



{kind=link}
{kind=link}
{kind=link}
{kind=link}
种群密度对沙米异速生长的影响
[范高华1, 2  , 黄迎新
, 黄迎新1* , 赵学勇3 , 神祥金1 ]
, 黄迎新, 赵学勇|
|


作者简介:范高华(1991-),男,甘肃兰州人,在读硕士。E-mail: fangaohua@iga.ac.cn
植物各器官之间的异速生长关系决定了植物的生活史对策,进而影响种群内个体之间的共存和生物多样性的维持。传统方法无法区分植物性状对其的响应是环境造成的,还是由于个体大小差异造成的,而异速生长分析能够解决这一问题。为此,本研究利用异速生长分析方法,通过模拟沙米在退化草地恢复过程中不同的种群密度(16, 44.4, 100和400株/m2),研究其各器官之间的异速生长关系。结果表明,沙米各器官之间都具有显著的异速生长关系:其中根和地上、根和主茎、叶和根、主茎和地上等器官生物量之间的变化不随种群密度的变化而变化,属于表观可塑性;根和茎、茎和地上、叶和茎等器官生物量之间的异速生长及生物量比值均受种群密度的显著影响,属于真正可塑性,这表明种群密度变化的确影响了沙米的生长变化。研究同时发现,沙米叶和地上生物量之间的异速指数仅在44.4株/m2种群密度下接近于0.75,即叶生物量与个体大小呈3/4次幂关系,符合生态代谢理论,而在其他密度时异速指数与0.75有显著差异( P<0.001),表明环境变化可能会导致沙米生长策略发生改变。种群密度对沙米繁殖和其他器官之间的异速生长关系影响不显著,表明在不同种群密度下,沙米仍保持相同的繁殖策略;然而种群密度在沙米生育期对繁殖和其他器官之间的异速常数和个体大小均产生了极显著的影响。因此,在探讨繁殖器官与个体大小的关系时,还需要关注植物生育期的变化,只有生育期相同的时候,才能比较不同研究之间的差异。
The allometric relationships between plant organs determine the strategies of life history, affecting the coexistence and biodiversity of species. Traditional methods are not able to distinguish the responses of plant traits to environmental or genetic differences but allometric analysis can help solve this problem. Allometric analysis was used to study the relationships between different organs of Agriophyllum squarrosum by simulating different population densities of A. squarrosum (16, 44.4, 100 and 400 plants/m2) during the restoration of degraded grassland. The results showed that there are significant allometric relationships between organ pairs. Changes in root vs. aboveground biomass, root vs. main stem biomass, leaf vs. root biomass and main stem vs. aboveground biomass were not influenced by population density, indicating ‘apparent plasticity’. The allometric relationships of root vs. stem, stem vs. aboveground, leaf vs. stem biomass and their respective biomass ratio were significantly affected by population density, reflecting ‘real plasticity’. The allometric index of leaf vs. aboveground biomass at 44.4 plants/m2 was approximately 0.75, while the allometric indices at other plant densities were significantly different from 0.75 ( P<0.001), suggesting that environmental changes resulted in changes in growth strategy of A. squarrosum. The influence of population density on the allometric relationships between reproductive and other organs is not significant, indicating that A. squarrosum will adopt the same strategy under different densities, but that density differences will result in different allometric scaling and size of individual reproductive organ during different periods of reproductive development. It is important to consider changes in plant growth stages when investigating the relationships between reproductive organs and individual plants.
