similarity, GS), 按非加权配对法(UPGMA)和SHAN程序聚类分析; SSR位点的多态性信息量按PIC=1-∑ {Invalid MML}计算, 式中, Pi表示i位点的基因频率。
51份自交系分别在3, 15和20 cm播深下播种, 而后对其出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长及根长6个性状进行测量。由表2可知, 随着播深增加, 各自交系的中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和大体呈增加趋势, 而出苗率呈降低趋势, 苗长及根长呈现出先下降后增加的趋势, 这些性状在各自交系间变化幅度不同。在3 cm 播深条件下, 各自交系6个性状平均分别为 87.1%, 1.1 cm, 2.1 cm, 3.2 cm, 18.4 cm, 24.4 cm; 在15 cm播深下, 各自交系6个性状平均分别为31.2%, 3.9 cm, 3.8 cm, 7.7 cm, 11.0 cm和18.6 cm; 在20 cm播深下, 6个性状的平均值分别为16.9%, 4.1 cm, 4.5 cm, 8.7 cm, 14.8 cm和21.1 cm。这些数据表明, 播深明显影响玉米的表型性状, 并且在15和20 cm播深条件下的出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长及根长能为玉米耐深播种质提供有效的筛选。就出苗率而言, 大部分玉米自交系的出苗率都随播深增加而降低, W64A、N192、A654、DM和Va35-2在15和20 cm两种深度下出苗率都较高, 且随播深的增加其出苗率下降幅度仅在25.0%~37.5%之间, 说明播深对这些自交系的出苗率影响较小。另外, 增加播种深度可以促进中胚轴伸长, 但伸长的幅度在各自交系间不同。就胚芽鞘长而言, 大部分玉米自交系的胚芽鞘长有所增加, 并且自交系B64的胚芽鞘长增加最多为2.7 cm。另外, 在3 cm播深条件下各自交系的苗长和根长较长, 随着播深的增加到15 cm条件下苗长和根长下降, 而播深增加到20 cm条件下, 苗长和根长又开始增加, 各自交系在20 cm播深下的苗长是15 cm播深下苗长的0.6~2.3倍, 并且在15和20 cm两种播种深度下, 出苗率高的自交系其苗长也较长。
| 表2 51份玉米自交系在不同播种深度下的性状 Table 2 The seedling traits of 51 maize inbred lines at different sowing depths |
如表3所示, 不同播种深度下, 玉米自交系幼苗相关性状的方差分析结果显示, 出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长及根长6个性状在自交系间达到极显著差异水平, 说明这些性状受自交系本身遗传特性的控制, 因此在耐深播鉴定时对出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长等性状选择是有效的。出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长与播深的差异达到极显著水平(P< 0.01), 而根长与播深的差异达到显著水平(P< 0.05), 说明播深极显著影响自交系的出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长, 显著影响根长, 并且这些性状随着播种深度的增加其变化增大。另外, 出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长在自交系与播深互作间的差异也达到极显著水平, 而根长在自交系与播深互作间的差异没有达到显著水平, 说明不同性状对播深的响应程度不同, 相对于其他5个性状, 根长对播深的响应较差。
| 表3 不同播深处理下玉米自交系幼苗性状的方差分析 Table 3 Variance analysis of seedling traits for maize inbred lines at different sowing depths |
