

{kind=link}
{kind=link}
牦牛粪沉积下高寒草甸植物群落种间关联研究
[孔杨云 , 于应文
, 于应文* , 侯扶江]
, 于应文, 侯扶江]
|
|


作者简介:孔杨云(1992-),女,甘肃永靖人,在读硕士。E-mail:kongyy15@lzu.edu.cn
植物群落构成和种间关联,反映物种在空间上的分布及相互依存和制约关系。为探究牦牛粪沉积和植被斑块互作下草地植物群落演变趋势及稳定性机制,以青藏高原东北缘高寒草甸禾草/杂类草斑块和嵩草/杂类草斑块为对象,分析牦牛粪沉积下两种斑块中主要植物种的重要值( IV)、Jaccard种间关联指数和Spearman秩相关系数的变化规律。结果表明,1)粪沉积使禾草/杂类草斑块的建群种扁穗冰草的 IV增加和西北针茅的 IV降低,嵩草/杂类草斑块的建群种线叶嵩草的 IV降低;2)禾草/杂类草斑块中,粪沉积(DP)和对照(CK)的正负关联种对比为0.61和0.63,显著联结种对为6.6%和6.4%;嵩草/杂类草斑块中,DP和CK的正负关联种对比为0.90和0.72,显著联结种对为2.8%和14.5%;且二者显著联结的种对多存在于优势种或亚优势种与伴生种之间;3)两种斑块中多数种对间联结性弱,群落处于不稳定阶段;粪沉积使嵩草/杂类草斑块的稳定性增强,利于禾草/杂类草斑块的演变;4)依据优势种对环境的适应方式和主导生态因子,将两种斑块的群落植物种划分为喜阳耐旱植物、喜阴喜湿植物和喜阳喜湿植物三大生态种组。
Plant community composition and interspecific associations reflect inter-dependence and competition between species, influencing their spatial distribution. To explore the evolutionary trends and stability mechanisms of grassland communities influenced by the interaction between yak dung depositions and vegetation we analyzed the importance values ( IV) of different species, Jaccard association indices and Spearman rank correlation coefficients of the main species present in patches of grass/forb and Kobresia/forb swards under yak dung deposition in an alpine meadow on the northeast Qinghai-Tibetan Plateau. The results indicated that: 1) dung deposition (DP) increased the Agropyron cristatum IV and decreased the Stipa krylovii IV in grass/forb patches, while decreasing the Kobresia capillifolia IV for Kobresia/forb parches; 2) in grass/forb patches, the species-pairs ratios of positive and negative associations under DP and control (CK) were 0.61 and 0.63, respectively and their respective associated species-pairs were 6.6% and 6.4%. For Kobresia/forb patches, the species-pairs ratio of positive and negative associations in DP and CK were 0.90 and 0.72, respectively and their respective associated species-pairs were 2.8% in DP and 14.5% in CK, exhibited mainly between the dominant or sub-dominant species and the companion species; 3) the interspecific association of most species-pairs was weak in both vegetation types; yak dung strengthened the stability of Kobresia/forb patches benefiting the generation of grass/forb patches; 4) according to their adaptability to environment and the key ecological factors affecting plants, the main plant species of two patches were divided into three ecological groups: shade-intolerant and drought-tolerant plants, shade-tolerant and drought-intolerant plants, and, shade-intolerant and drought-intolerant plants.
