





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
黄土区封育和放牧草地物种多样性和地上生物量对氮素添加的响应
[赵洁1  , 李伟
, 李伟2 , 井光花2 , 魏琳2 , 程积民1, 2, * ]
, 李伟]
|
|


作者简介:赵洁(1992-),女,陕西西安人,硕士。E-mail:1440039188@qq.com
氮素添加对草地生态系统地上生物量具有普遍促进作用,但对解释引起物种多样性丧失的机制仍存在较大争议。本研究以黄土高原半干旱区长期封育和连续放牧草地为对象,通过设置不同氮素添加水平来探索两种典型的草地管理方式下群落地上生物量、物种多样性、群落组分结构对氮素添加的响应及其异同以及物种多样性与地上生物量的关系,并研究这些响应的潜在生态学机制。结果显示,1)氮素添加显著提高长期封育草地地上生物量的同时降低了物种多样性;放牧草地群落地上生物量、物种多样性对氮素添加的响应与封育草地相同,但其响应程度均小于封育施肥草地;2)草地群落物种多样性随地表凋落物生物量增加而降低,随植被透光率增加而升高; 3)封育施肥草地Shannon-Wiener多样性指数与地上生物量存在显著线性负相关关系,而放牧施肥草地的物种多样性与地上生物量不存在显著的相关关系。以上结果表明放牧能够减缓施肥对物种多样性的负效应,在轻微降低物种多样性的前提下显著提高群落地上生物量,是一种合理的草地管理方式。
Nitrogen addition can increase the aboveground biomass of the plant community in grasslands. However, it can also lead to a loss of species diversity, but the mechanisms involved in this process are unclear. The objective of this study was to explore the effects of nitrogen addition on aboveground biomass, species diversity, community composition, and the relationship between species richness and aboveground biomass in long-term fenced grassland and grazed grassland on the semi-arid Loess Plateau. We focused on the differences and similarities in the grassland responses under these two typical management systems, and tried to identify the the ecological mechanisms underlying these responses. We added nitrogen at different rates [0, 5, 10, 20, 40, 80 g CO(NH2)2/(m2·yr)] to long-term fenced grassland and grazed grassland. After 3 years of treatment, we measured aboveground biomass, calculated species diversity indexes (species richness, Shannon-Wiener index, and Pielou index) and analyzed the relationship between species diversity and productivity using regression analyses. The results showed that, in the long-term fenced grassland, nitrogen addition significantly increased the aboveground biomass and decreased species diversity, while the grazed grassland showed similar but weaker responses to nitrogen addition. The species diversity in fenced and fertilized grassland decreased as the litter biomass increased, and also increased with greater light penetration. In comparison, species diversity in grazed and fertilized grassland decreased slowly and did not change significantly with greater light penetration. The Shannon-Wiener index was significantly negatively correlated with above-ground productivity in fenced and fertilized grassland, but no such relationship existed in grazed grassland. These results indicated that grazing can relieve the negative effect of fertilization on species diversity. Therefore, grazing is a reasonable grassland management method to retain species diversity while increasing the community aboveground biomass by fertilization.
