

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
外源复合激素对匍匐翦股颖抗褐斑病的诱导抗性
[赵春旭1*, *  , 姜寒玉
, 姜寒玉2*, * , 董文科1 , 陈红2 , 方彦霞2 , 谢丽萍2 , 马晖玲1, * ]
, 姜寒玉, 董文科|
|


作者简介:赵春旭(1985-),男,甘肃兰州人,在读博士。E-mail: zhaocx@gsau.edu.cn;姜寒玉(1980-),女,甘肃定西人,副教授,博士。E-mail:jianghy@gsau.edu.cn。**共同第一作者These authors contributed equally to this work.
以匍匐翦股颖品种“Penn-A4”为材料,研究以不同浓度的油菜素内酯(EBR)、水杨酸(SA)和乙烯(ET)组合的外源复合激素和3个连续诱导时间(5、7、10 d)为变量,通过测定病情指数、防治效果、抗氧化酶活性及抗坏血酸-谷胱甘肽循环变化,分析外源复合激素及连续诱导时间对匍匐翦股颖抗褐斑病的诱导效应。结果表明,不同激素组合和不同连续诱导时间处理均可提高匍匐翦股颖的抗病性,其中ET+EBR组合连续诱导处理7 d的病情指数最低,为13.36,显著低于其他处理,此时防治效果为40.51%;另外,此组合诱导处理后,匍匐翦股颖幼苗叶片的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)分别比对照(CK)提高了130.53%、197.40%和101.66%;抗坏血酸过氧化物酶(APX)、脱氢抗坏血酸还原酶(DHAR)、谷胱甘肽还原酶(GR)活性分别比对照(CK)提高了316.26%、41.95%和80.34%;脱氧抗坏血酸(AsA)、氧化型谷胱甘肽(GSH)含量以及脱氧抗坏血酸/氧化型抗坏血酸(AsA/DHA)、氧化型谷胱甘肽/还原型谷胱甘肽(GSH/GSSG)也均高于对照(CK)和其他处理。表明EBR、SA和ET三种物质组成的复合外源物可以诱导匍匐翦股颖抗褐斑病,其中ET+EBR组合连续诱导处理7 d的效果最好。该研究结果为新型复合诱导抗病剂的应用推广提供了技术支持。
, JIANG Han-yu, DONG Wen-keComposite plant hormone formulations with different concentrations of epibrassinolide (EBR), salicylic acid (SA) and ethephon (ET) were applied exogenously to “Penn-A4” bentgrass with induction times of 5, 7, and 10 days to test if the hormone treatments could enhance disease resistance. The effect of the exogenous composite hormone application and induction time treatments on the brown spot resistance of creeping bentgrass was assessed by determining a disease index, the control efficacy, antioxidant enzyme activity and ascorbic acid/glutathione cycle changes. Some treatments did improve the disease resistance of creeping bentgrass. The disease index for an ET+EBR hormone treatment with 7 days induction was significantly lower than other treatments, and the anti-disease effect was quantified at 40.51%. Moreover, superoxide dismutase (SOD), catalase (CAT) and peroxidase (POD) activities increased by 130.53%, 197.40% and 101.66%, respectively, while ascorbate peroxidase (APX), dehydroascorbate reduction (DHAR), and glutathione reductase (GR) activities increased by 316.26%, 41.95% and 80.34%, respectively, compared with the control, after this treatment. In addition, active deoxyascorbic acid (AsA), oxidized glutathione (GSH) content, and AsA/hydroascorbic acid (DHA), and GSH/reduced glutathione (GSSG) ratios were all higher for the ET+EBR combination, than for the control and other treatments. In summary, exogenous application of mixtures of EBR, SA and ET were able to induce brown spot resistance in creeping bentgrass, and an ET+EBR combination with an induction period of 7 days had the best effect. The results of this study provide technical information for the development and promotion of novel hormone-based agents for induction of plant disease resistance.