植物种间关联通常由群落生境差异影响物种分布而引起, 是群落基本特征之一[1] 。根据群落内种对对生境的选择及其相互促进和抑制关系, 将群落植物种间关系分为正相关、负相关和不相关[2] 。种对间表现为正相关, 是由于物种具有相近的生物学特性和对环境具有相似的生态适应性以及一个种群的存在可以为另一个种群的生存提供条件; 种对间表现为负相关, 是由于物种生物学特性和对环境喜好的不同及相互竞争[3] 。植物种间关联既是一种对物种存在与否二元数据的定性分析, 也是对物种数量特征数据的定量分析[2] 。研究草地植物种间关联, 有助于深入了解草地植物物种间的相互依存和制约关系及群落结构和演替动态变化。采食、践踏和排泄物沉积作为放牧系统中家畜作用于草地的3种方式, 均是草地植物种间关系和植被构成发生变化的主要影响因素, 在草地植被演替中起重要作用[4]。
青藏高原高寒草甸放牧历史悠久, 拥有世界独特的面积为1.3亿hm2高寒草地及1400万头牦牛, 从而牦牛排泄物沉积量大, 分布广[5] 。由于垫状牛粪沉积对草地植物具有窒息作用, 使粪斑下植物枯萎死亡[6] ; 且粪沉积后期或粪斑破碎后因粪中养分的施肥作用, 促进粪块及其毗邻区域植物生长; 二者均通过影响草地植被构成的改变而引起小尺度植被异质性的形成。因此, 研究牦牛粪沉积下, 高寒草甸植被构成及种间关联, 对于高寒草甸植被演替动态和植物种间共存竞争及群落稳定性机制的揭示具有重要意义[7, 8, 9]。
诸多学者对群落类型[10]和不同管理方式(封育和放牧)[11, 12]及鼠丘密度[13]下, 青藏高原高寒草甸植物群落构成及种间关联进行了分析。研究表明, 高寒草甸植物种间关联与封育年限有关, 短期封育表现为微弱负关联, 5~6年的封育时间无关联[11]; 自由放牧较划区轮牧降低高寒草甸主要植物种间的关联程度和群落稳定性[12]; 线叶嵩草(Kobresia capillifolia)草地群落总体关联性表现为显著负相关且处于不稳定阶段[10]; 高山嵩草(Kobresia pygmaea)草地群落的总体联结性与高原鼠兔有效洞口密度有关, 由低洞口密度(≤ 224 个/hm2)下的正关联转变为高洞口密度(> 224 个/hm2)下的负关联[13]。但家畜排泄物沉积下, 高寒草甸植物构成及种间关系的研究, 限于牦牛粪沉积下草地植物种生态位[14]及植物种子库构成特征[15, 16]分析。由于草地植被异质性和家畜采食的非均匀分布, 使粪斑和草地植被斑块产生叠加效应, 而植被构成对粪沉积的响应因群落植物种构成不同[17]。截至目前, 牦牛粪沉积与草地植被构成的整合研究尚未见报道。
本研究采用Jaccard种间关联度指数及Spearman秩相关方法, 对牦牛粪沉积下青藏高原东北缘高寒草甸禾草/杂类草斑块和嵩草/杂类草斑块植物群落构成和种间关联进行定量分析, 揭示高寒草甸植物物种共存竞争及群落稳定性机制, 为高寒草甸的管理提供理论基础和实践依据。
研究区位于青藏高原东缘的甘肃省天祝县抓喜秀龙乡的高寒草甸上, 地理坐标为E 102° 40'-102° 47', N 37° 11'-37° 14', 海拔2960 m。年均气温-0.1 ℃, 1月均温-18.3 ℃, 7月均温12.7 ℃, > 0 ℃年积温 1380 ℃; 年降水量416 mm, 无绝对无霜期, 仅分冷热两季, 土壤类型为亚高山草甸土。草地植被斑块主要有禾草/杂类草、嵩草/杂类草、珠芽蓼(Polygonum viviparum)斑块等。草地植物种主要有矮嵩草(Kobresia humilis)、线叶嵩草、扁穗冰草(Agropyron cristatum)、西北针茅(Stipa krylovii)、洽草(Koeloria cristata)、早熟禾(Poa annua)、垂穗披碱草(Elymus nutans)、珠芽蓼和球花蒿(Artemisia smithii)等。
