
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
基于MaxEnt模型和不同气候变化情景的单叶蔓荆潜在地理分布预测
[王亚领1  , 李浩
, 李浩1 , 杨旋1 , 郭彦龙2 , 李维德1, * ]
, 李浩]
|
|


作者简介:王亚领(1991-),男,湖北黄冈人,在读硕士。E-mail:wangyaling14@lzu.edu.cn
单叶蔓荆为我国常用中药蔓荆子的来源之一,不仅具有良好的药用价值,还具有很高的生态效益,能很好地防风固沙和保持水土。预测气候变化对该物种分布范围的影响可以为单叶蔓荆的可持续利用提供科学基础和参考依据。本研究利用获得的单叶蔓荆126个地理分布记录和22个环境因子,利用MaxEnt模型分析了单叶蔓荆在我国全国范围内的潜在地理分布,并基于该模型预测了4种气候情景下21世纪50和70年代单叶蔓荆分布范围。结果表明,最大熵模型预测单叶蔓荆潜在生境分布的精度较高(接收者操作特征曲线下方的平均面积为0.988),海拔、平均气温日较差、最冷季度降水量和最干月份降水量是影响单叶蔓荆分布的主要气候因子。由模型预测可知,在4种气候情景下,单叶蔓荆在全国适宜生境和低适宜生境的数量均有不同幅度的增加,适宜生境增加较快,低适宜生境增加缓慢;到2050s阶段,适宜生境比例由当前的5.03%分别上升到15.88%、17.00%、17.59%和23.11%;低适宜生境比例由当前的8.86%分别上升到11.09%、10.31%、11.53%和12.96%;到2070s阶段,适宜生境比例分别上升到21.22%、22.21%、24.57%和30.66%;低适宜生境比例分别上升到11.85%、12.07%、13.99%和14.66%。空间分布上,单叶蔓荆的适宜生境和低适宜生境的范围及几何中心都由沿海地区向内陆扩散;湖南和江西两省的适宜生境比例增长较快,尤其在四川境内,当前只有很小比例的低适宜生境,随着气候的变化,低适宜生境面积有所上升,并且适宜生境开始出现且增长速度较快。
Vitex trifolia var. simplicifolia ( Viticis fructus) is not only valuable as a medicinal plant, but also ecologically important. It can function as a wind break, and it stabilizes sand and soil, and it conserves water. Predicting the impact of climate change on the spatial distribution of V. trifolia var. simplicifolia may provide a scientific basis and reference for the sustainable use of this important plant. Using a Maximum Entropy (MaxEnt) model, we simulated the geographical distribution of V. trifolia under the current climatic conditions in China based on species presence data at 126 locations and data for 22 environmental factors. Then, we used the model to predict the future distributions of V. trifolia in two periods (2050s and 2070s) under four different climate change scenarios. The results showed that the MaxEnt model was highly accurate (mean area under ROC curve, 0.988). The main climatic factors influencing the geographic distribution of V. trifolia were altitude, mean diurnal air temperature range, precipitation in the coldest quarter, and precipitation in the driest month. The model simulations indicated that, under the four scenarios, V. trifolia will widen its distribution because of a rapid increase in suitable habitat areas and a slow increase in marginally suitable habitat areas. During the period of 2041-2060, the potential distribution area of suitable habitat would increase from the current ratio of 5.03% to 15.88%, 17.00%, 17.59%, and 23.11% under scenarios 1-4, respectively. The potential distribution area of marginally suitable habitat would increase from the current ratio of 8.86% to 11.09%, 10.31%, 11.53%, and 12.96% under scenarios 1-4, respectively. During the period of 2061-2080, the potential distribution area of suitable habitat would increase to 21.22%, 22.21%, 24.57%, and 30.66% under scenarios 1-4, respectively, and the potential distribution area of marginally suitable habitat would increase to 11.85%, 12.07%, 13.99%, and 14.66% under scenarios 1-4, respectively. In terms of the spatial distribution of the potential habitat area of V. trifolia, both the distributional range and the center of distribution of suitable and marginally suitable habitat areas would shift from coastal areas to inland. The distribution of suitable habitat area in Hunan and Jiangxi provinces would increase rapidly. Especially in Sichuan Province, where there is only a small percentage of marginally suitable habitat area currently, the potential marginally suitable habitat area would increase, and the suitable habitat area would appear and increase rapidly under climate change.
