


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
不同草地利用方式对暖性(灌)草丛类草地固碳能力的影响
[赵威*  , 李琳]
, 李琳]
, 李琳]
|
|


作者简介:赵威(1975-),男,内蒙古巴彦淖尔人,副教授,博士。E-mail:zhaowei1@haust.edu.cn
利用方式差异对草地生态系统碳循环影响不同,评价不同利用方式下各类型草地生态系统固碳能力有助于制定相应生态管理策略,对减缓草地温室气体排放具有现实意义。采用野外调查和室内试验相结合的方法,对河南不同利用方式下(围封未利用、零散放牧+割草、季节性放牧和全年放牧)两种典型草地(暖性草丛和暖性灌草丛)固碳能力进行研究。结果表明,暖性草丛植被生物量在不同利用方式上表现为围封未利用>零散放牧+割草>季节性放牧>全年放牧,但差异不显著( P>0.05);而暖性灌草丛植被生物量在零散放牧+割草条件下显著大于围封未利用和全年放牧方式( P<0.05)。在围封未利用条件下,暖性草丛植被生物量显著大于暖性灌草丛( P<0.05)。两种草地地上碳密度和根系碳密度在不同利用方式下差异均不显著( P>0.05)。从植被碳密度来看,暖性草丛在不同利用方式下表现为围封未利用>零散放牧+割草>全年放牧>季节性放牧,暖性灌草丛为零散放牧+割草>围封未利用>季节性放牧>全年放牧。季节性放牧条件下暖性草丛土壤碳密度(13369.07 g C·m-2)显著大于围封未利用方式(2544.25 g C·m-2)( P<0.05);暖性灌草丛土壤碳密度在不同利用方式下表现为零散放牧+割草>围封未利用>季节性放牧>全年放牧,但差异不显著( P>0.05)。不论在何种利用方式下,两种类型草地的植被碳密度均主要由根系所贡献,分别达到87.42%与81.52%;而生态系统碳密度均主要由土壤所贡献,分别达到91.72%与84.98%。双因素方差分析表明,土壤碳密度是决定河南两种类型草地固碳能力的根本因素,而与利用方式无关。本研究可为河南草地资源合理利用提供科学依据,为精确评估河南草地固碳能力提供数据支撑。
Differences in utilization patterns affect the carbon cycle of grassland ecosystems. Evaluating the carbon sequestration capacity of grassland ecosystems under different utilization patterns will help to establish relevant ecological management strategies, and so have practical significance for mitigating greenhouse gas emissions from grasslands. Based on a field survey and laboratory tests, the carbon sequestration capacity of two typical grassland types (WT: warm temperate tussock, and WS: warm temperate shrub-tussock) under four utilization patterns (FUG: fenced and unexploited grassland, SMG: scattered grazing+mown grassland, SGG: seasonally grazed grassland and AGG: all-year-grazed grassland) in Henan Province were studied. The results indicated that no difference was found in the vegetation biomass of WT ( P>0.05). However, the vegetation biomass of WS under SMG was significantly greater than that of FUG and AGG ( P<0.05). Under FUG, the vegetation biomass of WT was significantly greater than that of WS ( P<0.05). No significant difference existed in the underground and root carbon density between WT and WS ( P>0.05). With respect to the carbon density of vegetation (shoot+root), the management regimes ranked: FUG>SMG>AGG>SGG (2187.93-4211.56 g C·m-2) in WT. However, the soil carbon density of WT under SGG (13369.07 g C·m-2) was significantly greater than that of FUG (2544.25 g C·m-2) ( P<0.05). For WS, the root carbon storage was no significant difference among them ( P>0.05). For all utilization patterns, the root system was the major contributor to vegetation carbon density in both WT (87.42%) and WS (81.52%). With respect to the carbon density of the total grassland ecosystem, soil C (excluding roots), accounted for 91.72% and 84.98% of total C in WT and WS, respectively. Hence, soil carbon density was the major factor determining carbon sequestration capacity of both types of grassland, under all utilization patterns. This study provides scientific data to assist with rational allocation of grassland resources and for accurate assessment of carbon sequestration capacity of Henan grasslands.