多数研究表明氮素添加在提高生物量的同时会造成草地生态系统物种多样性下降[1, 2]。氮素添加对草地地上生物量的影响在年限和氮素水平上可能存在阈值, 草地地上生物量在到达此阈值时达到峰值, 之后氮素对地上生物量促进效应将减小甚至转为负向影响[1, 3, 4] 。Stevens等[5]根据逐步回归预测氮沉降速率为0.25 g N/(m2· yr)时, 平均每4 m2将有一个物种消失。但对引起物种多样性下降的原因尚存在争议, 目前人们提出的解释机制主要有以下几种:光竞争、凋落物积累、集合水平自疏假说、铵中毒与土壤酸化[6, 7, 8, 9, 10]。其中光竞争的理论认为, 氮素的增加解除了生态系统对氮素的限制, 嗜氮植物快速生长, 在养分吸收、高度、盖度上占得优势, 荫蔽一些非嗜氮、生长较慢或矮小的物种, 降低物种多样性。即种间竞争由地下对养分的竞争转为地上对光的竞争, 植物对养分和光竞争力的差异决定了最终群落物种组成格局[11]。另一方面, 氮素增加后伴随着植物生产力提高, 凋落物积累量也增加, 影响了一些种子萌发和幼苗生长, 也是物种多样性降低的原因之一[12]。植物群落与环境之间的相互作用极其复杂多面, 上述研究及假说虽然可以从一定程度上解释物种多样性对施肥的响应, 但是到目前还没有较为统一的认识, 氮沉降导致物种多样性丧失的机制还有很大的研究空间。
尽管国内外学者对氮沉降已经进行了许多研究[13, 14, 15, 16, 17], 但对氮素和水分限制的黄土高原半干旱草地生态系统的研究还相对较少, 且其对氮素添加的响应可能更加敏感[18]。同时近年来也有少数研究表明放牧可能会减缓施肥对物种多样性的改变[3, 19]。基于上述原因及前人研究基础选取黄土高原典型草原长期封育和连续放牧草地为研究对象, 通过氮素添加实验研究两种管理方式下草地地上生物量和物种多样性对氮素添加的响应, 为黄土高原已恢复天然草地的后续合理利用以及我国生态草牧业的可持续发展提供理论依据。本研究的主要目标是:1)比较封育和放牧草地地上生物量和物种多样性随着不同氮素添加水平变化的异同; 2)研究放牧和施肥对物种多样性的影响及原因; 3)探索黄土高原典型草原上物种多样性与地上生物量的关系。
研究地位于宁夏固原市东北部的云雾山国家草地自然保护区实验区(106° 21'-106° 27' E, 36° 10'-36° 17' N), 海拔1800~2100 m。该区域气候属于中温带半干旱区, 年平均气温5 ℃, 最冷月1月(平均气温-14 ℃), 最热月7月(22~25 ℃), 年降水量400~450 mm, 降雨季节分配不均, 全年65%~75%的降水集中在7-9月, 蒸发量1330~1640 mm。年日照时数为2500 h, 无霜期112~140 d。
保护区植被主要是温带典型草原, 其建群种和优势种以白莲蒿(Artemisia sacrorum)、大针茅(Stipa grandis)为主, 伴生种类型以猪毛蒿(A. scoparia)、百里香(Thymus mongolicus)、星毛委陵菜(Potentilla acaulis)为主。
实验选取长期封育和连续放牧两种管理方式下的草地, 采用随机区组设计[3], 自2013年起进行长期氮素添加实验, 于每年生长季初期即4月底施肥, 为提高肥料利用率于小雨天施肥, 按实验设计的施用量用手撒施。其中封育草地封育30年; 放牧草地连续30年进行放牧实验, 放牧强度为4只羊/hm2, 放牧期为6-8月, 每月放牧1次, 每次10 d, 放牧时间为早上6点将家畜赶入放牧区自由采食, 下午6点赶回畜圈休息。以尿素作为肥料, 6个氮素添加水平依次为:CK, 0 g/(m2· yr); N5, 5 g/(m2· yr); N10, 10 g/(m2· yr); N20, 20 g/(m2· yr); N40, 40 g/(m2· yr); N80, 80 g/(m2· yr), 等价于氮素分别为0、2.34、4.67、9.34、18.68、37.35 g/(m2· yr), 其中N5为低氮水平, N10 和N20为中氮水平, N40和 N80为高氮水平。每个水平6个重复, 每块样地上设置36个4 m× 4 m小区, 各区间有2 m缓冲带。