环境因素是决定物种分布的最主要因素, 并且物种分布格局的变化对环境因素的变化最为敏感。气候作为环境因素中最主要的因素之一, 是我们重要的关注对象和研究方向。目前, 准确确定气候变化对濒危物种分布的影响是有效保护物种的理论基础, 也是当前保护生物学家面临的重要问题[1]。
物种分布模型(species distribution modeling, SDM)作为一种评估入侵物种潜在威胁的模型, 越来越广泛的应用于个体生态学研究[2], 此外, 该类模型在预估环境因素变化对物种分布影响方面也日益发挥着重要的作用, 在预测种群分布动态及区域物种多样性变化领域已得到广泛应用[3, 4]。最大熵模型 (maximum entropy model, MaxEnt)是一种基于有限的已知信息推断未知概率分布的数学方法, 其理论起源于信息科学, 是统计学研究的重要内容, 认为实现方式数量最大的可能性, 就是实际观察到的真实状况[5, 6]。MaxEnt模型作为一种预测物种未来地理潜在分布的定量化分析工具, 已经在很多领域得到了应用, 包括药材、珍稀树木、哺乳动物等多种物种的分布预测[7, 8, 9]。MaxEnt对于物种分布数据不全的情况也有很好的分析结果, 说明该模型具有很强的适用性[10]。
单叶蔓荆(Vitex trifolia var. simplicifolia)属于马鞭草科, 是一种灌木。开花后结的果实称蔓荆子, 蔓荆子是一种药用价值很高的中药材, 不但可以清湿热, 还具有止血的作用。单叶蔓荆主要分布在我国山东、江西、湖南、安徽、浙江、海南等省, 尤其在江西和山东两省分布最广泛[11]。单叶蔓荆是一种药用价值和生态价值都很高的药材, 已被我国列入《国家重点保护野生药材物种名录》, 但近年来由于人为活动的影响, 再加上自然环境的剧烈变化, 其野生数量和生长环境都遭到了严重的影响[11]。
到目前为止, 国内外对单叶蔓荆的研究主要集中在生态特性、生理特性、药理活性、群落特点以及化学成分[12, 13, 14]。目前国内外在单叶蔓荆地理分布性预测方面尚属空白, 导致单叶蔓荆的分布范围没有得到详细的记录。众所周知, 环境因素的变化, 尤其是气候的变化必然会严重改变单叶蔓荆的生长环境, 进而影响其地理分布, 所以, 预测不同气候条件下该物种的潜在地理分布, 对其保护和可持续利用以及评估都具有重要的意义。为此, 本研究首先根据MaxEnt模型绘制了当前气候情形下我国的单叶蔓荆潜在分布图, 在政府间气候变化专门委员会(IPCC)发布的RCP2.6(最低温室气体排放情景)、RCP4.5(中等温室气体排放情景)、RCP6.0(较高温室气体排放情景)、RCP8.5(最高温室气体排放情景)4种气候情景下, 基于该模型对21世纪50和70年代单叶蔓荆的分布范围进行了预测。此外, 还阐述了单叶蔓荆的空间分布与环境因子的关系, 为单叶蔓荆原生生境的保护和资源利用评估等研究提供理论依据。