人类活动干扰引发的自然生态系统碳释放是导致全球气候变化的重要因素, 受到国际社会的普遍关注[1, 2]。如何有效减缓草地生态系统CO2过量释放, 持续增强其固碳能力, 已经成为草地生态学研究的核心问题[3]。在陆地生态系统中, 草地生态系统既是碳源也是碳汇, 并在生物多样性维持、水土保持、水源涵养以及畜牧业发展中发挥着重要作用[4]。全球草地面积约为4.45× 106 hm2, 总碳储量可达7.61× 1011 t, 是陆地生态系统碳库的重要组成部分[5]。我国约有天然草地面积4.06× 108 hm2, 占整个国土面积的41%[6], 草地植被碳储量约占陆地总植被碳储量的16.7%[7]。许多学者对我国草地生态系统碳库总量进行了估算[8, 9, 10], 但由于采用方法不同使得结果存在较大差异, 所得数值介于0.56~3.32 Pg C之间, 相差近6倍。其原因主要是在对各类型草地进行碳估算时, 不能精确区分区域土地利用方式差异对估算结果所产生的重大影响。
土地利用变化在全球碳平衡及气候变化中起着重要作用, Foley等[11]的研究表明, 人类活动排放的CO2中约35%直接来源于土地利用方式。不同的利用方式会改变植被组成, 同时也是影响土壤碳收支的重要因素, 进而影响整个陆地生态系统的碳源与碳汇功能[12]。强烈的干扰则会导致众多自然因素的变化, 从而改变生态过程, 影响整个生态系统碳固定进程[13]。作为人类各种土地利用活动的综合反映, 土壤变化是影响土地状况最直接与最深刻的因素[14, 15, 16]。土壤有机碳密度水平不仅影响植被生产潜力的高低, 其动态变化和区域差异也会影响土壤有机碳与大气CO2交换的生态环境效应[17]。通常状况下, 农业利用会降低土壤有机碳密度, 对温室气体排放的贡献率可达20%[18]。近年来, 许多学者对我国各类生态系统土壤碳密度的区域差异进行研究[19, 20], 但针对不同利用方式下河南草地土壤有机碳密度变化的鲜有报道。
由于天然牧草与大田作物对气候、土壤等自然因子要求相似, 我国草地畜牧业与种植业呈相互交错态势。作为最广泛的利用方式之一, 放牧对草地生态系统碳储量有着深刻的影响, 过度放牧导致草地退化与水土流失, 是造成草地碳储量变化的重要原因。在2000年之前, 我国对天然草地缺少监管, 大多数草地采取粗放的自然放牧方式, 因此大面积草地出现了不同程度的退化、沙化[21, 22, 23]。Wang 等[24]研究表明, 中国北方的中、重度退化草地共造成了近1.24 Pg C的净碳损失。围封禁牧是我国恢复退化草地的主要措施之一, 研究表明, 在重牧区实施围栏封育措施, 草地的固碳潜力每年可增至12.01 Tg C[25]。但近年来有关围封禁牧与放牧对草地固碳能力的影响也存在争议, 主要是由于处于不同状态、不同类型和不同放牧历史的草地, 其固碳能力对禁牧与放牧的响应机制呈现多样化态势[26]。
河南气候温和, 雨量充沛, 拥有各类草地面积约4.60× 106 hm2, 占全省土地面积的27.54%, 占全国草地面积的1.20%, 但由于河南农业历史悠久, 人口众多, 平原地区草地早已开垦, 现存草地多集中在西北、南部浅山丘陵区, 且分布零散[27], 导致草地资源开发利用不平衡, 畜牧业生产水平较低。本研究在河南选择具有代表性的39个面上采样点进行调查取样, 对不同利用方式下草地有机碳密度及其分布进行研究, 并比较两种典型草地类型(暖性草丛、暖性灌草丛)的差异, 以期为河南草地资源合理利用提供科学依据, 为精确评估河南草地固碳能力提供数据支撑。
河南省是我国中原腹心地带, 地跨31° 23'-36° 22' N, 110° 21'-116° 39' E, 位于黄河中下游, 呈西高东低地势, 北、西、南三面被太行山脉、伏牛山脉、桐柏山脉、大别山脉环绕, 中、东部为黄淮海平原, 西南部为南阳盆地。河南属暖温带、湿润至半湿润季风气候, 四季分明。年平均气温12~16 ℃, 无霜期180~240 d, 全年日照时数2000~2600 h, 年平均降水量500~900 mm, 多集中在夏季。河南省大部分草地分布在豫北、豫南、豫西等地, 地形多丘陵、低山、中山、高山, 土壤类型主要为褐土、钙质石质土、棕壤性土、黄棕壤和黄绵土等。植物群落主要由优势种黄背草(Themeda japonica)、白羊草(Bothriochloa ischaemum)、铁杆蒿(Artemisia gmelinii)、白茅(Imperata cylindrica)、多花胡枝子(Lespedeza floribunda)等与其他杂类草构成, 具有耐寒、耐旱的生态特征。