不同的植物器官代表了植物的不同功能, 植物各器官所占生物量分配比例的变化指示了植物资源获取以及能量分配的调整, 植物各器官之间的异速生长关系一直是植物生态学研究的重要内容。当植物生长的环境发生变化时, 其生理性状以及生态性状会产生相应的适应性变化。特别是关于自疏法则以及最终产量恒值法则等方面的研究, 促进了生态学家对植物密度变化的生态学意义的广泛关注。近年来, 生态学家对植物响应种群密度和时间变化的异速生长进行了大量研究[1, 2, 3]。已有的研究结果表明, 传统分析方法无法揭示哪些变化是由种群密度和时间变化引起的, 哪些变化是由个体大小变化引起的[4, 5], 而异速生长分析能够排除个体大小变化的因素, 分析植物不同性状之间的关系, 揭示植物性状响应种群密度的变化规律, 从而受到了广泛关注[2, 6]。
异速生长(allometry)是指植物不同器官或不同性状之间不同比例的生长关系[7]。通常是描述植物生长过程中个体大小与植物体结构、功能之间的定量关系, 是一个描述植物体形态学上进化的重要方法, 通常以幂函数的形式表示[8, 9, 10, 11]: Y=β Xa, 式中:X是与Y不同的另一个属性, Y是某个需要研究的属性值, X是植物的个体大小, 它也可以表示植物体的两个不同器官大小或不同属性之间的相关关系; α 是异速指数; β 是异速常数。
植物体的异速生长是由物种遗传性所决定的一种固有特性, 在植物的生长发育过程中, 植物的各部分器官呈一种明显的异速生长规律[12, 13]。因此, 异速生长分析能够系统阐明种群密度、个体大小以及植物性状之间的内在关系[5, 14]。然而, 最近有研究表明, 植物的异速生长显著受生育期的影响, 通过一次取样进行异速生长分析的结果是不严谨的, 应该进行多次取样, 对植物整个生育期进行系统研究, 才能揭示植物不同器官的异速生长关系[5]。
沙米(Agriophyllum squarrosum)又称沙蓬, 为藜科一年生草本, 是我国北方地区的一种藜科沙生植物, 株高14~60 cm[15, 16]; 沙米种群主要生长在流动沙丘上, 在半固定沙丘和固定沙丘中仅生长在裸沙斑上, 是流动沙丘恢复过程中的主要先锋物种, 也是人工固沙的一种常见植被[17, 18]。它能增加下垫面粗糙度, 能有效地防风阻沙; 在退化沙地或沙丘的演替过程中, 其密度变化极为剧烈。以往关于沙米的研究主要集中在种子萌发以及生理生态学特征等方面[19], 而有关种群密度变化对沙米生长特性的影响研究较少, 深入系统地研究沙米生长特性与种群密度的关系有助于了解退化土地恢复机理, 对沙漠化的防治具有重要意义。因此, 本研究探究了种群密度变化对沙米营养生长和繁殖生长的影响, 以及对营养生长与繁殖生长之间异速生长关系的影响, 并确定沙米在不同种群密度下的适应策略, 一方面探讨植物性状变化的生态学机理, 另一方面以期能为退化土地的恢复和荒漠化防治提供一定的理论基础。
试验地点位于内蒙古自治区奈曼旗境内中国科学院奈曼沙漠化研究站(42° 54' N, 120° 41' E)。该地区属半干旱气候, 年均降水量366 mm, 年蒸发量1935 mm, 年均气温6.5 ℃, ≥ 10 ℃年积温3000 ℃以上, 无霜期150 d。土壤类型为沙质栗钙土, 沙土基质分布广泛, 风沙活动强烈, 年均风速3.5 m/s, 8级以上大风年均21次, 主要发生在3-5月[18]。