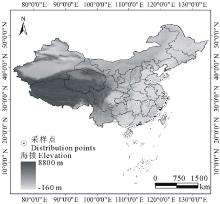
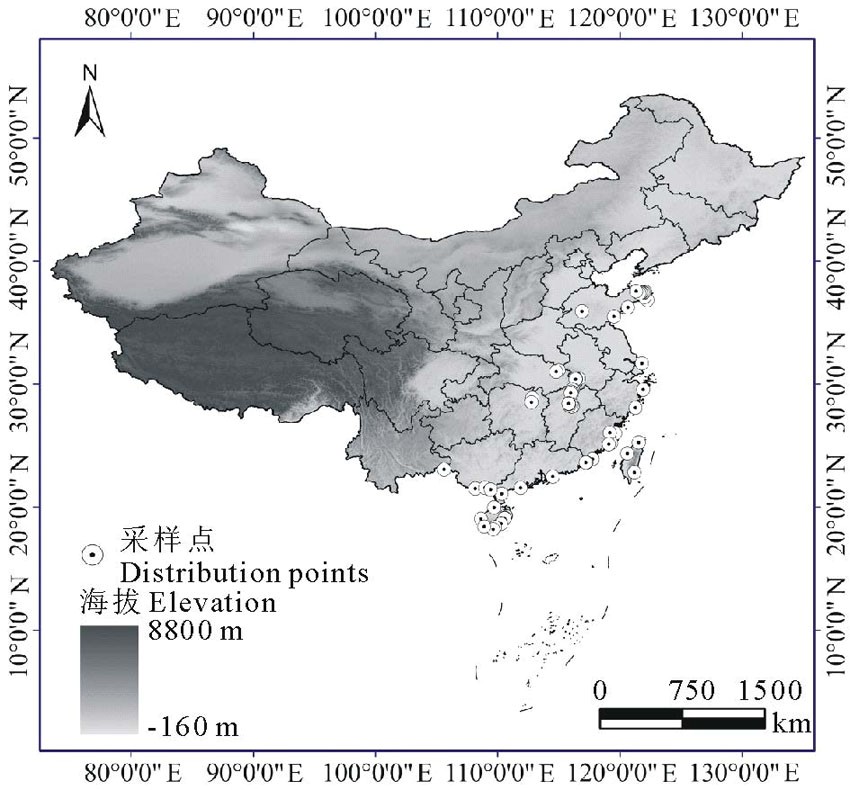 | 图1 已知单叶蔓荆分布点的地理位置Fig.1 The known geographic locations of V. trifolia var.simplicifolia |
首先通过查阅标本和相关文献确定已知的单叶蔓荆分布位点, 由中国数字植物标本馆(http://www.cvh.org.cn)得到44个单叶蔓荆分布点的信息, 通过查阅文献[11, 15, 16]得到82个单叶蔓荆分布点的信息, 共获得126个单叶蔓荆种群分布点的信息。根据单叶蔓荆已知分布位点的信息(图1), 依据126个分布点的信息和空间分辨率为30″(1 km2)的数字高程模型(digital elevation model, DEM), 我们设定研究区为中国全境(图1), 即预测单叶蔓荆在我国全国范围内的潜在分布情况。
通过访问世界气候数据库(WORLDCLIM version 1.3, http://www.worldclim.org)可以得到气候环境数据, 该数据库收集了1950-2000年世界各地的气象站所记录的详细气象信息, 采用插值法生成空间分辨率为30″(1 km2)的全球气候栅格数据[17]。本研究区涉及4个排放场景(RCP2.6、RCP4.5、RCP6.0和RCP8.5)。与前人研究经常采用的排放情景(special report on emissions scenarios, SRES)相比[18, 19], RCPs情景更加详细考虑了应对气候变化的各种策略对未来温室气体排放的影响, 更加科学地描述了未来气候变化的预估结果[20]。本研究选取温度、降水量等相关的22个对物种分布有重要影响的环境因子变量[21](表1), 进一步得到22个环境因子1950-2000年的年平均图层数据。
| 表1 用于MaxEnt模型中的环境因子变量 Table 1 The environment variables in MaxEnt |
选用基于最大熵理论的MaxEnt模型来模拟气候变化下单叶蔓荆潜在生境分布, 试验时间为2016年3月, 实验重复10次。MaxEnt模型具有较高的预测精度与良好的稳定性, 在物种分布模型中具有一定的竞争力[22]。