1.2.1 样地选择 在20世纪90年代全国草地类型图和2000年土地利用图基础上, 结合1997-1998年郭孝等[27, 28]对河南草地的调查结果, 于2012年进行了不同利用方式下河南草地植被与土壤固碳状况调查研究。主要选取暖性草丛和暖性灌草丛两种区域典型草地, 同时通过大量走访调查, 根据河南草地利用现实情况, 最终确定4种主要利用方式。其中, 围封未利用草地为退耕(还林)还草工程确定的围封区域, 限制放牧与割草; 零散放牧+割草草地为拥有10~20只山羊或2~3头牛的农户, 利用农闲时间零散放牧或者农忙时割草饲喂家畜的草地; 季节性放牧草地为拥有20~30只山羊或5~8只牛的专业养殖户, 利用夏秋季节牧草长势最好时在附近草坡进行放牧的草地, 春冬季则进行棚圈饲喂; 全年放牧草地为拥有30~50只山羊或8~10只牛的专业养殖户, 在所承包的丘陵坡地上进行全年放牧的草地。
1.2.2 样地分布及取样原则 本研究于2014-2015年间每年的7-9月, 在草地群落生物量累积最大时期取样。根据两种草地的典型性和代表性共设置了39个面上采样点, 分别位于河南省安阳、三门峡、洛阳、南阳、信阳等8个地市, 其中围封未利用样地9个, 零散放牧+割草样地11个, 季节性放牧样地9个, 全年放牧样地10个(图1)。在设置的39个样点中, 按照坡度方向设置一条100 m的样线, 每隔10 m设定一个1 m× 1 m的草本植物样方, 包括5个草本分种样方和5个草本不分种样方, 分种样方确定不同利用方式下两种类型草地的优势种及其群落特征, 不分种样方测定两种类型草地的地上与地下生物量, 以及植被与土壤碳含量。此外, 样线每隔20 m设定一个5 m× 5 m的灌木样方, 调查灌木种类与群落特征。
群落盖度采用目测法估计样方内各物种冠层的投影面积占样方面积的比例; 物种多样性选用Magalef丰富度指数(D)、Pielou均匀度指数(J)和Shannon-Wiener指数(H)测定, 计算公式如下:
式中:Pi为种i的相对重要值, 即(相对盖度+相对高度+相对频度)/3; S为种i在样方内的株丛数; N为个体总数。
土壤容重的测定采用环刀法。每个样线中心挖一个长、宽、深分别为150 cm× 50 cm× 100 cm的取样坑, 用5 cm环刀按照0~5 cm、5~10 cm、10~20 cm、20~30 cm、30~50 cm、50~70 cm、70~100 cm的不同深度从上至下取样, 每层取5个重复。装袋后带回实验室105 ℃烘干至恒重, 然后称量其干重。土壤有机碳含量采用重铬酸钾外加热法测定[2], 全氮含量采用Carlo-Erba的NA1500元素分析仪进行测定。土壤容重=土壤烘干重/环刀体积。碳氮比=土壤全碳/全氮。草地群落特征及土壤理化性质见表1。
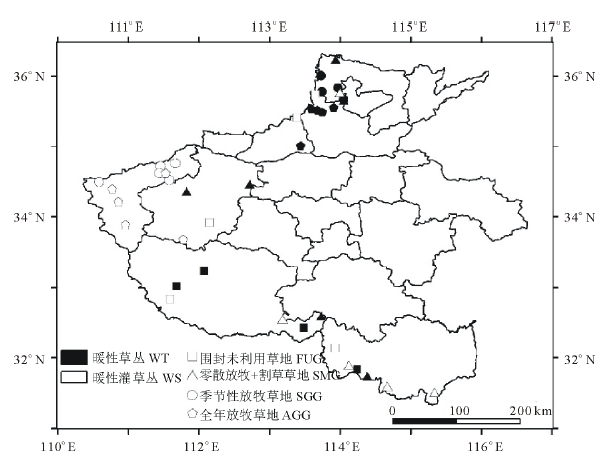 | 图1 取样点分布Fig.1 Distribution of sampled plots WT: 暖性草丛Warm-temperate tussock; WS: 暖性灌草丛Warm-temperate shrub tussock; FUG: 围封未利用草地Fenced and unexploited grassland; SMG: 零散放牧+割草草地Scattered grazing+mowing grassland; SGG: 季节性放牧草地Seasonal grazing grassland; AGG: 全年放牧草地All-year grazing grassland. 下同The same below. |
| 表1 不同利用方式下两种类型草地基本群落特征与土壤理化性质 Table 1 Basic community and physical-chemical characteristics of two types of grassland under different utilization patterns |
在不分种样方内采用收获法测定群落地上生物量, 地上活体生物量采用齐地刈割法, 将样方内植物地面以上的绿色部分用剪刀齐地剪下, 装袋保存, 在65 ℃下48 h烘干至恒重, 称量, 得到活体生物量。凋落物生物量是指地面以上还没有发生明显分解的多年积累的植物死亡残体, 半分解层是指枯枝落叶等中度分解的有机层, 用小耙子收集地上的凋落物和半分解层, 除去土粒和杂质, 带回实验室烘干至恒重, 称量。地上生物量为活体生物量与凋落物生物量之和, 植被生物量为地上生物量与根系生物量之和。