2007年5月初, 根据沙地上沙米自然种群随着退化沙地恢复过程中的密度变化范围, 在沙质草地通过机械剔除杂草, 对沙米进行种群密度处理, 共设置4个处理水平, 每个处理4个重复, 共16个小区, 小区面积(3 m× 3 m):1)16株/m2(D1); 2)44.4株/m2(D2); 3)100株/m2(D3); 4)400株/m2(D4)。在2叶期(大约种植2周左右)进行间苗, 按照D1~D4密度处理定苗。整个生长阶段, 每3~4周进行一次人工除草。
于2007年6月22日、7月14日、8月6日、8月23日和9月9日分别进行破坏性取样, 每次每个小区随机取样10株, 每种密度处理共计40株。用清水将每株沙米的根冲洗干净, 以保证获取较为完整的根系, 并将每株植物分成根、茎(主茎和侧枝分开)、叶和繁殖器官(包括花和果)4部分, 将植物放置于80 ℃烘箱内烘干称重, 然后测量得到根、茎、叶、主茎、地上和繁殖器官等的生物量(繁殖只取9月份数据进行分析), 计算叶/地上、叶/根、叶/茎、叶/主茎、根冠比、根/茎、根/主茎、茎/地上、主茎/地上、繁殖/地上、繁殖/根、繁殖/茎、繁殖/叶及繁殖/主茎等器官生物量比以及根、茎、叶、主茎和繁殖等器官的生物量分配, 器官生物量分配=器官生物量/总生物量, 即各器官相对生物量。
采用SPSS 17.0软件进行数据分析, 采用单因素[种群密度(D)]方差分析比较各器官生物量特性以及生物量分配的可塑性。处理间差异显著性的检验采用F-检验, 处理间平均数的比较采用最小显著性差异检验(LSD检验)。采用经典的异速方程进行异速生长分析, Y=β Xα , 式中:Y是某个需要研究的器官的属性值; X是植物的个体大小或另一个器官的属性值; α 是异速指数, 即斜率; β 是异速常数, 即Y轴截距。采用减小主轴回归(reduced major axis regression, RMA)拟合经对数转换后的数据, 确定异速指数:α RMA=α OLS/r(α OLS是最小二乘法得到的回归系数; r是相关系数), 以去除变量偏差对回归系数的影响[20]。用R软件的SMATR包, 将数据log10转换后, 进行标准主轴回归, 分析密度对异速生长关系的影响, 主要包括异速指数(斜率)、异速常数(截距)以及个体大小(X值变化), 并比较异速指数与0.75或1的差异[21]。
通过比较沙米叶与其他器官生物量之间的异速关系, 发现叶与根生物量之间的异速生长不随种群密度的变化而变化, 而叶和其他器官生物量之间具有显著的异速生长关系:叶∶ 地上、叶∶ 茎、叶∶ 主茎生物量的异速生长显著受种群密度的影响(P< 0.05)(图1, 表1)。密度对沙米叶∶ 地上、叶∶ 根、叶∶ 茎、叶∶ 主茎生物量的异速常数和个体大小均产生了极显著的影响(P< 0.001); 叶∶ 地上、叶∶ 根、叶∶ 茎、叶∶ 主茎生物量的异速指数均小于1(图1, 表1)。随着种群密度的增大, 沙米叶/地上、叶/茎生物量呈增大趋势, 叶/根、叶/主茎生物量呈减小趋势(表1)。沙米叶/地上、叶/根生物量在不同种群密度之间差异极显著(P< 0.001), D1、D2、D3与D4密度间差异显著, D1、D2、D3密度之间差异不显著; 叶/茎、叶/主茎生物量在不同种群密度之间差异极显著(P< 0.01), 叶/茎生物量D1与D2、D3、D4密度间差异显著, D2、D3、D4密度之间差异不显著, 叶/主茎生物量D1、D2与D4密度间差异显著, 其他密度之间差异不显著(表1)。沙米叶生物量特性明显受种群密度变化的影响, 叶生物量随种群密度的增大而显著减小, 叶生物量与密度之间呈极显著负相关(F=96.321, P< 0.001), D1~D4密度之间差异均显著(表2)。密度对沙米叶生物量分配的影响不显著(F=1.065, P=0.363), D1~D4密度之间差异均不显著(表3)。