将单叶蔓荆分布数据和22个环境数据导入MaxEnt3.3.3, 从已知的分布点随机选取75%作为训练集来训练模型, 25%的分布点作为验证集来验证。模型输出数据格式为ASC图层, 图层中每个点的值代表单叶蔓荆在该区域对环境的适生性, 取值范围为[0.1], 然后利用ArcGIS 9.3软件将ASC数据转换成栅格格式。
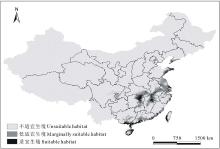
运用软件ArcGIS 9.3对栅格数据文件进行重分类, 生境适应性取值范围为0到1之间, 根据适宜生境评价指数[23, 24, 25]进行评价, 将研究区单叶蔓荆生境划分为3类(图2), 即不适宜生境(P< 0.25), 低适宜生境(0.25≤ P< 0.50), 适宜生境(0.50≤ P≤ 1.00)。利用ArcGIS 9.3软件, 可以得到研究区单叶蔓荆潜在空间分布具体结果(表2)。
基于MaxEnt3.3.3, 利用22个环境因子对单叶蔓荆潜在分布建立模型。在环境参数设置中开启刀切法(Jackknife), MaxEnt模型会计算22个评价因子对预测的贡献率。具体就是在模型建立过程中, 首先依次将每一个环境变量分别去除, 然后对剩余的21个环境变量建立模型, 最后对所有的环境变量建立模型并进行计算, 接下来分析被去除的环境变量与遗漏误差(omission error)之间的相关性, 如果被去除的环境变量导致遗漏误差显著提高, 则表明该环境变量对模型的预测结果有显著的影响[26]。
采用ROC(receiver operating characteristic curve)进行模型精度的验证。ROC曲线的具体做法如下:以预测值作为可能的判断阈值, 然后计算灵敏度和特异度[27], 以1减去特异度的值(1-specificity)为横坐标, 以灵敏度(1-omission rate)为纵坐标, 然后就可以绘制出ROC曲线, ROC曲线与横坐标围成的图形的面积称为AUC(area under ROC curve), 取值范围为[0, 1]。
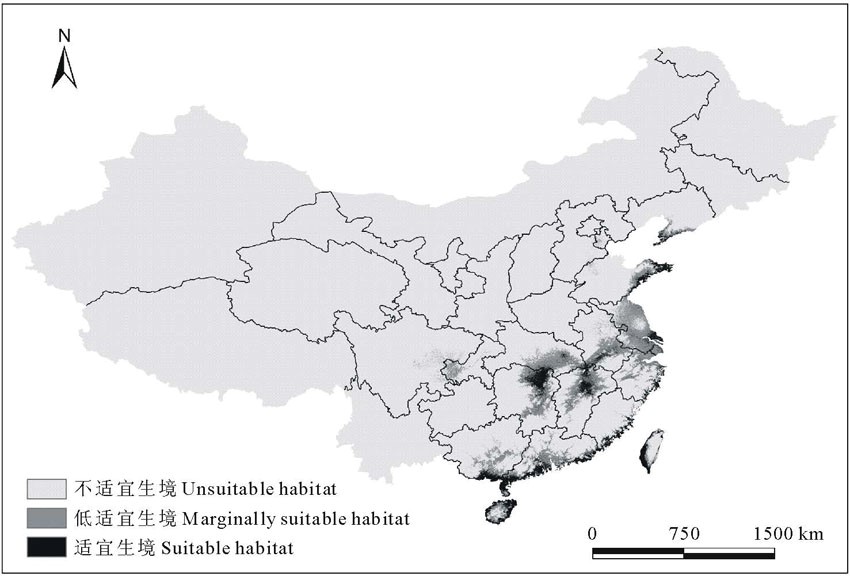 | 图2 单叶蔓荆的当前地理分布情况Fig.2 The current geographic distribution of V. trifolia var.simplicifolia |
| 表2 研究区各省市单叶蔓荆不同等级适生区面积及其百分比 Table 2 Areas and percentage of habitat suitability distribution of V. trifolia var.simplicifolia in different provinces and cities |