地下生物量的测定采用根钻法分层取样。根钻直径为7.5 cm, 根系取样分为0~5 cm、5~10 cm、10~20 cm、20~30 cm、30~50 cm、50~70 cm、70~100 cm 7个土层, 每层取3钻后合并为一个样品, 在0.125 mm筛中清洗干净后装入网袋风干, 在称重前再放进烘箱65 ℃烘至恒量。土壤样品采集方法与根系生物量相同, 分层装好后, 带回实验室置于室内阴凉处风干, 去除杂质, 研磨后过0.25 mm筛备用。
将烘干的植物样品粉碎研磨后, 用Carlo-Erba的NA1500元素分析仪测定地上植物样品的总碳含量。根据公式计算植物地上活体、凋落物、半分解层和根系的碳密度。地上碳密度为活体、凋落物与半分解层碳密度之和, 植被碳密度为地上碳密度与根系碳密度之和, 生态系统碳密度为植被碳密度与土壤有机碳密度之和。
Cd=α × M
式中:Cd为碳密度(g C· m-2); α 为生物量有机碳含量(%); M为地上或地下生物量(g· m-2)。
根据以下公式计算土壤有机碳密度(SOCdensity, g C· m-2):
SOCdensity=
式中:n为土层数; Di为不同土层厚度(cm); θ i为土壤容重(g· cm-3); Ci为不同土层土壤有机碳含量(%); δ i为> 2 mm砾石含量(体积%)。
利用Microsoft Excel进行数据整理与表格绘制。利用SPSS 18.0软件one-way ANOVA模块进行单因素方差分析、two-way ANOVA模块进行多因素方差分析以及Duncan多重比较。利用OriginPro 9.0软件作图。
如表 2 所示, 不同利用方式对暖性草丛的活体生物量、凋落物生物量以及地上生物量均无显著影响(P> 0.05), 在同一利用方式下, 暖性草丛的活体生物量、凋落物生物量以及地上生物量也无显著差异(P> 0.05); 与暖性草丛类似, 暖性灌草丛呈现相同的规律。同一利用方式下, 暖性草丛与暖性灌草丛地上生物量也没有显著差异(P> 0.05), 但暖性草丛在零散放牧+割草条件下地上生物量最高, 为292.43 g· m-2, 而暖性灌草丛在季节性放牧条件下地上生物量最高, 为356.81 g· m-2。不同利用方式对暖性草丛的根系生物量没有显著影响(P> 0.05), 但对暖性灌草丛根系生物量有一定影响。在零散放牧+割草条件下, 暖性灌草丛的根系生物量显著大于围封未利用与全年放牧条件(P< 0.05)。在围封未利用条件下, 暖性草丛的根系生物量显著大于暖性灌草丛(P< 0.05), 但其他3种利用方式下, 两种类型草地之间并无显著差异(P> 0.05)。不同利用方式对暖性草丛的植被生物量没有显著影响(P> 0.05), 但对暖性灌草丛植被生物量有一定影响。在零散放牧+割草条件下, 暖性灌草丛的植被生物量显著大于围封未利用与全年放牧条件(P< 0.05)。在围封未利用条件下, 暖性草丛的植被生物量显著大于暖性灌草丛(P< 0.05), 但其他3种利用方式下, 两种类型草地之间并无显著差异(P> 0.05)。不论在何种利用方式下, 两种类型草地的根系生物量都是植被生物量的主体, 占整个植被生物量的60%以上。
| 表2 不同利用方式下两种类型草地植被生物量构成 Table 2 Vegetation biomass composition of two types of grassland under different utilization patterns (g· m-2) |
不同利用方式下两种类型草地活体和凋落物碳密度差异如图2所示。4种利用方式下暖性草丛活体平均碳密度为64.02 g C· m-2, 大小顺序为:季节性放牧> 全年放牧> 零散放牧+割草> 围封未利用, 其中季节性放牧草地活体碳密度最大, 为80.04 g C· m-2, 但与其他3种利用方式相比差异并不显著(P> 0.05)。4种利用方式下暖性灌草丛活体平均碳密度为68.14 g C· m-2, 大小顺序为:零散放牧+割草> 围封未利用> 季节性放牧> 全年放牧, 其中零散放牧+割草草地活体碳密度最大为83.72 g C· m-2, 但与其他3种利用方式相比差异同样不显著(P> 0.05)。相同利用方式下两种类型草地活体碳密度均无显著差异(P> 0.05)。