| 表1 种群密度对沙米各器官间生物量比特性的影响 (平均值± 标准误) Table 1 The influence of A. squarrosum under population density on the biomass ratio characteristics of the different organs (means± SE) |
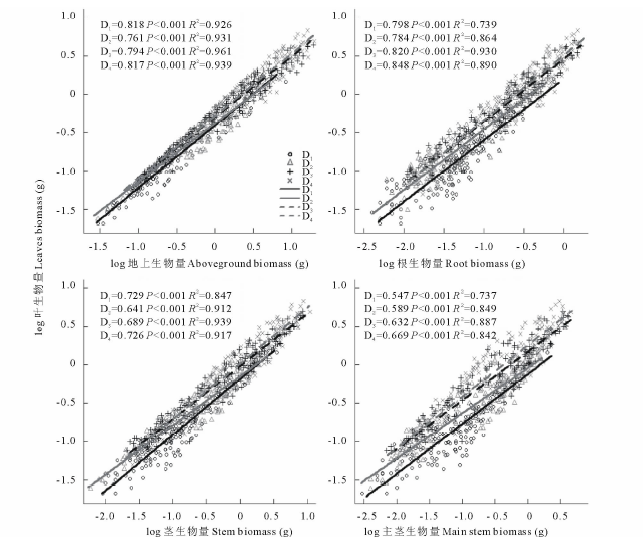 | 图1 叶生物量与其他器官生物量之间的异速关系D1, D2, D3, D4表示16, 44.4, 100和400株/m2处理, 下同。D1, D2, D3, D4 symbol for 16, 44.4, 100 and 400 plants/m2 treatments. The same below.Fig.1 The allometric relationship between leaf biomass and other organ biomass |
随着种群密度的变化, 沙米根∶ 茎生物量间具有显著的异速生长关系(P=0.006), 根∶ 地上、根∶ 主茎生物量的异速生长关系均不显著(P> 0.05) (图 2, 表1)。种群密度对沙米根∶ 茎生物量的异速常数的影响不显著(P=0.075), 对个体大小的影响极显著(P< 0.001); 对沙米根∶ 地上、根∶ 主茎生物量的异速常数和个体大小均产生了极显著的影响(P< 0.001); 沙米根∶ 地上生物量在D1密度时的异速指数大于1, 而在其他密度时的异速指数均小于1, 根∶ 茎、根∶ 主茎生物量的异速指数均小于1(图 2, 表1)。随着种群密度的增大, 沙米根冠比、根/茎、根/主茎生物量均呈增大趋势(表1)。根冠比、根/茎生物量在不同种群密度之间差异极显著(P< 0.001), 根/主茎生物量在不同种群密度之间差异不显著(P=2.003), 根冠比、根/茎生物量D1与D2密度之间差异不显著, 其他密度间差异显著, 根/主茎生物量D1与D4、D3与D4密度间差异显著, 其他密度间差异不显著(表1)。沙米根生物量特性明显受种群密度变化的影响, 根生物量随种群密度的增大而显著减小, 根生物量与密度间呈极显著负相关(F=74.074, P< 0.001), D3与D4密度间差异不显著, 其他密度间差异显著(表2)。沙米根生物量分配随种群密度的增大而增大, 密度对根生物量分配的影响极显著(F=66.693, P< 0.001), D1与D2密度间差异不显著, 其他密度间差异显著(表3)。
 | 图2 根生物量与其他器官生物量之间的异速关系Fig.2 The allometric relationship between root biomass and other organ biomass |
| 表2 种群密度对沙米生物量特性的影响 (平均值± 标准误) Table 2 The influence of population density on A. squarrosum biomass characteristics (means± SE)g |