不同播深条件下, 中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长等性状与出苗率的相关分析表明(表4), 在3 cm播深条件下的出苗率与中胚轴长、中胚轴和胚芽鞘之和、苗长3个表型性状呈显著正相关; 而在15和20 cm播深下, 出苗率与中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长都极显著正相关, 说明在这两种播深处理下, 中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长及根长在玉米幼苗出土方面发挥着重要作用, 它们共同作用将玉米幼苗送出地表。同时, 由表4可以看出, 播深与出苗率、苗长和根长呈极显著负相关, 而与中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和呈极显著正相关, 说明播深影响玉米自交系的出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长等性状。
| 表4 不同播深处理下玉米自交系幼苗性状的相关分析 Table 4 Correlation analysis of the seedling traits for maize inbred lines at different sowing depths |
作物的深播特性遗传基础复杂, 并且易受环境影响, 这给作物的耐深播性研究带来了诸多困难, 而准确地筛选耐深播鉴定指标及科学地评价种质的耐深播特性具有重要意义。本研究分别在3, 15和20 cm播深条件下, 利用出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长以及根长6个性状作为耐深播性鉴定指标, 结合隶属函数法进行玉米自交系的耐深播性的综合评价。根据隶属值的大小将51份自交系划分为强耐深播类型(隶属值> 0.55), 中等耐深播类型(隶属值=0.45~0.55)和弱耐深播类型(隶属值< 0.45)。51份玉米自交系耐深播性综合评价结果表明(表5), N192、W64A、DM、Mo17、A654、502优等6份自交系的耐深播性在0.57~0.75之间, 平均0.63, 变异系数为11.0%, 属于耐深播类型。自交系B73、黄C、Pa91、TS109、T58、郑22、430、Va35、Va35-2、TS110、B64等11份材料的耐深播性在0.46~0.55之间, 平均0.50, 变异系数为7.1%, 属于中等耐深播类型。其余34份自交系的耐深播性在0.08~0.44之间, 平均0.28, 变异系数为32.9%, 属于弱耐深播类型。
| 表5 51份玉米自交系耐深播性综合评价 Table 5 Comprehensive evaluation of deep-sowing tolerant ability of 51 maize inbred lines |
为了探讨耐深播种质类群, 选择均匀分布于玉米10条染色体上的扩增条带清晰稳定、多态性丰富的70对SSR标记, 对51份玉米自交系进行遗传多样性分析。所得结果如下(表6), 70对SSR引物在51份自交系间共检测到222个等位基因, 每对引物检测到2~6个, 平均3.17个。每对引物的多态性信息量(PIC)变化范围为0.265~0.801, 平均0.579, 其中引物phi114的PIC最小为0.265, 引物umc1196的PIC最大为0.801。51份自交系间的遗传相似系数变化范围为0.491~0.946, 其中自330与TS66的遗传相似系数最小为0.491, R802A与R802遗传相似系数最大为0.946, 说明自330与TS66的亲缘关系最远, 而R802A与R802的亲缘关系最近。然后, 我们利用UPGMA聚类分析法将51份自交系划分为Ⅰ 和Ⅱ 两大优势群, 共6个亚群(图1)。Ⅰ 群包括A、B、C 3个亚群, 占供试材料的52.9%, Ⅱ 群包括D、E、F 3个亚群, 占供试材料的47.1%。A亚群共10份自交系, 包括黄早四、430、K12、502优、K805、R802A、R802、W182BW、W64A、N192属于四平头亚群(SPT); B亚群包括齐319、178、T58是PB亚群, 共3份自交系; C亚群为兰卡斯特亚群(Lan)共14份材料, 包括Mo17、Va26、自330、01743-2、黄C、ND246、Pa91、Mo22、Mo12、DV、A654、AR234、TS109、H105W; D亚群含有丹340、DM、吉853、WN11、综3、Va35、Va35-2等7份自交系, 属于旅大红骨亚群(LRC); E亚群是Reid亚群, 共8份材料; F亚群是PA亚群共9份材料。
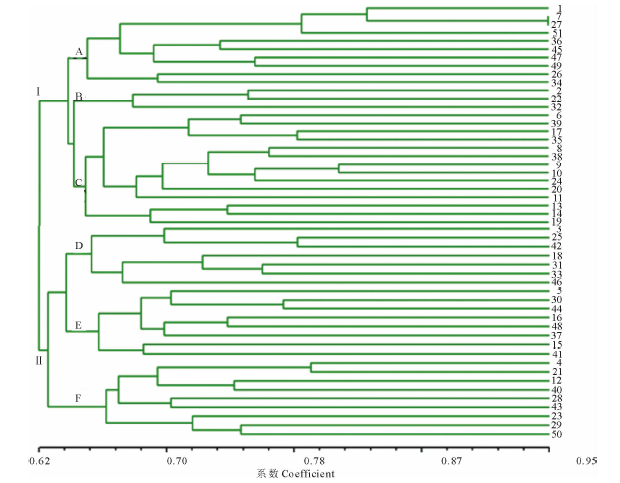 | 图1 51份玉米自交系SSR聚类分析1~51代表材料编号。1~51 is material number.Fig.1 Dendrogram of 51 maize inbred lines clustered by SSR markers |