4种利用方式下暖性草丛凋落物平均碳密度为21.75 g C· m-2, 大小顺序为:围封未利用> 零散放牧+割草> 季节性放牧> 全年放牧, 其中围封未利用草地凋落物碳密度最大为37.74 g C· m-2, 但与其他3种利用方式相比差异不显著(P> 0.05)。4种利用方式下暖性灌草丛凋落物平均碳密度为34.02 g C· m-2, 大小顺序为:季节性放牧> 围封未利用> 零散放牧+割草> 全年放牧, 其中季节性放牧草地凋落物碳密度最大为51.80 g C· m-2, 但与其他3种利用方式相比差异并不显著(P> 0.05)。相同利用方式下2种类型草地凋落物碳密度无显著差异(P> 0.05)。
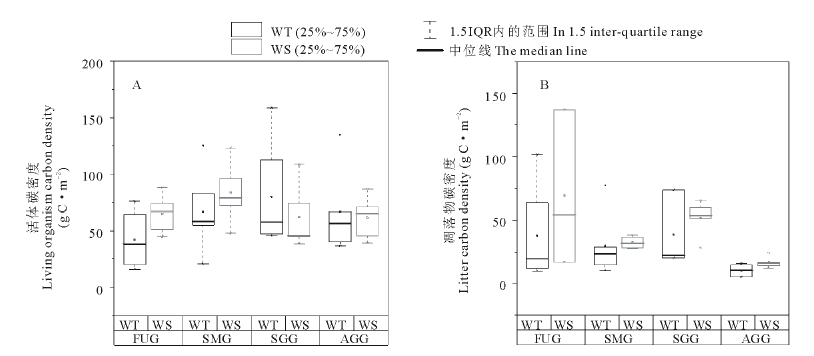 | 图2 不同利用方式下两种类型草地活体和凋落物碳密度变化Fig.2 Changes of living organism and litter carbon density of two types of grassland under different utilization patterns * 表示异常值, □表示平均值。“ * ” indicated outlier, “ □” indicated mean. 下同The same below. |
不同利用方式下两种类型草地地上和根系碳密度如图3所示。4种利用方式下暖性草丛地上平均碳密度为89.48 g C· m-2, 大小顺序为:季节性放牧> 零散放牧+割草> 围封未利用> 全年放牧, 其中季节性放牧草地地上碳密度最大, 为109.01 g C· m-2, 但与其他3种利用方式相比差异并不显著(P> 0.05)。4种利用方式下暖性灌草丛地上平均碳密度为102.17 g C· m-2, 大小顺序为:季节性放牧> 零散放牧+割草> 围封未利用> 全年放牧, 与暖性草丛趋势一样, 其中季节性放牧草地地上碳密度最大, 为114.03 g C· m-2, 但与其他3种利用方式相比差异并不显著(P> 0.05)。相同利用方式下2种草地类型地上碳密度均无显著差异(P> 0.05)。
4种利用方式下暖性草丛根系平均碳密度为595.99 g C· m-2, 大小顺序为:围封未利用> 零散放牧+割草> 全年放牧> 季节性放牧, 其中围封未利用草地根系碳密度最大, 为797.61 g C· m-2, 但与其他3种利用方式相比差异并不显著(P> 0.05); 4种利用方式下暖性灌草丛根系平均碳密度为450.64 g C· m-2, 低于暖性草丛根系平均碳密度, 大小顺序为:零散放牧+割草> 围封未利用> 季节性放牧> 全年放牧, 其中零散放牧+割草草地根系碳密度最大, 为543.65 g C· m-2, 全年放牧草地根系碳密度最小, 为229.17 g C· m-2, 二者之间差异并不显著(P> 0.05)。全年放牧条件下暖性草丛根系碳密度显著大于暖性灌草丛(P< 0.05), 但其他3种利用方式下两种类型草地差异并不显著(P> 0.05)。
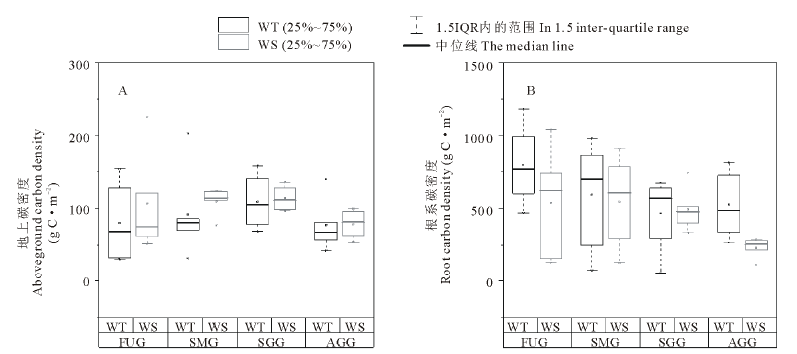 | 图3 不同利用方式下两种类型草地地上和根系碳密度变化Fig.3 Changes of aboveground and root carbon density of two types of grassland under different utilization patterns |
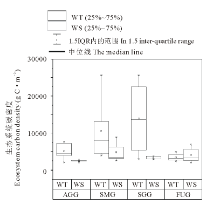
不同利用方式下两种类型草地植被和土壤碳密度如图4所示。4种利用方式下暖性草丛植被平均碳密度为681.76 g C· m-2, 大小顺序为:围封未利用> 零散放牧+割草> 全年放牧> 季节性放牧, 其中围封未利用草地植被碳密度最大, 为877.58 g C· m-2, 但与其他3种利用方式相比差异并不显著(P> 0.05)。4种利用方式下暖性灌草丛植被平均碳密度为552.80 g C· m-2, 大小顺序为:零散放牧+割草> 围封未利用> 季节性放牧> 全年放牧, 其中零散放牧+割草草地植被碳密度最大, 为653.32 g C· m-2, 全年放牧最小为307.43 g C· m-2, 但差异并不显著(P> 0.05)。全年放牧条件下暖性草丛植被碳密度显著大于暖性灌草丛(P< 0.05), 但其他3种利用方式下植被碳密度均无显著差异(P> 0.05)。