于2015年在植物生长旺期即8月上旬在每个小区内随机作0.5 m× 0.5 m的样方进行植被群落调查, 包括物种组成、数目、各物种植株平均高度、多度及盖度。剪取植物地上部分并按物种分类装入信封, 同时收集样方内凋落物带回实验室80 ℃烘干至恒重, 分别测定植物地上生物量与凋落物生物量。
在天气晴朗的日子用数字照度仪(LX-1010B, 中国)在上午11点至下午13点间测定群落光辐射。在每个小区内随机选取3处分别测定冠层光辐射和地面光辐射。
功能群将具有相似生活型、特性或对环境因子响应类似的植物进行分组, 基于功能群的研究方法是评估物种和群落对环境因子改变所做出响应的有效手段[20]。参考王谢等[21]的方法按可食性进行功能群划分并计算各功能群地上生物量占总地上生物量的百分比以表征不同物种在群落中的功能。将植物群落以可食性划分为4个组:可食禾草类(grass species group, GG), 包含莎草科干生苔草(Carex aridula)、非禾本科可食草类(forbs species group, FG)、可食性低劣或有毒害草类(noxious species group, NG)、豆科类(leguminous species group, LG)。
本研究计算物种重要值、α 多样性和植被透光率。α 多样性包括物种丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度指数。计算公式如下:
(1) 物种重要值
物种重要值=(相对生物量+相对盖度+相对高度)× 100/3
(2) α 多样性指数
Patrick丰富度指数:R=S
Shannon-Wiener多样性指数:H'=-∑ Piln Pi
Pielou均匀度指数:J=H'/ln S'
式中: S为样方的平均物种数; Pi为相对多度, 即物种i的地上生物量占样方内所有物种地上生物量百分比; S'为各样方中的物种数。
(3) 植被透光率
植被透光率=(地表光合有效辐射/冠层光合有效辐射)× 100%
利用R软件进行数据整理、分析及作图。采用双因素方差(Two-way ANOVA)分析草地管理方式和氮素添加对草地地上生物量和物种多样性的影响, 用Tukey法进行多重比较, 进一步分析封育草地和放牧草地各施肥水平间及同一施肥水平下封育草地和放牧草地地上生物量与物种多样性差异。采用Pearson法检验物种多样性与地上生物量间相关性, 并做一般线性回归。用Shapiro-Wilk test和Bartlett test检验数据正态性和方差齐性。
双因素方差分析结果显示(表1), 草地管理方式和氮素添加对草地地上生物量皆有显著影响, 但其交互效应并不显著。与封育草地相比, 放牧使地上生物量下降27.7%(236.72 g/m2到171.23 g/m2); 施肥后封育草地地上生物量总体上呈低氮急速上升— 下降— 中氮缓慢上升— 高氮下降的双峰趋势, 各氮素水平下地上生物量皆比未施肥(CK)时高, 且在N5水平时达到最大, 多重比较结果得出其与CK、N10、N80水平时地上生物量差异显著(P< 0.01); 在放牧施肥草地, 地上生物量的变化趋势与封育施肥草地相似, 与未施肥(CK)相比, 氮素浓度为N5、 N10、 N80水平时地上生物量显著增高(P< 0.05), 且在N10水平下地上生物量达到最大(图1); 两样地地上生物量在N5水平下差异显著(P< 0.05)。
| 表1 草地管理方式和氮素添加对草地地上生物量和物种多样性影响的双因素方差分析 Table 1 Two-way ANOVA results of the influence of grazing and nitrogen addition to aboveground biomass and species diversity in grassland |