系及其分配特性种群密度对沙米茎∶ 地上生物量的异速生长产生了极显著的影响(P< 0.001), 对主茎∶ 地上生物量的异速生长影响不显著(P=0.171); 对茎∶ 地上、主茎∶ 地上生物量的异速常数和个体大小均产生了极显著的影响(P< 0.001); 茎∶ 地上、主茎∶ 地上生物量的异速指数均大于1(图3, 表1)。沙米茎/地上生物量随种群密度的增大呈减小趋势, 而主茎/地上生物量则相反(表1)。 茎/地上、主茎/地上生物量在不同种群密度之间差异极显著(P< 0.001), 茎/地上生物量D1与D3、D4密度间差异显著, 其他密度之间差异不显著, 主茎/地上生物量D1与D2密度间差异不显著, 其他密度之间差异显著(表1)。沙米茎、主茎以及地上生物量特性明显受种群密度变化的影响, 茎、主茎以及地上生物量随着种群密度的增大而减小, 茎、主茎以及地上生物量与密度之间呈极显著负相关(F=64.626, F=56.421, F=66.704, P< 0.001), 沙米茎、地上生物量D3与D4密度间差异不显著, 其他密度间差异显著, 沙米主茎生物量D1~D4密度间差异均显著(表2)。随着种群密度的增大, 沙米茎、地上生物量分配呈减小趋势, 主茎生物量分配呈增大趋势, 种群密度对沙米茎、主茎和地上生物量分配均产生了极显著的影响(F=7.008, F=9.187, F=66.693, P< 0.001), 茎生物量分配D1与D3、D1与D4、D2与D3密度间差异显著, 其他密度之间差异不显著, 地上生物量分配D1与D2密度间差异不显著, 其他密度之间差异显著, 主茎生物量分配D1与D2、D3与D4密度间差异不显著, 其他密度之间差异显著(表3)。
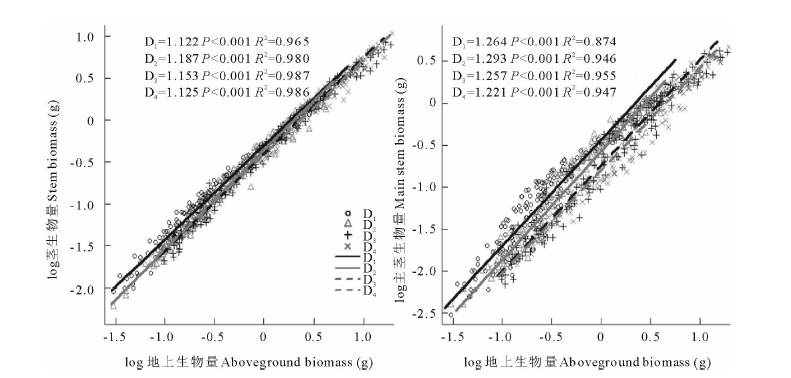 | 图3 茎/主茎生物量与地上生物量之间的异速关系Fig.3 The allometric relationship between stem/main stem biomass and aboveground biomass |
| 表3 种群密度对沙米生物量分配特性的影响 (平均值± 标准误) Table 3 The influence of population density on A. squarrosum biomass distribution characteristics (means± SE)g |
由于沙米集中在9月份进行繁殖, 所以本研究选择9月份的数据作为繁殖与其他器官生物量间异速生长分析的数据。结果表明, 种群密度对沙米繁殖∶ 地上、繁殖∶ 根、繁殖∶ 茎、繁殖∶ 叶、繁殖∶ 主茎生物量的异速生长关系均无显著影响(P> 0.885), 对繁殖∶ 地上、繁殖∶ 根、繁殖∶ 茎、繁殖∶ 叶、繁殖∶ 主茎生物量的异速常数和个体大小均产生了极显著的影响(P< 0.004) (图 4, 表1)。繁殖∶ 地上、繁殖∶ 根、繁殖∶ 茎、繁殖∶ 叶、繁殖∶ 主茎生物量的异速指数均大于1(图 4)。总体上看, 随着种群密度的增大, 繁殖/根、繁殖/主茎生物量呈减小趋势, 而繁殖/地上、繁殖/茎、繁殖/叶生物量不随种群密度的变化而变化(表1)。