2012年5月末, 在研究区多年放牧牦牛的禾草/杂类草斑块与嵩草/杂类草斑块普遍出现的高寒草甸冷季牧场上, 设置面积为0.3~0.5 hm2的样地3块。在设置的3块样地的两类斑块上, 用木桩和彩色绳子分别标记沉积约3个月内的牦牛粪斑及其配对的对照样方各20~30个, 粪斑之间间距10 m以上, 每类斑块共标记60~90个粪斑。各样地设置后仍作为牦牛冬春放牧地, 除每年6-10月休牧外。草地利用率为80%~90%。
在研究区牦牛粪斑大小观测为0.02~0.03 m2的前期基础上, 考虑牛粪对草地植物的影响效应通常约为粪斑大小的2倍[18], 将本研究牦牛粪斑大小设置为0.1 m2, 以客观反映牦牛粪斑对草地植物的作用效应。
依据牦牛粪斑对植物作用效应的前期观测结果, 即牦牛粪沉积对草地植物的作用效应一般在1年多后才表现出来。故在2013年7-8月(牦牛粪沉积约1年多后), 进行草地植物特性的测定。具体植被特征观测时, 以标记的粪斑为中心, 设置0.1 m2的正方形样方, 作为粪斑处理(dung patch, DP); 再在离标记的粪斑周围50~60 cm处草地上, 设置无粪尿排泄物沉积的与各标记牦牛粪斑配对的0.1 m2的正方形对照样方(control, CK)。分别测定各标记粪斑及其配对对照样方内的所有植物种的地上生物量和分种盖度, 地上生物量以干重计。因标记牦牛粪斑的部分破坏等, 用于本研究群落植物种重要值和种间关联分析的数据, 每类斑块为38对。
以分种生物量和盖度数据为基础, 计算两种群落的粪斑及其配对对照区域的植物种重要值。植物种重要值(importance values, IV)计算公式为:IV=(RC+RB)/2。式中:RC和RB分别表示植物种的相对盖度(relative coverage, RC)和相对生物量(relative biomass, RB), 样方中某种的相对盖度(或相对生物量)等于该种盖度(或生物量)除以所有植物种的盖度(或生物量)之和。
JI指数反映的是物种共同出现的百分率。本研究采用Jaccard指数[2](简称JI指数)测定物种间关联程度, 它们以2× 2列关联表为基础计算, 其公式为:JI=a/(a+b+c)。式中:a为所有样方中种A和种B共同出现的样方数, b、c分别为所有样方中种A或种B单独出现的样方数。JI指数满足当a=0时, 取值为0, 表示种A和种B不同时出现在同一样方内, 即种A和种B完全独立或相异; 当a=S(样方数)时, b=c=0, 取值为1, 表示物种A、B同时出现在所有样方中, 即种A与种B绝对关联; 当c=0时, 表示完全关联, 即种B依赖于种A, 种B随种A出现而出现, 而种A可以单独出现。在借鉴张金屯[2]划分方法, 将JI指数划分为以下4个等级, 0.75< JI≤ 1为强联结, 0.5< JI≤ 0.75为次强联结, 0.25< JI≤ 0.5为弱联结, 0≤ JI≤ 0.25为无联结, 以对嵩草/杂类草和禾草/杂类草草地群落主要植物种种间关联进行分析。
以植物种重要值数据为依据, 采用SPSS 16.0中Correlate中的Bivariate模块, 计算两种群落粪斑和对照区域主要植物种间Spearman秩相关系数及显著性检验[19]。