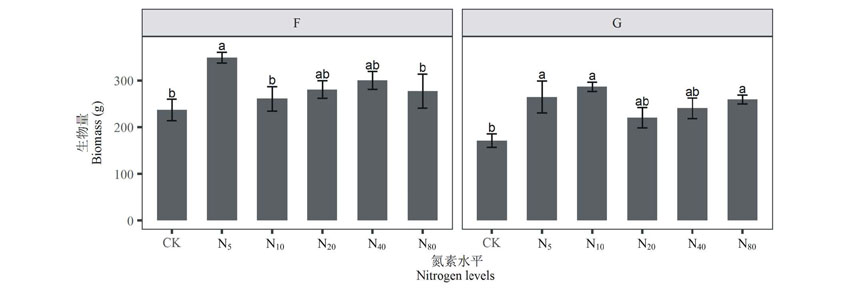 | 图1 封育草地(F)和放牧草地(G)地上生物量对氮素添加的响应(平均值± 标准误差) CK、N5、N10、N20、N40、N80指氮素添加量分别为0、5、10、20、40、80 g CO(NH2)2/(m2· yr)。不同小写字母表示封育施肥草地和放牧施肥草地各施肥水平间差异显著(P< 0.05)。下同The same below.Fig.1 Responses of community aboveground biomass to nitrogen addition (mean± SE) CK, N5, N10, N20, N40, N80 represent nitrogen addition levels of 0, 5, 10, 20, 40, 80 g CO(NH2)2/(m2· yr). Different small letters indicate significant differences among the nitrogen addition levels within the two sites (P< 0.05). |
依照物种重要值> 10%为优势物种, > 5%为亚优势种, < 5%为伴生种的标准划分群落物种[4] 。由图2可见, 封育施肥草地的优势物种甘青针茅、大针茅、白莲蒿等较高大的嗜氮植物重要值随氮素水平增加而增加, 个体高度较低的蓬子菜则呈降低趋势; 个体高度较为中等的亚优势物种赖草、披碱草、火绒草、翼茎风毛菊等重要值在低、中氮水平增加, 在高氮水平下又降低, 群落下层的甘菊、多毛并头黄芩、百里香等低矮的物种重要值则表现为连续下降; 重要值< 5%的一些物种, 如本氏针茅、冰草、蚓果芥等几乎可看做偶见种, 随着氮素增加自群落中消失。
在放牧施肥草地中情况大有不同, 优势物种中不仅出现了光稃茅香, 更值得注意的是出现了在封育施肥草地中作为伴生种的百里香和干生苔草; 甘菊作为亚优势种出现, 与封育施肥草地不同的是其重要值随氮素增加而连续增加, 在封育施肥草地中为偶见种的甘肃蒿也在各氮素水平出现; 多种封育施肥草地中不存在的物种, 如西山委陵菜、西藏点地梅、星毛委陵菜、青海苜蓿等个体高度很低的物种成为放牧施肥草地的伴生种。
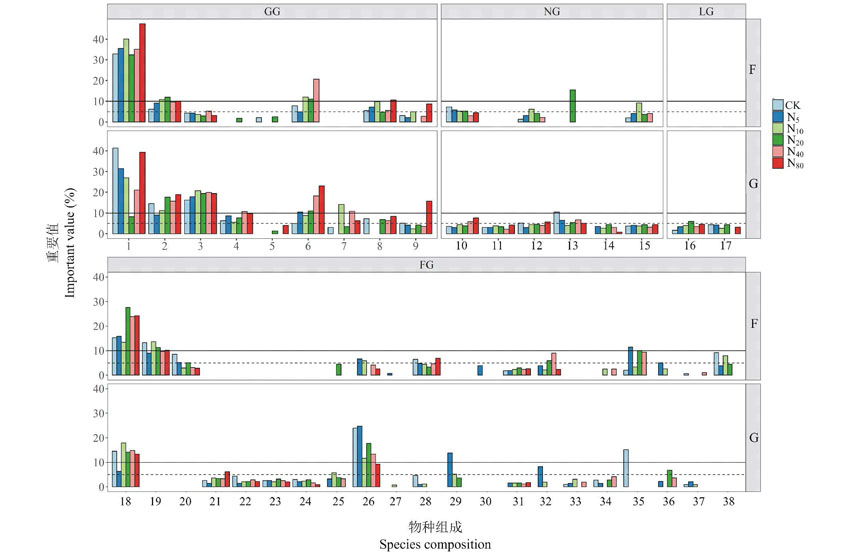 | 图2 封育草地(F)和放牧草地(G)群落物种组成及重要值对氮素添加的响应 按可食性分组的功能群, 其中GG为可食禾草类(包括莎草科)、FG为非禾本科可食草类、NG为可食性低劣或有毒害草类、LG为豆科类。Functional group divided by edibility, GG (grass and sedge species group), FG (forbs species group), NG (noxious species group), LG (leguminous species group). 下同 The same below.1: 甘青针茅 Stipa przewalskyi; 2: 大针茅 Stipa grandis; 3: 赖草 Leymus secalinus; 4: 干生苔草 Carex aridula; 5: 光稃茅香 Anthoxanthum glabrum; 6:披碱草 Elymus excelsus; 7: 本氏针茅 Stipa bungeana; 8: 早熟禾 Poa sphondylodes; 9: 冰草 Agropyron cristatum; 10: 甘肃蒿 Artemisia gansuensis; 11: 甘菊 Chrysanthemum