AUC可以反映模型的预测效果, 其值越大表示模型预测精度越高, 反之说明模型预测效果越差。由于AUC值不受诊断阈值影响, 并且对物种不敏感, 目前被公认为是最优的模型预测指标[28]。ROC曲线的评估准则为:AUC处于0.5~0.6之间时, 认为建立的模型失败; AUC处于0.6~0.7之间时, 认为建立的模型预测效果较差, 勉强可以接受; AUC处于0.7~0.8之间时, 认为模型一般, 可以接受; AUC处于0.8~0.9之间时, 认为建立的模型较好; AUC处于0.9~1.0之间时, 可以认为建立的模型很好, 预测效果是令人满意的[28]。
由图2和表2可知, 单叶蔓荆在研究区的适生范围主要集中在我国沿海以及内陆的中部。适宜生境的沿海区域包括海南省四周、广西壮族自治区的防城港东南部、北海市以及钦州市南部, 广东省的湛江市、茂名市南部、阳江市南部、江门市、中山市、深圳市南部、汕头市、汕尾市海丰县、揭阳市东南部、潮州市东部, 福建省的漳州市东南部、厦门市、莆田市的东部和福州市的东部, 台湾西部沿海地区, 浙江省的温州市东部、台州市东南部和宁波市东部部分地区, 江苏省的南通市和无锡市东北部, 上海市的东部地区, 山东省的威海市、青岛市南部和日照市东部, 辽宁省的大连市南部; 内陆包括湖南省的岳阳市、长沙市西部和湘潭市北部, 湖北省的武汉市东北部, 江西省的南昌市、临川市北部和九江市东部, 安徽省的贵池市西北部。
低适宜生境的沿海附近包括海南省的内陆部分地区, 广西壮族自治区钦州市中部和北部、南宁市、贵港市和合山市, 广东省的茂名市北部、佛山市、广州市、东莞市、惠州市、深圳市北部和揭阳市北部, 浙江省的绍兴市南部、宁波市南部、嘉兴市和湖州市, 江苏省的苏州市、常州市、无锡市西南部、镇江市、泰州市、扬州市、淮阴市和盐城市北部, 山东省的青岛市北部, 湖南省的衡阳市和株洲市南部; 内陆包括湖北省的仙桃市和孝感市南部, 安徽省的马鞍山市和巢湖市, 江西省的吉安市和宜春市东部。
分别对三级适宜生境的面积进行计算和统计, 可知单叶蔓荆的适宜生境面积、低适宜生境面积和不适宜生境面积分别占研究区总面积的5.03%、8.86%和86.11%。通过分析结果可知, 在所研究的区域内, 单叶蔓荆的适宜生境比例偏小, 可见单叶蔓荆的生长区域狭窄。
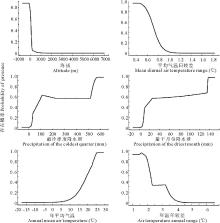