由表1可知, 禾草/杂类草斑块中, DP和CK的优势种和亚优势种均为扁穗冰草、西北针茅和洽草, 粪沉积使扁穗冰草的重要值增加26%, 使西北针茅的重要值降低19%; DP主要伴生种有早熟禾、大籽蒿、矮嵩草、紫花针茅、天蓝韭、阴山扁蓿豆、线叶嵩草、垂穗披碱草、冷蒿、藨草、二裂委陵菜和高山紫菀; CK主要伴生种有矮嵩草、早熟禾、大籽蒿、紫花针茅、天蓝韭、阴山扁蓿豆、二裂委陵菜、赖草、藨草和线叶嵩草; 且DP的线叶嵩草重要值约为CK的2.0倍。嵩草/杂类草斑块中, DP和CK的优势种和亚优势种分别为线叶嵩草和矮嵩草, 粪沉积使线叶嵩草重要值降低10%; DP主要伴生种有紫花针茅、珠芽蓼、垂穗披碱草、苔草、洽草、球花蒿和瑞香狼毒, CK主要伴生种有紫花针茅、珠芽蓼、洽草、高山嵩草、球花蒿、瑞香狼毒、垂穗披碱草、甘肃棘豆和乳白香青; 且斑处垂穗披碱草和洽草的重要值分别约为对照的3.6和0.7倍。说明, 粪沉积对禾草/杂类草斑块中植物种重要值构成的影响比嵩草/杂类草斑块的大, 利于禾草/杂类草群落植物种的演变。
| 表1 禾草/杂类草和嵩草/杂类草斑块中主要植物种重要值 Table 1 The importance values (IV) of main species in grass/forb and Kobresia/forb patches (IV≥ 0.01) |
主要植物种种间Jaccard指数显示, 禾草/杂类草斑块中的DP和CK中, 联结性较强的种对为11.4%和14.0%; 嵩草/杂类草斑块的DP和CK中, 联结性较强的种对为38.9%和38.2%(表2)。说明, 两种斑块中, 较多种对联结性弱, 种间独立性较强, 且禾草/杂类草斑块比嵩草/杂类草斑块的植物种对的联结更为松散。
| 表2 禾草/杂类草斑块和嵩草/杂类草斑块主要植物种种间Jaccard指数 Table 2 The interspecific Jaccard indices of main plant species in grass/forb and Kobresia/forb patches |
Spearman秩相关分析显示, 禾草/杂类草斑块的正相关和负相关的种对数, DP为34和56对, CK为30和48对, DP和CK的正负关联种对比为0.61和0.63; 显著相关和不相关种对数, DP为6和84对, CK为5和73对, 显著联结种对为6.6%和6.4%(图1)。嵩草/杂类草斑块正相关和负相关的种对数, DP为17和19对, CK为23和32对, DP和CK的正负关联种对比为0.90和0.72; 显著相关和不相关种对数, DP为1和35对, CK为8和47对, 显著联结种对为2.8%和14.5%(图2)。说明, 两种斑块中多数种对间联结性弱, 群落处于不稳定阶段, 粪沉积使嵩草/杂类草斑块的稳定性增强。
 | 图1 禾草/杂类草斑块主要植物种Spearman秩相关分析半矩阵图Fig.1 Semi-matrix of Spearman rank correlation analysis of main species in grass/forb patches* P< 0.05, * * P< 0.01, the same below. |
在禾草/杂类草斑块中, DP为扁穗冰草-天蓝韭和西北针茅-藨草种对间显著正联结, 扁穗冰草-阴山扁蓿豆、扁穗冰草-垂穗披碱草、扁穗冰草-藨草和西北针茅-紫花针茅种对间显著负联结; CK为洽草-天蓝韭、洽草-大籽蒿、扁穗冰草-二裂委陵菜种对间极显著正联结, 西北针茅-紫花针茅、西北针茅-藨草、洽草-矮嵩草、洽草-大籽蒿、扁穗冰草-赖草、矮嵩草-天蓝韭和天蓝韭-赖草种对间显著负联结(图1)。在嵩草/杂类草斑块中, DP仅苔草-洽草种对间显著负联结, CK为矮嵩草-垂穗披碱草和瑞香狼毒-乳白香青种对间显著正联结, 珠芽蓼-洽草和瑞香狼毒-垂穗披碱草、线叶嵩草-矮嵩草、线叶嵩草-珠芽蓼、线叶嵩草-垂穗披碱草和垂穗披碱草-乳白香青种对间显著负联结(图2)。说明, 显著联结的种对多存在于优势种或亚优势种与伴生种之间。