lavandulifolium; 12: 火绒草 Leontopodium leontopodioides; 13: 阿尔泰狗娃花 Heteropappus altaicus; 14: 狼毒 Stellera chamaejasme; 15: 黄毛棘豆 Oxytropis ochrantha; 16: 青海苜蓿 Medicago archiducis-nicolaii; 17: 小果黄耆 Astragalus zacharensis; 18: 白莲蒿 Artemisia sacrorum; 19: 百里香 Thymus mongolicus; 20: 蓬子菜 Viola dissecta; 21: 翼茎风毛菊 Saussurea japonica var. pteroclada; 22: 丝毛飞廉 Carduus crispus; 23: 长柱沙参 Adenophora stenanthina; 24: 细叶沙参 Adenophora capillaria subsp. paniculata; 25: 多毛并头黄芩 Scutellaria scordifolia var. villosissima; 26: 二裂委陵菜 Potentilla bifurca; 27: 白花枝子花 Dracocephalum heterophyllum; 28: 蚓果芥 Neotorularia humilis; 29: 急折百蕊草 Thesium refractum; 30: 细叶远志 Polygala sibirica; 31: 野韭 Allium ramosum; 32: 北柴胡 Bupleurum chinense; 33: 西山委陵菜 Potentilla sischanensis; 34: 西藏点地梅 Androsace mariae; 35: 裂叶堇菜 Viola dissecta; 36: 星毛委陵菜 Potentilla acaulis; 37: 早开堇菜 Viola prionantha; 38: 北方还阳参 Crepis crocea.Fig.2 Responses of species composition and important value to nitrogen addition |
以可食性分组的功能群在两样地中皆以可食禾草(包括莎草科)为主, 其地上生物量占地上总生物量比例平均值分别为59.36%和50.74%。在封育施肥草地中, 可食禾草地上生物量所占比重随氮素水平的变化增加不明显, 甚至在N20水平下降22%(图3), 结合图2可知处于优势地位的甘青针茅在N20水平下大幅的下降造成了这种结果; 非禾本科可食草类和可食性低劣或有毒害草类地上生物量皆呈先增加再降低的趋势, 氮素水平间差异不显著; 封育施肥草地中未出现豆科类草。放牧施肥草地中, 可食禾草地上生物量所占比重随着氮素水平增加连续增加, 在N80时与CK形成显著差异(P< 0.01), 增加1.39倍, 除了在封育施肥草地中也存在的大针茅、甘青针茅等禾本科草类, 干生苔草、本氏针茅、光稃茅香和早熟禾地上生物量亦明显增加(图2); 非禾本科可食草类的比重则呈降低趋势, CK、N5、N20水平下的地上生物量占比皆与N80差异显著(P< 0.01), 分别下降63.13%、65.58%、54.38%; 可食性低劣或有毒害草类地上生物量占比随氮素增加而降低, 但各处理间差异不显著; 与重要值分析相对应, 放牧施肥草地出现了可食性优良的豆科牧草(图3)。
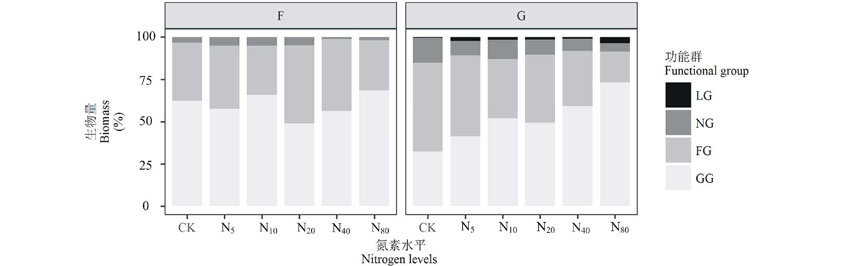 | 图3 封育草地(F)和放牧草地(G)各功能群地上生物量对氮素添加的响应Fig.3 Responses of aboveground biomass of functional group to nitrogen addition |