研究结果表明, 在所选择的22个环境因子中, 主要因子的贡献率为:海拔最高为41.3%, 平均气温日较差、最冷季度降水量和最干月份降水量贡献率分别为24.8%、10.1%和8.5%, 年平均气温和年温年较差贡献率分别为4.1%和3.0%。只列出了贡献率最高的前6个因子, 它们的累积贡献率就高达91.8%。为了进一步分析气候因子对单叶蔓荆分布的影响, 将上述影响最大的6个因子分别导入到MaxEn模型中, 建立单因子模型, 同时绘制出单变量响应曲线, 即单叶蔓荆地理分布概率与主导气候因子的关系(图3), 然后分别计算各主导气候因子的阈值(存在概率> 0.3):BIO(海拔)小于30 m, BIO2(平均气温日较差)小于0.75 ℃, BIO19(最冷季度降水量)大于50 mm, BIO14(最干月份降水量)大于10 mm, BIO1(年平均气温)大于16 ℃, BIO7(年温年较差)小于3.2 ℃。
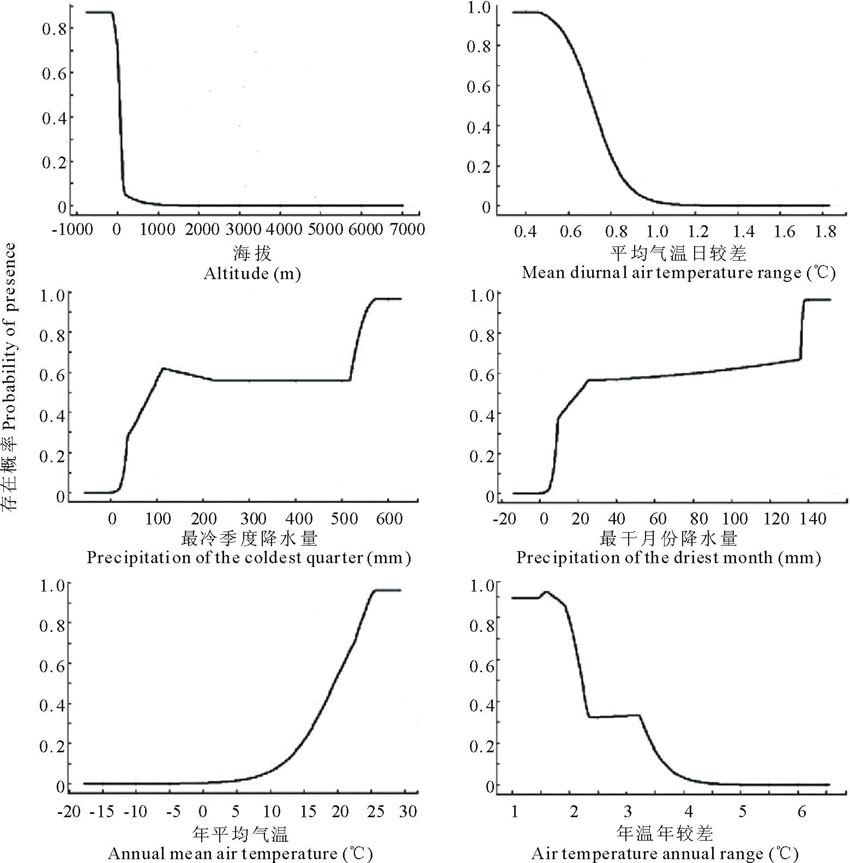 | 图3 6个主导气候因子与单叶蔓荆生存概率的关系Fig.3 Probability of survival between six dominant climate factors and V. trifolia var.simplicifolia |
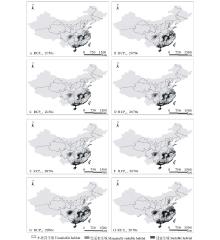
在不同的气候环境下, 利用MaxEnt模型模拟了单叶蔓荆的未来潜在地理分布。将研究区单叶蔓荆未来气候条件下生境适宜性分为3类, 可以得到研究区单叶蔓荆潜在空间分布(图4)和不同气候变化情景下研究区单叶蔓荆适宜生境和低适宜面积百分比(图5)。
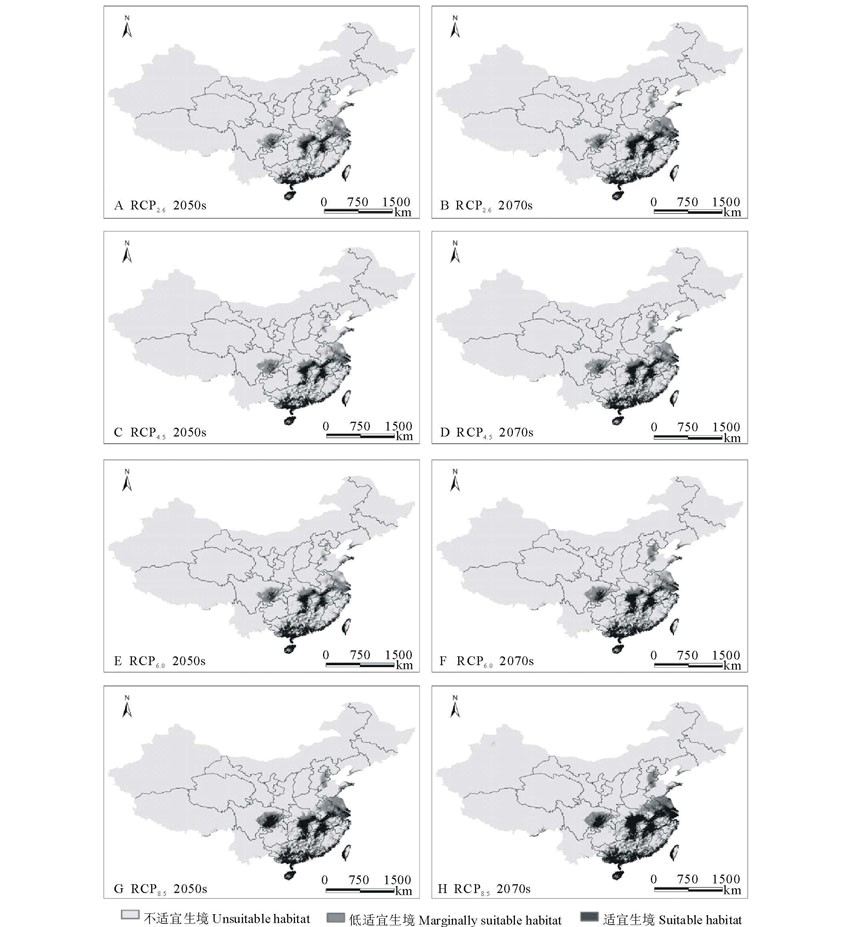 | 图4 不同气候变化情景下单叶蔓荆生境适应性分布 A和B、C和D、E和F及G和H分别表示在RCP2.6、RCP4.5、RCP6.0和RCP8.5气候情景下21世纪50和70年代。Fig.4 Distribution of habitat suitability for V. trifolia var.simplicifolia under different climate change scenarios A and B, C and D, E and F, G and H represent the RCP2.6, RCP4.5, RCP6.0 and RCP8.5 climate scenario in 2050s and 2070s respectively. |
由图4和图5可知, 在未来RCP8.5、RCP6.0、RCP4.5和RCP2.6四种情景下, 单叶蔓荆在研究区的适宜生境(评价指数0.50~1.00)增加程度都不相同。当前气候环境下单叶蔓荆的适宜生境比例为5.03%, 由模型预测结果可以看出, 在RCP2.6、RCP4.5、RCP6.0和RCP8.5四种情景下至21世纪50年代, 预测的适宜生境面积分别占研究区总面积的15.88%、17.00%、17.59%和23.11%。