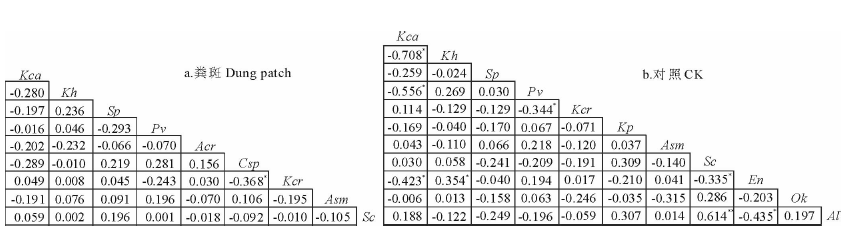 | 图2 嵩草/杂类草斑块主要植物种Spearman秩相关分析半矩阵图Fig.2 Semi-matrix of Spearman rank correlation analysis of main species in Kobresia/forb patches |
高寒草甸植物群落植物种的生态习性不同, 依据群落主要植物种对主导环境因子, 如光照和水分的适应方式, 以及植物种间关联性特征, 将两种斑块群落植物种划分为三大生态种组(表3)。其中, 第一生态种组为多数禾本科、豆科和菊科等喜阳耐旱植物, 主要包括扁穗冰草、西北针茅、紫花针茅、赖草、高山嵩草、甘肃棘豆、阴山扁蓿豆和大籽蒿等13种处于草层的中上层, 具有喜光、耐旱、耐土壤贫瘠等趋同特征的植物, 草甸退化标志植物瑞香狼毒亦出现在该组, 组内多物种间呈显著相关, 群落处于不稳定状态。第二生态种组禾本科的早熟禾、莎草科的藨草和苔草及蓼科的珠芽蓼4种喜阴喜湿植物, 因本研究区高寒草甸水分不足, 第二生态种组植物种少且组内各物种间无显著关联, 植物种间独立性较强。第三生态种组为禾本科的垂穗披碱草和洽草, 莎草科的矮嵩草和线叶嵩草, 菊科的球花蒿和百合科的天蓝韭等喜阳喜湿的植物, 高寒草甸很难满足其生境要求而仅由6种植物种构成, 其中线叶嵩草与垂穗披碱草和矮嵩草呈极显著负关联而垂穗披碱草和矮嵩草呈显著正关联, 且线叶嵩草是嵩草/杂类草斑块的优势种, 种组内竞争激烈, 种组有演替的趋势。
| 表3 禾草/杂类草和嵩草/杂类草斑块群落中主要植物种生态分组 Table 3 Ecological species groups of main species in grass/forb and Kobresia/forb patches |
本研究主要植物种Jaccard指数和种间Spearman秩相关分析均显示, 无论粪沉积与否, 禾草/杂类草斑块和嵩草/杂类草斑块中, 多数种对间联结性弱, 群落成熟度较低, 结构简单, 整体上处于不稳定状态。这是由于草地在长期演替过程中, 植物种的生态位进一步分化, 生境异质性增加[8], 各物种都占据有利的位置, 和谐共处, 相互依赖和相互竞争均大大降低, 多数种对联结程度不强, 独立性强[9], 从而使两种斑块的群落结构不稳定, 具有明显的次生性或正处于演替前期阶段。
本研究嵩草/杂类草斑块和禾草/杂类草斑块的植物种对数的正负关联比均< 1, 但前者更接近于1, 从而前者的群落稳定性比后者强。其主要原因是嵩草/杂类草斑块中的优势植物为嵩草属的线叶嵩草和矮嵩草, 其在长期适应寒冷气候的进化过程中, 形成特殊稳定的适应外界不良环境的维持性特征, 如叶片少, 植株矮化、细绒化和草毡表层加厚等极度发育等, 从而具有较强的抗外界干扰和自我调控能力[20]; 而禾草/杂类草斑块中的优势植物为扁穗冰草、西北针茅和洽草等, 因其叶片宽而多, 茎秆直立高大, 根系较深, 具较多须根, 对外界干扰敏感。这与房飞等[21]对新疆天山南坡高寒草原夏季牧场羊茅(Festuca ovina)+紫花针茅群落, 以及王伟等[10]和郭倩等[22]报道的青海省称多县藏嵩草(Kobresia tibetica)群落和高山嵩草+异针茅(Stipa aliena)/杂类草群落及河南县线叶嵩草草地群落, 植物种对的正负关联比均≥ 1, 且群落整体上处于稳定发展阶段的研究结果存在分异。这可能与二者取样大小不同有关, 本研究为测定粪斑效应, 将样方大小设为0.1 m2, 而这些相关研究的样方大小为0.25 m2, 一般样方大则正关联的种对数多, 反之亦然; 也可能是二者样方数量、群落自身物种数的多寡和群落类型等差异造成[11, 21, 22]。