双因素方差分析结果显示(表1), 草地管理方式对3种多样性指数都具有显著影响, 氮素添加对物种丰富度具有显著影响, 但对Shannon-Wiener多样性指数和Pielou均匀度指数无显著影响, 且交互作用对3种指数亦无显著影响。由图4可见, 3种多样性指数在封育草地中随施肥水平的递增皆呈连续下降趋势, 但各处理间差异不显著, 与CK相比, N80水平下的物种丰富度指数、Shannon-Wiener多样性指数和Pielou均匀度指数分别下降31.3%、32.64%、22.58%; 在放牧施肥草地中, 3种指数皆为先增加后降低, 其中物种丰富度指数和Shannon-Wiener多样性指数在N10水平时达到最大, 均匀度指数在N20水平时升至峰值。两样地的3种多样性指数分别在N10、N80、N80水平下差异显著(P< 0.05)。
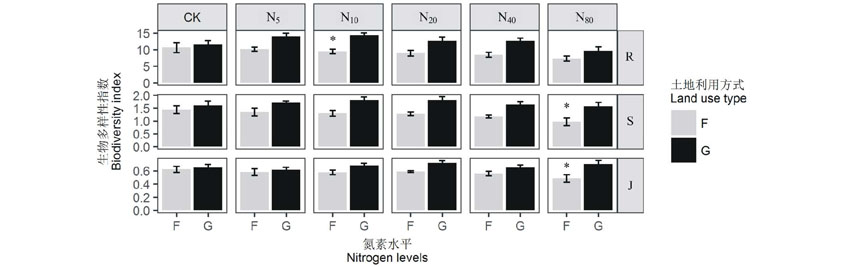 | 图4 封育草地(F)和放牧草地(G)α 多样性对氮素添加的响应 R、S、J分别代表物种丰富度、Shannon-Wiener多样性指数、Pielou均匀度指数。* 表示封育草地与放牧草地在同一处理下差异显著(P< 0.05)。 R, S, J represent species richness, Shannon-Wiener index and Pielou index. * indicates significant difference among fenced and grazed plots under the same treatment (P< 0.05).Fig.4 Responses of alpha diversity to nitrogen addition |
封育施肥草地的Shannon-Wiener多样性指数和地上生物量呈显著负相关(r=-0.47, P< 0.01), 在放牧施肥草地两变量并无显著相关性(图5)。
相关分析结果显示, Shannon-Wiener多样性指数与凋落物生物量呈显著负相关(r=-0.51, P< 0.001), 随凋落物生物量线性降低(图6); Shannon-Wiener多样性指数与植被透光率呈显著正相关(r=0.47, P< 0.001), 随植被透光率线性增加(图6)。
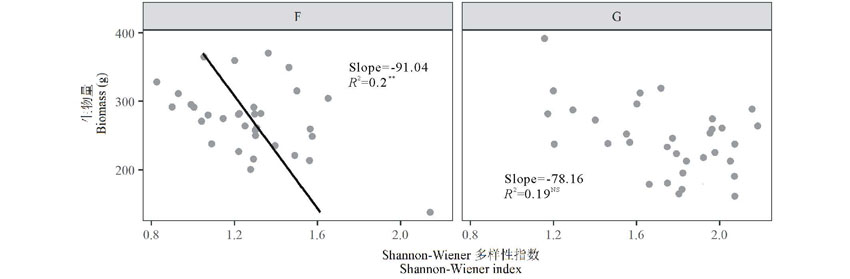 | 图5 氮素添加后封育草地(F)和放牧草地(G)物种多样性指数与地上生物量之间的关系 以Shannon-Wiener多样性指数为解释变量, 地上生物量为响应变量建立回归模型。Regression model was established by using Shannon-Wiener index as explanatory variable and aboveground biomass as response variable. 显著性水平为Significant differences are reported as: NS, P> 0.05; * , P< 0.05; * * , P< 0.01; * * * , P< 0.001; 下同 The same below.Fig.5 Relationship between biodiversity index and aboveground biomass after nitrogen addiction |
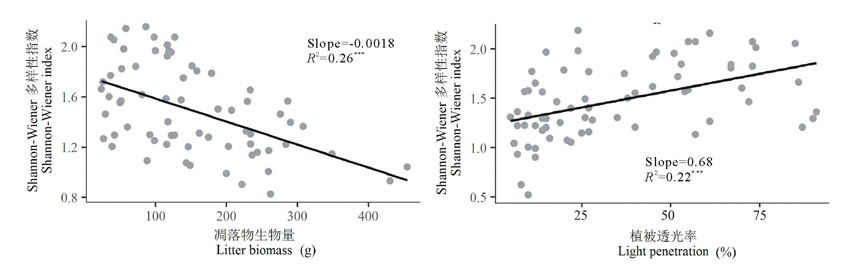 | 图6 氮素添加后物种多样性指数与凋落物生物量和植被透光率之间的关系 以Shannon-Wiener多样性指数为响应变量, 凋落物生物量、植被透光率为解释变量建立回归模型。Regression models were established by using Shannon-Wiener index as response variable and litter aboveground biomass, vegetation light penetration as explanatory variables.Fig.6 Relationship between biodiversity index and aboveground biomass, vegetation light penetration |