随后, 分别计算了在15和20 cm播深条件下每个亚群6个表型性状(出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长和根长)的平均值(表7)。结果表明, 在这两种播种深度下, SPT、Lan和LRC亚群的出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、在两种播深下的平均耐深播性分别为0.43, 0.34, 0.35, 0.39, 0.34, 0.33, 说明6个亚群间的耐深播性不同, SPT的耐深播性最强, LRC的次之, Lan的中间, PA的最小。6个亚群内各自交系耐深播性的变异系数分别为42.7%, 45.3%, 42.8%, 49.7%, 38.7%和41.0%, 说明亚群内各自交系的耐深播性差异较大。在SPT亚群内含有3份强耐深播系和1份中等耐深播系, Lan亚群内含有2份强耐深播系和3份中等耐深播系, LRC亚群内含有1份强耐深播系和2份中等耐深播系, 而PB、Reid和PA亚群内只含有1, 2和2份中等耐深播系, 6个亚群内强或中等耐深播系各占供试自交系的7.8%, 9.8%, 5.9%, 2.0%, 3.9%和3.9%, 说明SPT、Lan和LRC 3个亚群在两种播种深度下, 其出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长都表现良好, 耐深播性较强, 含有较多强或中等耐深播系, 是重要的耐深播种质类群。
| 表6 70对SSR引物在51份玉米自交系中检测到的等位基因数及多态性信息量 Table 6 Allele numbers and PIC values for 70 SSR primers found in 51 maize inbred lines |
| 表7 51份玉米自交系UPGMA聚类结果 Table 7 Cluster results using UPGMA based on 70 SSR markers among 51 maize inbred lines |
作物深播时由于土壤阻力增大, 空气减少, 往往表现为营养物质消耗加剧, 出苗时间延长, 出苗率降低, 幼苗活力减弱。王柳英和纪亚君[23]报道播种深度与牧草种子萌发过程中营养物质消耗量显著正相关。Sanusan等[24]报道深播与幼苗干物质积累存在一定的差异。岳丽杰等[25]报道在播种深度为3, 6和9 cm条件下, 随播种深度的增加, 玉米的出苗率逐渐降低, 出苗时间逐渐延迟, 苗长、中胚轴长、苗干重、中胚轴干重逐渐增加, 主根长、根冠比、幼苗整体度逐渐减小。彭云玲等[18]报道在10和15 cm播种深度条件下, 随播种深度的增加, 玉米自交系的根长、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和呈增加趋势, 出苗率、苗长、根数呈下降趋势。为了进一步研究玉米种质的耐深播性, 本研究增加了播种深度, 在播深3(作为对照深度), 15和20 cm条件下进行耐深播性研究, 得到了相似的研究结果。随着播种深度的增加, 玉米自交系中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和大体呈增加趋势, 而出苗率呈下降趋势, 苗长及根长呈现出先下降后增加的趋势, 幼苗变弱且活力降低。另外, 研究中我们发现, 在出苗的过程中有些玉米自交系中胚轴弯曲, 其幼苗顶土能力较弱、出苗率较低, 比如自交系R802A、AR234、ND246等在15和20 cm播深下的出苗率都为0.0%; 而有些玉米自交系的中胚轴直立, 其幼苗顶土能力较强, 出苗率较高, 如自交系W64A、DM、A654在15和20 cm播深下的出苗率分别为80.0%和80.0%, 90.0%和60.0%, 60.0%和50.0%。除此之外, 目前对深播条件下作物体内内源激素、生理生化特性变化的研究较少[26, 27, 28], 更不清楚其在作物耐深播过程中的作用机理, 因此笔者课题组正对播深与激素、生理生化特性的关系进行试验研究, 以便揭示其在作物耐深播过程中的作用机理。