繁殖/地上、繁殖/根、繁殖/茎、繁殖/叶、繁殖/主茎生物量在不同种群密度之间均无显著相关性(P> 0.445), 繁殖/地上、繁殖/茎、繁殖/叶生物量D1~D4密度间差异均不显著, 繁殖/根生物量D1、D2、D3与D4密度间差异显著, D1、D2、D3密度之间差异不显著, 繁殖/主茎生物量D1与D4密度间差异显著, 其他密度之间差异不显著(表1)。沙米繁殖生物量特性明显受沙米种群密度变化的影响, 随着种群密度的增大, 繁殖生物量呈减小趋势, 繁殖生物量与密度之间呈极显著负相关(F=10.337, P< 0.001), D1与D2、D3与D4密度间差异不显著, 其他密度之间差异显著(表2)。密度对沙米繁殖生物量分配无显著影响(F=0.785, P=0.505), D1~D4密度间差异均不显著(表3)。
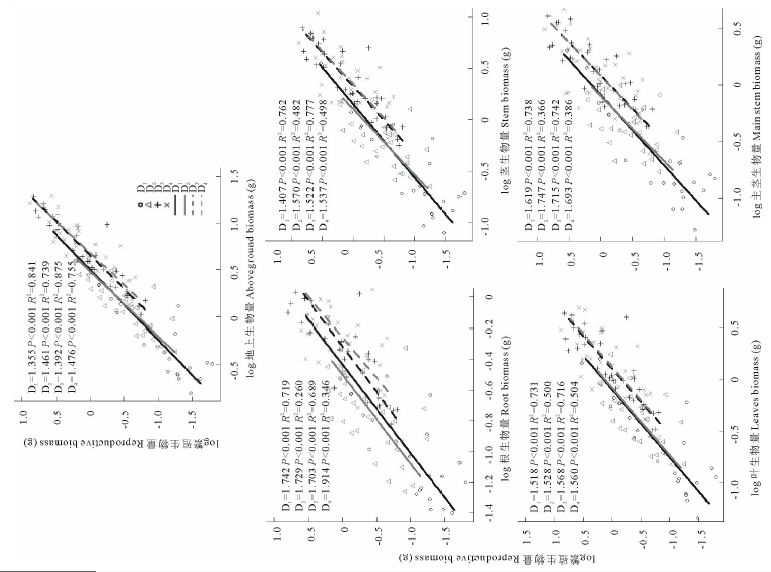 | 图4 繁殖生物量与其他器官生物量之间的异速关系Fig.4 The allometric relationship between reproductive biomass and other organ biomass |
种群密度调控作用的本质是植物种群内个体间因密度变化而产生对空间与资源的竞争以及个体之间的相互干扰, 而植物资源获取以及能量分配, 一直是植物生态学领域的研究热点[22], 本研究结果与前人的研究结果均表明各器官之间都具有显著的异速生长关系。在植物生活史的不同阶段, 光合产物和土壤营养等环境资源在植物地上和地下构件的分配格局以及各构件生物量之间特定的异速生长关系导致不同构件种群的异速指数大不相同[23]。异速指数的差异是异速生长过程中植物各部分构件之间存在特定的异速生长模式的反映, 也是植物体内有限资源的分配体现。虽然3/4法则是从生物能量学和异速生长的假设出发, 利用个体新陈代谢与个体大小的基本异速生长关系和环境资源对植物生长限制的基础上推导出植物个体大小和种群密度的数量关系[10], 但由种群密度导致的光合产物和土壤营养等环境资源在植物不同构件的分配比例的不同导致异速指数偏离3/4, 因此, 并不存在一个对植物不同构件生物量普适的生物量-种群密度的异速生长关系。
当外界环境发生改变时, 植物的个体大小和叶生物量分配也会产生相应的变化。本研究中, 叶∶ 根生物量间的异速生长受种群密度的影响不显著, 而叶/根生物量显著受种群密度影响, 这表明叶和根之间的变化是受个体大小的制约, 而不是真正响应种群密度而发生的变化, 这种可塑性是表观可塑性[5, 14]。叶与其他器官生物量间的异速生长及生物量比值均受种群密度的显著影响, 属于真正可塑性, 这表明种群密度变化的确影响了叶的生长变化。叶生物量与地上生物量的异速指数在D2密度时接近于0.75(P=0.470), 即叶的生物量与个体大小呈3/4次幂关系, 符合生态代谢理论, 但是, D1、D3和D4密度时异速指数与0.75有显著差异(P< 0.001), 表明环境变化会导致这个斜率发生改变, D2密度可能是沙米在荒漠化生境中生长的最佳种群密度。因此, 在未来的研究中, 需要考虑环境的异质性对不同器官之间生物量的影响。叶生物量与茎生物量的异速指数在D1和D4密度时接近于0.75(P=0.355, P=0.157), 符合生态代谢理论的结论, 但是, D2和D3密度时异速指数与0.75有显著差异(P< 0.001), 表明随着种群密度的变化, 异速指数在1/2~3/4范围内变化, 这与Deng等[24]对植物种群个体间几何关系和代谢理论的研究一致。