本研究粪沉积使嵩草/杂类草斑块群落稳定性增强, 而对禾草/杂类草斑块群落稳定性影响小。类似施肥研究表明, 施肥降低群落稳定性[23], 施N肥及N、P、K单施或联施或施羊粪, 均增加青藏高原矮嵩草群落和退化高寒草甸草地中禾草比例, 降低杂类草比例[24, 25]。本研究与后者结果存在分异, 其主要原因是二者所施肥料类型不同。本研究牛粪不但富含氮、磷、钾及多种有机养分, 且为垫状形态, 其沉积后首先在草地上形成致密粪垫层, 进而对粪垫下的草地植物造成窒息而使其枯黄死亡, 因群落结构和植物建群种构成不同, 嵩草/杂类草斑块比禾草/杂类草斑块中处于上层的高大禾草和杂类草比例少, 从而前者比后者因粪窒息造成的枯黄死亡比例少。同时, 由于禾草相对莎草对牛粪和施肥反应更敏感, 更易吸收肥料中的养分[14], 粪沉积后期, 禾草/杂类草斑块中的禾草因在短时间易吸收利用牦牛粪中的养分, 生长增强, 迅速占领群落上层空间, 进而遮蔽其他矮小物种, 以此弥补了粪沉积前期因粪斑覆盖草地而造成的损失, 从而使其在与其他功能群植物的竞争中取胜, 而嵩草/杂类草斑块中的莎草吸收牛粪中的养分少且慢。因此, 粪沉积后, 嵩草/杂类草斑块向趋于嵩草群落的方向发展, 更趋于稳定; 而禾草/杂类草群落向趋于禾草的方向发展, 更趋于不稳定。同时, 由于粪斑具有特殊气味及寄生虫, 粪沉积初期-中期, 家畜弃食粪斑及其毗邻区域植物[26]; 随粪块沉积时间的延长及粪中养分的释放, 粪斑及其周围植物通过土-草系统吸收粪中养分后形成高营养斑块, 反过来又吸引家畜采食[26, 27], 从而家畜粪沉积和采食互作对草地植被的复杂效应, 使粪块及其周围区域植物群落构成和种间关系发生改变[6]。另一方面, 家畜具有在食源值与食源报酬之间的空间判断和权衡能力, 其也频繁采食草地上数量多、面积大的中、低营养植被斑块[28]。可见, 草地群落稳定性和发展演替受放牧干扰复杂效应的调控。本研究仅阐释了家畜粪沉积下草地植物群落构成及种间关系变化, 而牛粪沉积对草地植被的作用过程和机制, 还需进一步研究。
植物种间关联或物种联结性是指不同物种在空间上的相互联结性, 体现物种间的依赖性与排斥性, 正联结种对对环境或资源有相互依存的相似需求, 负联结种对生境或资源具有相互排斥的不同需求[29]。植物群落中优势种和亚优势种对群落的结构和稳定性起重要作用[30]。本研究禾草/杂类草斑块中, 粪沉积使西北针茅-藨草的种间关联由负显著变为正显著, 西北针茅-紫花针茅由显著负关联变为极显著负关联。这是由于西北针茅属密丛型浅根禾草, 具抗旱耐寒特征, 藨草属莎草科挺拔直立型根状茎植物, 具抗寒耐湿特征, 二者因生态特征差异对生境和资源具不同要求而互相排斥, 但随着牦牛粪的沉积, 两者吸收粪中养分资源而具有相似的资源需求所导致; 西北针茅和紫花针茅均为密丛型浅根同属禾草, 在长期进化和不同微环境适应过程中, 对生境具有不同的生态适应性或相互竞争而具显著负关联, 是其对不同空间资源利用的结果, 而粪沉积下二者对粪中养分资源利用程度的竞争加剧而转为极显著负关联。
本研究分析了高寒草甸植物种间关联及群落构成特征, 推测了群落的稳定性和演替动态, 其结果有助于认识草地群落的基本特征和物种替代机制, 为放牧家畜排泄物的深入研究奠定了基础。
无论粪沉积与否, 禾草或嵩草/杂类草斑块总体上群落植物种间关系均表现为负联结, 多数种对间联结性弱, 禾草/杂类草斑块比嵩草/杂类草斑块的植物种对的联结更为松散, 群落处于不稳定阶段。粪沉积对禾草/杂类草斑块中建群种重要值的影响比嵩草/杂类草斑块的大, 利于禾草/杂类草斑块的演变, 使嵩草/杂类草斑块的稳定性增强。依据两种斑块群落优势种对环境的适应方式和主导生态因子, 将植物种划为喜阳耐旱植物、喜阴喜湿植物和喜阳喜湿生植物三大生态种组。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|