到21世纪70年代这4种情景下, 研究区单叶蔓荆适宜生境也均有一定幅度的增加, 并且在每种气候下增加的幅度都要大于21世纪50年代, 分别占研究区总面积的21.22%、22.21%、24.57%和30.66%。当前气候环境下单叶蔓荆的低适宜生境比例(评价指数0.25~0.50)为8.86%, 由模型预测结果可知, 在未来4种气候条件下单叶蔓荆的低适宜生境比例变化范围为10.31%~14.66%, 相对变化较小, 并且在RCP2.6、RCP4.5、RCP6.0和RCP8.5情景下到21世纪50和70年代, 该类生境的面积比例也都有一定程度的增加, 其中50年代分别占研究区总面积的11.09%、10.31%、11.53%和12.96%, 在70年代分别占研究区总面积的11.85%、12.07%、13.99%和14.66%。在空间分布上, 由图4及图5可以看出, 在不同气候变化条件下, 单叶蔓荆的适宜生境和低适宜生境的范围及几何中心均由沿海地区向内陆扩散; 内陆地区湖南和江西两省的适宜生境比例增长较快, 尤其在四川境内, 当前只有很小比例的低适宜生境, 随着气候的变化, 低适宜生境面积有所上升, 并且适宜生境开始出现且增长速度较快。
 | 图5 不同气候变化情景下研究区单叶蔓荆各种适生区面积百分比Fig.5 Percentage of areas of varying habitat suitability of V. trifolia var.simplicifolia under different climate change scenarios in the study area |
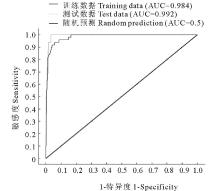
图6为本研究根据建立的模型所得到的ROC曲线。由图可以看出, 训练数据和测试数据的ROC曲线均远离随机分布模型的ROC曲线, 并且均靠向左上方, 表明曲线下的面积值较大接近1, 即AUC值较大。训练数据和测试数据的AUC值非常接近, 分别为0.984和0.992, 它们都显著大于随机分布模型的AUC值(0.5)。 通过AUC分析可知, MaxEnt模型对于单叶蔓荆潜在分布范围的预测具有较高的准确性。
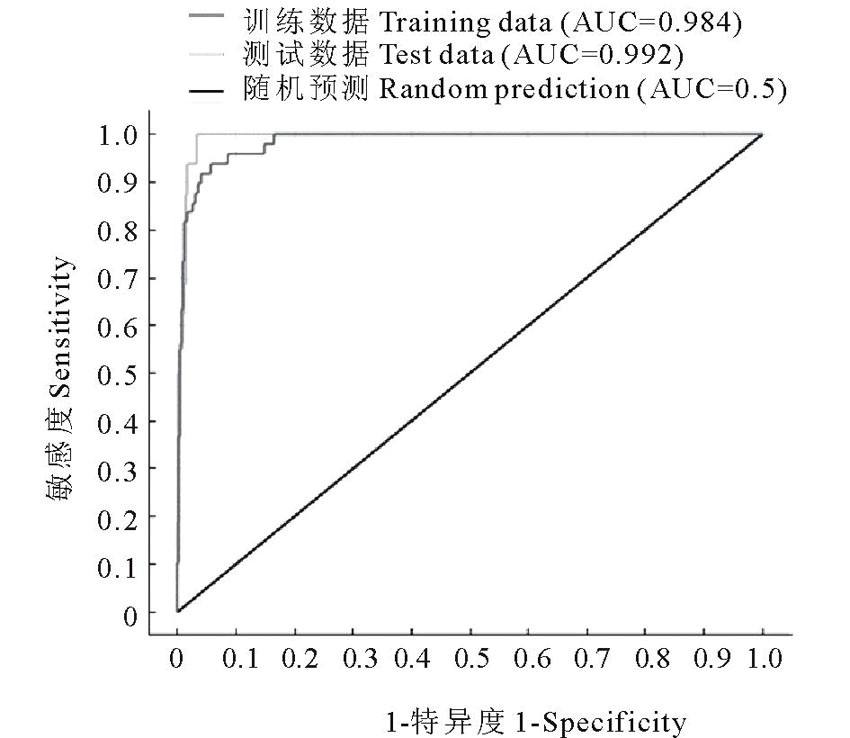 | 图6 ROC分析法检验MaxEnt预测的结果所得到的AUC值Fig.6 The AUC by using ROC methods to test the results of MaxEnt |
本研究利用MaxEnt模型对单叶蔓荆在当前气候条件下的适生区进行了研究, 但仅探讨了气候因素对单叶蔓荆分布格局的影响, 由于物种生长分布受诸多因素限制, 土壤环境、生物间的相互作用等都与物种分布有关, 进一步的研究可考虑土壤条件、局地生境对单叶蔓荆分布范围的影响。