作物的耐深播性是受多种因素综合作用的一个非常复杂的遗传性状, 而表型性状是作物内部生理结构变化的最直接体现, 能客观、直接的反应作物的耐深播能力。Yu和Bai[29]报道胚芽鞘与作物的耐深播性显著相关, 可以作为作物耐深播鉴定的重要指标。Amita等[30]指出小麦胚芽鞘长度是决定幼苗出土的重要动力。Troyer[31]报道随着播深的增加, 中胚轴长度逐渐增加。岳丽杰等[25]报道出苗率与中胚轴长关联度最大。彭云玲等[18]研究表明玉米的出苗率与中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长、根数显著或极显著相关。吴海燕等[17]报道玉米的出苗率、苗长、中胚轴长、胚芽鞘长与播深显著相关。潘前颖等[32]还比较了小麦与玉米的胚芽鞘、中胚轴在幼苗出土中的作用。本研究结果表明, 出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长及根长在自交系间差异显著; 在3, 15和20 cm播深条件下, 除了3 cm播深条件下的出苗率与胚芽鞘长和根长的正相关性没有达到显著以外, 其他情况下出苗率与性状间都呈显著和极显著正相关, 并且播深与出苗率、苗长、根长呈极显著负相关, 与中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和极显著正相关。因此这6个性状同时适合于15和20 cm播深下玉米耐深播性的鉴定。
对作物种质的耐深播性评价, 学者已经提出了一些方法。张磊等[16]在15 cm播深下, 利用根茎长对玉米自交系进行了评价, 将46份自交系划分为强中弱3种类型。张晓龙等[33]在小麦上利用供试材料的平均地中茎长, 做Duncan多重比较, 将供试材料的耐深播性划分为3个等级。彭云玲等[18]在15 cm播深下, 利用出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长、根数等7个性状对45份自交系进行聚类分析, 评价了各材料的耐深播性。虽然利用这些方法也能评价作物的耐深播特性, 但是这些方法只注重了个别指标, 或模糊的将材料聚成不同类型。本研究提出了一种耐深播性评价新方法, 即根据出苗率、胚芽鞘长、中胚轴长、中胚轴与胚芽鞘之和、苗长、根长等耐深播鉴定指标, 利用隶属函数法, 分别计算出材料在15和20 cm两种播深下的隶属值, 然后取其平均值作为材料的耐深播性大小。此方法不仅能综合多种环境、多个耐深播相关性状, 把耐深播性这一复杂性状进行数理统计上的定量表达, 明确每一材料的耐深播性大小, 而且大大地降低了环境对试验结果的影响, 减少了试验误差, 因此利用此方法更能客观、科学地评价玉米的耐深播性。
挖掘耐深播种质类群对玉米耐深播育种及耐深播种质筛选与改良具有重要意义。本研究以6个国内玉米杂种优势群的标准测验种:黄早四(四平头群)、Mo17(兰卡斯特群)、丹340(旅大红骨群)、齐319(PB)、B73(瑞德群)、掖478(PA)为参照, 利用70对SSR标记对51份供试材料的遗传多样性进行研究, 结果表明51份自交系可以划分为两大优势群或6个亚群。分群结果与自交系的系谱基本吻合, 说明此分群结果是科学、准确的。在此基础上, 笔者结合不同播深下供试材料各性状的表现及耐深播综合评价结果表明, 四平头(SPT)、兰卡斯特(Lan)和旅大红骨(LRC)亚群是重要的耐深播种质类群, 群内含有较多的强或中等耐深播系, 其耐深播性较强, 出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长表现良好。根据杂种优势的原理, 可以利用强耐深播系N192、W64A、502优(属SPT), Mo17、A654(属Lan), DM(属LRC)改良群内相应自交系的耐深播性。这些研究结果在理论和实践上都具有一定的意义。根据育种实践, 利用Ⅰ 优势群中的SPT和Lan与Ⅱ 优势群中的LRC组配杂交组合究竟能不能产生强耐深播玉米杂交组合, 还需要进一步的试验研究。
研究结果表明, 在15和20 cm两种播种深度下, 出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长6个性状都可以作为玉米耐深播性鉴定的参考指标; 利用隶属函数法能客观、科学地评价玉米的耐深播性; 在玉米种质中, 四平头(SPT)、兰卡斯特(Lan)和旅大红骨(LRC)亚群是重要的耐深播种质类群, 群内含有较多的强或中等耐深播系, 其耐深播性较强, 出苗率、中胚轴长、胚芽鞘长、中胚轴与胚芽鞘之和、苗长、根长表现良好, 其为玉米耐深播种质筛选及改良提供了理论依据, 为玉米耐深播QTL定位提供了材料。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [2] |
|
| [3] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [21] |
|
| [23] |
|
| [25] |
|
| [27] |
|
| [28] |
|
| [32] |
|
| [33] |
|