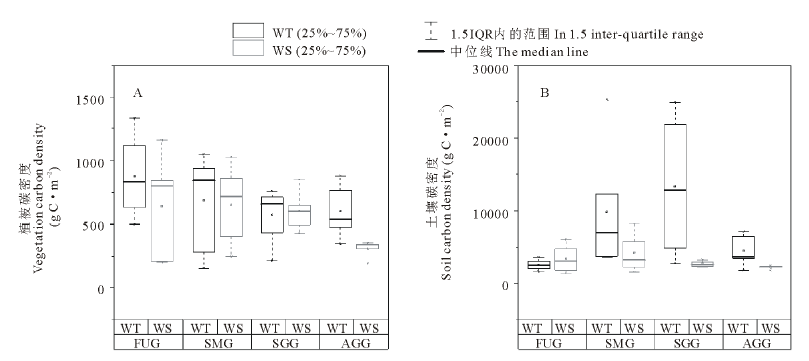 | 图4 不同利用方式下两种类型草地植被和土壤碳密度变化Fig.4 Changes of vegetation and soil carbon density of two types of grassland under different utilization patterns |
4种利用方式下暖性草丛土壤平均碳密度为7556.89 g C· m-2, 大小顺序为:季节性放牧> 零散放牧+割草> 全年放牧> 围封未利用, 其中零散放牧+割草草地土壤碳密度最大, 为13369.07 g C· m-2, 是平均值的1.77倍, 显著大于其他3种利用方式(P< 0.05)。4种利用方式下暖性灌草丛土壤平均碳密度为3128.67 g C· m-2, 低于暖性草丛土壤平均碳密度, 大小顺序为:零散放牧+割草> 围封未利用> 季节性放牧> 全年放牧, 其中零散放牧+割草草地土壤碳密度最大, 为4211.56 g C· m-2, 但与其他3种利用方式相比差异不显著(P> 0.05)。相同利用方式下2种类型草地土壤碳密度均无显著差异(P> 0.05)。
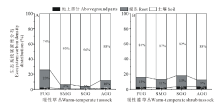
不同利用方式下两种类型草地生态系统碳密度及其各组分所占比例如图5与图6所示。草地生态系统有机碳含量约74.35%~95.97%由土壤贡献, 其次是根系贡献约3.34%~23.31%, 地上部分仅贡献0.78%~3.45%。4种利用方式下暖性草丛生态系统平均碳密度为8238.65 g C· m-2, 大小顺序为:季节性放牧> 零散放牧+割草> 全年放牧> 围封未利用, 其中季节性放牧草地生态系统碳密度最大, 为13929.86 g C· m-2, 显著高于围封未利用草地生态系统碳密度(3421.82 g C· m-2)(P< 0.05)。4种利用方式下暖性灌草丛生态系统平均碳密度为3681.47 g C· m-2, 大小顺序为:零散放牧+割草> 围封未利用> 季节性放牧> 全年放牧, 其中零散放牧+割草草地生态系统碳密度最大, 为4864.87 g C· m-2, 显著大于其他3种利用方式(P< 0.05)。全年放牧条件下暖性草丛生态系统碳密度显著大于暖性灌草丛(P< 0.05), 但其他3种利用方式下二者之间均无显著差异(P> 0.05)。
 | 图5 不同利用方式下两种类型草地生态系统碳密度变化Fig.5 Changes of ecosystem carbon density of two types of grassland under different utilization patterns |
 | 图6 不同利用方式下两种类型草地生态系统碳密度分布变化Fig.6 Changes of ecosystem carbon density distribution of two types of grassland under different utilization patterns |
对不同利用方式下两种类型草地各碳库碳密度进行单变量双因素方差的结果表明(表3), 草地类型和利用方式两个主因子对活体、凋落物、地上部分、根系、植被碳密度均无显著影响(P> 0.05), “ 草地类型× 利用方式” 的互作效应也不显著(P> 0.05)。但草地类型这一因子对土壤和生态系统碳密度有极显著影响(P< 0.01)。由此结果可知, 河南两种类型草地的固碳能力与草地类型本身有关, 而与利用方式和二者的互作效应无关, 主要体现在对土壤碳密度的影响最为显著, 从而决定了生态系统的固碳能力。
| 表3 草地类型与利用方式的双因素方差分析 Table 3 Two-factor analysis of variances between grassland types and utilization patterns |