叶生物量与主茎生物量的异速指数在D1~D4密度时与0.75有显著差异(P< 0.001), 异速指数在0.589~0.669范围内变化, 即种群密度的调控作用在D1~D4密度不断增加, 表明环境因素可以通过密度调控来影响植物之间的相互作用[11, 25, 26], 使其在不同生境压力下, 通过种群内的相互作用实现种群内的优胜劣汰。因此, 基于种群密度的变化, 沙米种群在生长过程中为应对环境资源供不应求的矛盾, 个体之间对有限资源的竞争会趋于激烈, 即种群个体之间微小的差异通过对有限资源的竞争呈现出来。这种可能基于基因或空间位置上的微小优势通过种群个体之间对有限资源的竞争而不断放大和积累, 使得具有优势的个体在资源竞争中胜出, 而处于劣势的个体被淘汰[11], 因此, 竞争可以在个体水平上改变植物的生长速度和性状, 进而影响植物个体之间的异速生长关系。
叶片是植物能量和物质转化的重要场所, 也是生态系统中物质循环和能量流动的主要驱动者[27]。叶生物量可以同时反映植物的光合能力和对土壤中水分及营养物质的吸收能力[24]。本研究中, 沙米叶生物量与种群密度成反比, 叶生物量随种群密度的增大而减小(表2), 这主要是由于密度的增加, 植物种内竞争加剧, 资源获取受到了限制, 所以叶的生长受到了抑制, 冠层底部的叶片因受到遮蔽而生长受阻[28, 29, 30, 31, 32]; 然而与叶生物量结果不同, 沙米叶生物量分配比例随密度增大而增大, 但是异速指数没有发生变化, 这表明在不同密度下, 虽然植物的叶生物量分配发生了变化, 但是其变化是由不同密度下, 个体大小差异造成的, 而不是响应环境变化而发生了真正的策略改变, 这种变化是表观可塑性, 而不是真正的可塑性[4, 14]。
本研究中, 根∶ 地上生物量间的异速生长受种群密度影响不显著, 而根/地上生物量显著受种群密度影响, 表明根和地上生物量间的变化是受个体大小制约的, 而不随种群密度发生变化, 属于表观可塑性[33, 34, 35]。亦有研究表明, 植物根与地上生物量间为等速生长关系, 即幂指数为1[33, 34, 36, 37]。根∶ 主茎生物量间的异速生长不随种群密度的变化而变化, 并且根/主茎生物量在不同种群密度间无显著差异, 属于表观可塑性。根∶ 茎生物量间的异速生长及生物量比例均受种群密度的显著影响, 表明种群密度的变化的确影响了根和茎的生长变化, 属于真正可塑性。主茎与根无异速生长关系, 但是随着密度的变化, 总茎生物量发生了显著变化, 这表明侧枝生物量的显著变化, 导致了植物总茎生物量的显著变化。
随着种群密度的变化, 沙米根生物量明显小于茎生物量(表2), 根生物量与茎生物量的异速指数在D1~D4密度时均小于1, 且与0.75有显著差异(P< 0.001), 表明随着沙米种群密度的增加, 茎生物量分配要大于根生物量分配, 这意味着沙米为维持生存, 必须充分利用地下的营养物质, 尤其是水分, 并通过茎等组织将营养物质输送到地上部分, 说明对于基因型相同的植物个体, 其根、茎等器官响应不同种群密度具有可塑性。因此, 在荒漠生境中沙米为了完成生活史, 根系相对发达, 占据较大空间, 并且茎秆粗硬, 具有较高的机械强度和抗风蚀能力, 这也为其他物种的入侵提供了庇佑, 促进了群落演替和植被发育进程, 从而改善退化土壤, 减轻荒漠化程度。