添加氮素后, 封育和放牧草地地上生物量与对照相比皆显著增加, 变化趋势大体上都为先增加后降低, 但二者在数量上有一定的差距(图1), 这与Bai等[3]的研究结果一致, 他认为两样地间生物量的差异源自于不同的植物-土壤反馈系统影响了植物对水的获取能力。在湿润的高寒草甸和干旱的荒漠草原进行氮素添加实验, 结果显示两群落的地上生物量分别在氮素浓度为23.34和0.35 g/m2时达到峰值[4, 22], 表明对位处干旱半干旱区的草地生态系统来说, 除了氮素, 水分也是植物生物量增加的限制因素之一[18]。陈静等[23]在科尔沁沙质草地实施的增雨和增氮实验发现, 单纯地添加水分或氮素对植被总生物量及各构件平均生物量无显著影响, 但在夏季增雨× 氮素添加处理下生物量则显著增加。
在本实验中, 封育施肥和放牧施肥草地地上生物量达到最高值时的氮素浓度分别为2.34和4.67 g/m2, 放牧施肥草地地上生物量到达峰值时的氮素浓度比封育施肥草地高, 表明封育草地地上生物量对氮素添加的响应更加敏感。封育草地中, 氮素添加前处于优势地位的甘青针茅、大针茅、白莲蒿等个体高度较高且根系发达的物种在添加氮素后依靠其生理特性可获得更多的营养和水分来生长繁殖, 其凋亡后成为营养丰富的凋落物, 一方面为植物、土壤微生物提供营养, 另一方面覆盖于地面减少水分散失, 起到一定的保水作用, 形成良性的水— 植物— 土壤反馈系统, 使地上生物量在氮素添加后快速增加。而放牧施肥草地则由于牲畜的啃食、踩踏凋落物积累量较少, 失去了保水和提供营养的凋落物层, 地上生物量的增长较封育施肥草地缓慢, 体现在与封育施肥草地地上生物量在数量上有一定差距且对氮素添加的响应较慢。
伴随氮素梯度增加带来的地上生物量增加同时也导致了凋落物生物量的积累, 而这可能是造成物种多样性降低的原因之一。Jensen等[24]移除了样方内的凋落物, 结果种子出苗率显著增加, 提高了物种丰富度并改变了物种组成, 表明凋落物在植物群落构建中起着关键性作用。Clark等[7]的凋落物移除实验显示, 群落中到达土壤表面的透射光增加了54%, 幼苗数量增加, 最终导致物种丰富度增加。根据以上及前人实验, 目前关于凋落物影响植物群落物种组成和物种多样性的机制有以下几点:1)阻碍种子出苗或幼苗建植(包括改变种子的物理环境导致萌发的关键因素缺失; 分泌化学物质改变化学环境抑制种子萌发; 形成机械屏障阻碍种子扩散); 2)削弱到达地面的光照强度, 影响幼苗或个体高度较低的植物获取光能; 3)占据裸露地面抑制了克隆植物无性繁殖[25]。
本实验封育施肥草地中, 随着氮素增加后凋落物的积累, 从功能群层面看, 禾本科草类受凋落物影响较小, 豆科草类受影响最大。从物种组成层面看, 一些个体高度较低的物种如蓬子菜、多毛并头黄芩、百里香等重要值随氮素增加而降低甚至从样方中消失(图2), 导致了物种多样性连续降低(图4); 放牧施肥草地中则由于牲畜的啃食与踩踏行为减小了凋落物积累量, 解除了凋落物对一些较低矮物种的限制, 出现了早熟禾、西藏点地梅、青海苜蓿等物种, 但氮素输入导致了禾本科植物地上生物量连续增高(图3), 凋落物也随之增高, 从而造成放牧施肥草地物种多样性先升高后下降的趋势, 但其物种多样性总体水平依然高于封育施肥草地。