草地生产力是反应草地生态系统健康的重要因素, 也是体现群落功能的重要载体。而草地群落地上现存生物量是反映草地物质积累状况和生产潜力的重要指标[29], 其形成过程与草地植物光合产物累积方式与受干扰程度密切相关[30]。草地利用方式差异对群落生产力产生重要影响[31], 而草地植被碳密度在很大程度上依赖于草地群落生物量的积累。围栏封育有利于实现退化草地系统的自我恢复[2, 32], 但围封时间过长不利于提高草地的生产力, 草地长期未利用会导致群落地上生物量逐渐降低、凋落物增加, 抑制了植物的再生和幼苗的建成, 从而阻碍了草地植被的更新[33, 34]。通过加速营养循环、改善冠层辐射状况, 适当放牧可以提高植物的光合能力, 优化资源分配, 促进草地植物进行补偿性生长[35], 从而增加草地地上生物量。根系是植物吸收和贮藏营养物质的重要器官, 根系的数量及其分布直接影响到草地植物吸收和储存营养物质的能力, 同时也可反映出植物的生存环境状况[2]。本研究结果显示, 不同利用方式下两种类型草地的地上生物量差异尽管不显著(P> 0.05), 但均随着利用方式的不同而变化, 尤其在零散放牧+割草和季节性放牧条件下达到最大值, 而全年放牧草地地上生物量最低。Zhou等[36]研究发现, 尽管围封增加了高寒灌丛草甸的地上植被生物量, 但在轻度放牧时地上生物量达到最高; 王艳芬等[29]也发现, 轻牧至中牧条件下植物生物量较围封未利用有所提高, 但随着放牧强度的增加, 地上生物量又随之降低。4种利用方式下草地的地下根系生物量是植被生物量的主体, 约占整个植被生物量的62%以上, 主要是由于本研究实地调查地下生物量时, 将活根和死根都考虑在内; 而且河南草地灌木分布广泛, 地下根系交错盘旋, 根系取样时不可避免取到灌木根系, 因而导致根系生物量在植被生物量中占据较大比例。放牧是一种复杂的人为干扰方式, 对草地群落既存在积极作用也有消极影响[37]。与围封未利用草地相比, 适当的放牧强度可以消除植物的生长冗余, 增加植物净初级生产潜力[38, 39]。随着利用强度的增加, 暖性草丛根系生物量随之降低, 而暖性灌草丛先增加后降低, 表明暖性草丛与暖性灌草丛在应对外界干扰时具有不同的响应机制, 暖性灌草丛植被地上部分的损失改变了生物量分配格局, 光合物质向地下根系转移, 从而使根系生物量显著增加[40]。
不同的利用方式会改变植被组成, 同时也是影响土壤碳收支的重要因素, 进而影响整个生态系统的碳源与碳汇功能[12]。本研究表明, 河南草地暖性草丛与暖性灌草丛的地上平均碳密度分别为89.48和102.17 g C· m-2, 根系平均碳密度分别为595.96和450.64 g C· m-2。Fan等[41]根据草地资源清查数据, 估算出北方草地地上碳密度为77.3 g C· m-2, 根系碳密度为1.21× 103 g C· m-2; 中国草地地上碳密度为119.00 g C· m-2, 根系碳密度为883.00 g C· m-2, 本研究结果与此较为接近, 但和我国第一次草地资源清查资料和遥感数据估算值[42, 43]存在较大差异, 其原因主要是草地分类系统和资料来源不同, 以及估算方法不同。本研究采用统一的草地生态系统调查规范, 草地生物量与碳密度均为实测数据, 因此准确性较高。在全年放牧条件下, 暖性草丛根系碳密度与植被碳密度均显著大于暖性灌草丛(P< 0.05), 主要原因是:1)暖性灌草丛虽然灌木丰富度高于暖性草丛, 但暖性草丛并不是没有灌木, 而是多分布有多花胡枝子、中华绣线菊(Spiraea chinensis)、卵叶鼠李(Rhamnus bungeana)等小型灌木和少量酸枣(Ziziphus jujube)、黄荆(Vitex negundo)幼苗, 在取样中高度小于0.5 m的灌木均按草本处理, 一定程度上增加暖性草丛的根系生物量; 2)暖性草丛中多分布有高大丛生的C4植物黄背草, 其根系非常发达, 对暖性草丛根系碳密度贡献非常明显。
开垦和放牧是草地最主要的利用方式[44], 强烈影响着草地土壤有机碳循环和土壤理化性质[45]。土壤容重作为判断土壤肥力状况的重要指标, 同时可是评价放牧草地退化状况的必备参考因素[46, 47]。本研究中土壤容重随着利用方式的变化而变化, 大小顺序为:全年放牧> 季节性放牧> 零散放牧+割草> 围封未利用, 其中全年放牧最大, 达1.84(暖性草丛)和1.67 g· cm-3(暖性灌草丛), 其原因主要是由于放牧家畜的踩踏导致土壤紧实度增加[48, 49], 表明草地放牧利用对两种草地的土壤产生了较大影响。此外, 河南草地土壤中砾石偏多, 这也是造成土壤容重过大的重要原因。土壤有机碳是草地土壤质量评价的重要指标, 与土壤的物理、化学和生物性质有紧密的联系[50]; 而氮素是植物生长发育所必需的大量营养元素, 是土壤养分的重要指标, 也是草地生态系统生产力限制因素[51]。草地利用导致河南草地土壤碳氮比呈下降趋势, 表明人为干扰一定程度影响了河南草地土壤碳固定, 这与已有的研究结果一致[52, 53, 54]。