本研究中, 主茎∶ 地上生物量间的异速生长关系受种群密度的影响不显著, 而主茎/地上生物量显著受种群密度的影响, 表明主茎和地上生物量间的变化是受个体大小制约的, 而不是真正响应种群密度而发生的变化, 属于表观可塑性。茎∶ 地上生物量间的异速生长及生物量比例均受种群密度的显著影响, 随着种群密度的增大, 茎生物量与个体大小呈异速生长关系, 这表明种群密度变化显著影响了茎的生长变化, 这种可塑性属于真正可塑性[7]。茎生物量与地上生物量的异速指数在D1~D4密度时均大于1, 且与1有显著差异(P< 0.001), 不符合最终产量恒值法则; 而且茎生物量与地上生物量在D1~D4密度时异速指数与0.75有显著差异(P< 0.001), 也不符合生态代谢理论。谢然等[38]对一年生荒漠植物的研究也支持这一结论。
以往研究关于植物生物量分配格局是否受到种群密度制约调节, 以及植物在不同竞争强度下将采取何种资源分配对策尚无一致定论。彭方仁等[39]、方华等[40]认为, 虽然种群密度对植物各器官生物量有显著影响, 但各器官生物量分配比例在不同种群密度条件下却大致相同。本研究中, 沙米各器官生物量均受种群密度的显著影响, 并且茎及地上生物量分配均随种群密度的增大呈减小趋势, 然而, 沙米叶及繁殖生物量分配不随种群密度的变化而变化, 根及主茎生物量分配随种群密度的增大呈增大趋势, 与上述结论不一致, 这可能与本研究的植物有关, 随着种群密度的增加, 沙米会选择增大根及主茎生物量的分配, 进而利用有限的环境资源完成其生活史。
生活史和竞争可能对植物繁殖分配产生重要影响[41], 当种群密度变化时, 植物的个体大小和繁殖分配也会产生相应的变化[42, 43]。沙米繁殖生物量与种群密度间差异显著, 繁殖分配及其与其他器官之间的异速指数均不随种群密度的变化而变化, 表明在不同密度处理下, 沙米保持相同的繁殖策略, 说明沙米的繁殖分配比例受其遗传性的控制而相当稳定, 并且是非大小依赖的, 这与黄迎新等[7]对沙米的研究结果一致。本研究中, 繁殖生物量与个体大小的异速指数在D1~D4密度时与0.75有显著差异(P< 0.001), 而且繁殖生物量与个体大小的异速指数在D1~D4密度时显著大于1(P< 0.002), 异速指数在1.355~1.914范围内变化, 显著区别于前人的结论。这可能与本研究方法有关, 本研究从繁殖器官出现一直到成熟期, 植物的繁殖器官从无到有, 变化十分显著, 而其他器官生物量的变化不像繁殖器官变化这样明显, 而且, 在生长后期, 植物生物量接近了稳定期, 所以导致了繁殖与地上生物量间的异速指数显著大于1。也有研究表明, 随着种群密度的增加, 植物可能趋向于将更多地资源分配给繁殖器官[44], 并且繁殖分配不受种群密度的影响[45], 本研究的结论与其一致, 沙米各器官生物量及各器官分配随着种群密度的增加而减小, 但繁殖生物量始终高于其他器官生物量; 尽管繁殖分配所占比例较低, 但在各种群密度之间无显著差异, 这表明沙米的繁殖输出是依赖于个体大小的, 即沙米的繁殖努力会随其平均个体大小的减小而减小, 而沙米的平均个体大小会随着沙米种群密度的增大而减小。本研究结果表明, 植物生育期的变化, 对繁殖生物量与个体大小的关系具有显著影响, 在探讨繁殖器官与个体大小的关系时, 需要关注植物生育期的变化, 只有生育期相同的时候, 才能比较不同研究之间的差异。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|