氮素的增加解除了生态系统对氮素的限制, 封育施肥草地中对氮素敏感、根系密度高、处于优势地位的嗜氮物种, 如禾本科的赖草、大针茅, 非禾本科可食草类的白莲蒿快速生长, 进一步在养分吸收、高度、盖度上占得优势, 降低植被透光率, 荫蔽一些对氮素不敏感生长较慢或矮小的物种, 从而降低了物种多样性; 而放牧施肥草地较高的植被透光率利于较低矮植物的拓殖和存活, 一些封育施肥草地中不存在的豆科牧草如青海苜蓿和小果黄耆也相继出现, 故随着植被透光率增加, 更多的光能到达地表, 物种多样性也随之增加(图5)。表明种间竞争由植物对地下养分的竞争转为地上对光资源的竞争, 植物对养分和光竞争力的差异造成的生态位分化决定了最终群落物种组成格局[11, 26]。
故结合光竞争和凋落物积累对群落物种组成和物种多样性的影响可见, 氮素添加后封育施肥草地可能由于凋落物积累和植被对光资源竞争强度的增强导致了物种多样性下降, 而放牧施肥草地凋落物积累量较少, 也没有改变光竞争强度, 其物种多样性下降速度低于封育施肥草地物种, 故推论:放牧可减缓施肥对物种多样性的降低作用。更有研究指出适度的干扰可以提高物种丰富度, 改善种群的生态位配置及最大限度地利用环境资源[27]。
张杰琦等[22]和于丽等[28]在封育草地添加氮素的实验结果显示, 物种丰富度和Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数皆与地上生物量呈显著的线性负相关关系。与其相同, 本实验封育施肥草地中, Shannon-Wiener多样性指数与地上生物量存在显著负相关关系; 在放牧施肥草地中, Shannon-Wiener多样性指数与地上生物量无显著的相关关系(图6), 但Shannon-Wiener多样性指数随地上上生物量增加总体上呈下降趋势, 且比封育施肥草地缓慢。这一结果与Bai等[3]的结论一致。
自20世纪60年代起, 物种多样性与生产力的关系就是植物生态学家们极具兴趣且充满争议性的话题。一些学者认为物种多样性与生产力之间的关系随着研究尺度的变化有着较为普遍的模式, 并利用Meta分析总结出5种被广泛接受的物种丰富度— 生产力关系(SRPR):正相关, 负相关, 单峰, U型和不相关[29]。研究者们认为模式的多变性取决于研究尺度, 即SPRP是“ 尺度依赖性” 的[30]。Meta分析的结果显示, 在较大的尺度上这种关系更倾向于表现为正相关, 而在较小的尺度上单峰与U型模式则更加普遍。但也有学者对这种综合性的结果持质疑态度, Adler等[31]通过在五大洲的标准化取样方法分别在局域、区域和全球尺度探索物种丰富度和初级生产力之间的关系, 结果并未找到明确的、具有普遍性的关系模式, 认为Meta分析忽略了不同研究间方法论的差异, 不能够作为可靠的结论, 生产力并不能作为一个很好的指标来预测物种多样性的变化, 即在局域、区域和全球尺度不存在一致的生产力与物种多样性关系。
本实验亦未得出物种多样性与地上生物量间特定的模型。因为在复杂多变的生态环境中, 影响物种多样性的因素是多元化的, 在后续的研究中应综合分析降雨量, 土壤营养元素、pH、含水量, 微生物活动, 温度等因子。
通过对黄土高原半干旱区长期封育和连续放牧草地进行氮素添加实验, 研究结果表明:1)施肥显著提高了封育草地地上生物量, 但当地上生物量达到一定阈值时也导致了物种多样性降低, 改变群落物种组成, 这些改变可能会反过来影响地上生物量; 2)放牧可在显著提高地上生物量的同时减小物种多样性降低程度。作为生态系统稳定性的核心指标, 较高的地上生物量与高物种多样性、高稳定性是相伴而生的[32]。所以适当的放牧是合理的天然草地管理方式, 这也符合中度干扰能增加物种多样性的干扰理论。3)未发现物种多样性与地上生物量间特定的关系模型, 在以后的研究中应加入更加多元化的因子探究各指标对物种多样性差异的解释率。另外, 相对于其他类型草地(如高寒草甸、荒漠草原等), 黄土高原典型草原对氮素添加的响应是否更加缓慢或不敏感, 仍需进行长期研究, 可从物种个体功能性状响应等方面来进一步解释其内在机制。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|