本研究利用MaxEnt模型定量展示了在未来RCP2.6、RCP4.5、RCP6.0和RCP8.5四种情景下研究区单叶蔓荆潜在的分布区域。研究结果表明, 在不同的气候条件下单叶蔓荆的适宜生境面积都将有所扩大, 同时在研究区涉及的各省、市中, 沿海岸以及内陆江西、湖南、安徽等省为单叶蔓荆未来主要的潜在适生区域, 单叶蔓荆的适宜生境和低适宜生境的范围及几何中心均由沿海地区向内陆扩散; 内陆地区湖南和江西两省的适宜生境比例增长较快。研究还发现, 在四川境内, 当前只有很小比例的低适宜生境, 并没有适宜生境, 但是随着气候的变化, 低适宜生境面积有所上升, 并且适宜生境开始出现且增长速度较快, 四川也逐渐成为单叶蔓荆未来主要潜在的适宜生长区域。
根据评价因子贡献率的结果, BIO(海拔)这一评价因子贡献率就高达41.3%, 说明海拔是影响单叶蔓荆适生区分布的决定性因子, 梁芳等[11]也提到, 单叶蔓荆主要分布在海拔为2~21 m(只有少数的在2000 m以上)地区, 对海拔要求极为苛刻, 本研究也证实了这一点, 单叶蔓荆适宜生存的海拔范围小于30 m, 与实际比较吻合。本研究还显示, 气温差评价因子的累计贡献率未起到最重要的作用, 说明气温差不是制约单叶蔓荆分布的关键因子。BIO19与BIO14两个评价因子的贡献率之和为19.6%, 它们都是与降水量相关的因子, 这说明了降水量也是影响单叶蔓荆适生区分布的一个重要的因子。
单叶蔓荆主要分布于热带与亚热带地区, 在粗沙地、石砾地和沙质草地上均能生长, 多生长于海滨、湖畔和沙滩等近水处及阳光充足地带。该植物主要集中在沿海海滨沙地, 对于沙质较少的土壤适应能力不强[11]。本研究对单叶蔓荆未来的分布预测也与上述描述的单叶蔓荆的分布范围相一致, 说明气候变化情景下仍然可靠。
单叶蔓荆是一种药用价值和生态价值都很高的药材, 预测其地理分布对于其资源调查、保护开发和资源可持续利用具有重要的意义。根据预测结果, 应该有目的地选择适宜生境和低适宜生境区域, 对单叶蔓荆进行采样开展野生资源调查研究。有效选取野生抚育GAP(good agricultural practice)基地地理位置, 充分考虑其未来适宜生境的区域, 尤其要重视未来单叶蔓荆潜在适宜分布的四川等地区, 对当地加大宣传保护力度, 展开相关措施, 提倡野生抚育, 加以有效的人工管理和合理的投资, 扩大单叶蔓荆的种群数量, 进而对其珍贵的价值进行可持续利用。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|