河南两种类型草地生态系统有机碳密度的74.35%~95.97%由土壤所贡献, 土壤碳密度决定着此类草地生态系统碳密度的变化。不同利用方式下暖性草丛土壤平均碳密度为7556.89 g C· m-2, 生态系统平均碳密度为8238.65 g C· m-2; 暖性灌草丛土壤平均碳密度为3128.67 g C· m-2, 生态系统平均碳密度为3681.47 g C· m-2。随着利用方式的变化两种类型草地土壤碳密度也随之变化, 分别在季节性放牧和零散放牧+割草草地达到最大。这与中度干扰假说相符合, 表明适度利用可以通过改变土壤理化性质, 增加生境的异质性, 从而造成草地群落植物组分结构和多样性格局发生变化, 进而对整个生态系统的结构和功能产生影响[55]。有研究表明, 禁牧可以减少草地地上植被的损失, 从而降低碳素的损失, 利用土壤碳储量积累, 在干旱与半干旱地区退化草地实行围栏禁牧后, 土壤碳储量高于放牧条件下土壤碳储量[56]。但也有研究证实禁牧对草地碳储量存在不利影响, 放牧在一定条件下可以增加土壤碳储量[57], 而暖性草丛草地全年放牧土壤碳密度大于围封未利用草地土壤碳密度, 可能与植被地上凋落物有关, 围封未利用草地地上凋落物的积累降低了植被光合效率, 从而降低碳从植被向土壤转移的速率, 对土壤碳储量的积累有负面作用[58, 59]。草地生态系统的土壤碳储量受植被生产力和土壤碳循环过程变化的影响, 植被地上生产力的增加会增加碳素在土壤中的积累, 在一些受放牧活动影响的、植被地上生产力下降的草地会出现土壤碳储量增加的现象, 这可能与植被地下生产力的增加或地下与地上生产力分配比例变化有关[26]。植被的群落结构会影响草地的生产力, 从而间接影响土壤碳储量, Schuman等[59]研究发现, 放牧活动会增加根系发达的格兰马草(Bouteloua gracilis)在群落中的比例, 改变了地上与地下生物量的分配, 从而影响土壤碳储量; 而草地中的豆科(Leguminosae)植物也会增加土壤的碳储量。植被生物量的增加, 植被呼吸作用也会增加, 对于一些物种来说, 光合作用固碳量与呼吸作用碳损失量之间的比值会下降, 会降低碳的固定和转移。放牧会减少地上植被的生物量, 但并不一定降低植被光合作用固碳量, 较高的光合效率和较低的呼吸效率反而会增加土壤碳的固定[60]。放牧对草地土壤碳储量的影响机制是复杂的, 目前的研究仍存在较多争议[26]。草地碳储量究竟在多大程度受利用方式影响, 通常认为依赖于不同草地类型土壤碳固定机制对放牧响应的差异[61]。本研究通过双因素方差分析得出, 利用方式并不是影响河南两种类型草地土壤碳储量变化的主要因素, 草地类型与利用方式的交互效应也不显著(P> 0.05), 而与草地类型本身极显著相关(P< 0.01), 表明河南草地土壤可能存在自身多样化的固碳机制, 这需要在今后的研究中进一步加以证实。
不同利用条件下暖性草丛的植被生物量无显著差异(P> 0.05), 在零散放牧+割草条件下暖性灌草丛植被生物量显著大于围封未利用和全年放牧方式(P< 0.05), 表明适度利用有利于提高暖性灌草丛草地生产力。
围封未利用条件下暖性草丛植被碳密度大于其他利用方式, 但差异不显著(P> 0.05)。暖性灌草丛在零散放牧+割草条件下植被碳密度最大, 在全年放牧条件下最低。表明利用强度的增加降低了草地植被生产力, 从而影响植被碳密度。季节性放牧条件下, 暖性草丛土壤碳密度显著大于围封未利用方式(P< 0.05); 暖性灌草丛土壤碳密度在不同利用方式下表现为零散放牧+割草> 围封未利用> 季节性放牧> 全年放牧, 但差异不显著(P> 0.05)。表明利用方式的改变会影响草地地上与地下的碳分配, 从而影响土壤碳储量。
暖性草丛生态系统碳密度在季节性放牧条件下显著高于围封未利用方式(P< 0.05), 而暖性灌草丛在零散放牧+割草(4864.88 g C· m-2)条件下显著高于全年放牧(P< 0.05)。暖性草丛生态系统平均碳密度高于暖性灌草丛, 表明河南不同类型草地对利用方式的响应不同, 进而反映出草地生态系统碳库的变化。双因素方差分析结果表明, 利用方式不是影响河南两种类型草地土壤碳库变化的主导因素, 而与草地类型本身有关, 表明河南草地土壤固碳存在自身的特有机制。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|