匍匐翦股颖(Agrostis stolonifera)是禾本科翦股颖属多年生草本植物, 因其草质细腻, 耐频繁低度修剪, 常被用于高尔夫球场果岭、草地网球场、草地保龄球场等精细草坪[1]。褐斑病是草坪病害中分布最广的病害之一, 是北方冷季型高尔夫球场果岭上发生最普遍、最严重的病害之一[2]。1914年褐斑病首次在草坪草上被发现, 后由Piper等[3]首次报道其病原菌为立枯丝核菌(Rhizoctonia solani), 之后又被众多研究者证实[4, 5, 6]。立枯丝核菌是一种土壤习居菌, 传播迅速, 寄主范围广, 最适生长温度为30 ℃, 一般在温暖潮湿的区域发病率较高, 并且该菌几乎可以侵染所有草坪草, 严重影响了草坪的质量、美观度以及绿化功能, 给草坪生产和经营带来了巨大损失。
目前, 草坪病害的防治主要依赖化学杀菌剂[7, 8], 但化学防治成本较高, 且严重污染环境, 同时病原物易对杀菌剂产生耐药性, 防治效果逐渐减弱。诱导植物抗病性是指利用物理、化学以及生物的方法, 激发植物自身对病原微生物的抗病性的现象[9, 10], 它具有环保且抗病周期长的优点, 已成为植物病害防治的研究热点, 许多化学因子可诱导植物产生抗病性。水杨酸(salicylic acid, SA)是植物体内一种简单的酚类物质, 是一些重要代谢过程的信号分子, 参与植株系统抗性的建立, 可以显著提高植株的抗病性; 研究表明, 外源SA诱导处理植株后, 体内多酚氧化酶和苯丙氨酸解氨酶活性显著上升, 诱导植株产生抗性[11]。油菜素内酯(epibrassinolide, EBR)是一种天然植物激素, 它已经被公认为第六类植物激素, 对植物的生长和发育、生物和非生物胁迫的发展起着至关重要的作用, 研究表明EBR可诱导多种植物对真菌、细菌及病毒产生抗性[12]。乙烯(ethylene, ET)是五大类植物激素之一, 参与了植物从种子萌发到成熟以及衰老的一系列生命过程的调节及生物和非生物胁迫下植物的抗性反应[13], 研究表明当植物受到病原物侵染时, 乙烯量急剧增加[14], 是病原菌侵染反应的报警信号并参与防御反应。尽管国内外针对植物诱导抗病性也已经展开了一系列的研究, 但目前国内外使用的植物抗病诱导剂仍存在诱导效果不佳, 效果不稳定, 诱导持续时间不长等缺点。为此, 本研究选择3种化学激素诱导剂, 即油菜素内酯(EBR)、水杨酸(SA)及乙烯(ET), 根据不同外源激素活性特点及作用原理, 研究复合激素以及连续诱导时间的协同增效功能, 旨在明确不同激素之间的协同效应, 筛选出诱导效果更好, 抗病持续时间更长的新型复合诱导剂。
供试材料为匍匐翦股颖“ Penn-A4” (A. stolonifera cv.“ Penn-A4” ), 由北京克劳沃草业技术开发中心提供; 诱导剂EBR、SA及ET均购自Sigma公司; 病原菌:匍匐翦股颖褐斑病病原菌为立枯丝核菌(Rhizoctonia solani), 购自中国科学院菌种保存中心。
试验于2017年5-8月在甘肃农业大学草业学院温室内进行。选取颗粒饱满的匍匐翦股颖种子, 室温下蒸馏水过夜浸泡后, 经75%的乙醇浸泡60 s, 然后经20% NaClO消毒15 min, 然后用无菌水冲洗4~5次, 最后用吸水纸将水吸干, 种植于含灭菌后营养土(营养土∶ 蛭石=2∶ 1, pH 6.5)的育苗钵(12 cm× 10 cm)中, 每盆0.2 g。
试验共设置4个激素组合, 即EBR+SA、ET+SA、ET+EBR、ET+EBR+SA(EBR 0.03 mg· L-1, SA 1 mmol· L-1, ET 0.03 μ g· mL-1, 1∶ 1组合)以及清水对照(CK)处理和3个连续喷施诱导时间, 即5、7、10 d, 共15个处理, 每个处理3次重复, 每个重复5盆, 待幼苗长出第3片真叶时, 根据不同激素组合和连续喷施诱导时间处理匍匐翦股颖幼苗, 每盆喷施15 mL外源复合激素。诱导处理后同时接种病原菌, 立枯丝核菌于PDB液体培养基(马铃薯块200 g· L-1, 葡萄糖20 g· L-1, 自然pH)中震荡(25 ℃, 120 r· min-1)培养2~3 d, 培养好的菌种配置成浓度为OD340=0.6~0.8的菌丝悬浮液, 采用喷雾法接种, 每盆20 mL。接种后第15天, 取样和测定分析。
1.3.1 病情指数及防治效果 接种后第15天, 统计病叶率并计算病情指数, 匍匐翦股颖褐斑病分级标准[15]为:0级:无症状; 1级:病斑面积占全株叶片面积的10%以下; 2级:病斑面积占全株叶片面积的10%~40%; 3级:病斑面积占全株叶片面积的40%~60%; 4级:病斑面积占全株叶片面积的60%~80%; 5级:病斑面积占全株叶片面积的90%以上。
防治效果=(对照发病率-处理发病率)/对照发病率× 100%
1.3.2 抗氧化酶活性测定 粗酶液的提取, 称取匍匐翦股颖叶片0.5 g放入研钵中, 加入液氮研磨组织破碎后加入2 mL预冷的磷酸缓冲液和2%聚乙烯吡咯烷酮(polyvinyl pyrrolidone, PVP)充分研磨, 然后转入离心管中, 用2 mL缓冲液充分清洗研钵, 转入离心管中, 4 ℃下15000 r· min-1离心20 min, 所得上清液即为粗酶提取液。将粗酶液分装入各管中进行抗氧化酶的活性和抗氧化物质含量测定。超氧化物歧化酶(superoxide dismutase, SOD)采用氮蓝四唑显色法(nitro blue tetrazolium chloride mono-hydrate, NBT)测定[16]; 过氧化氢酶(catalase, CAT)活性采用紫外比色法测定[16]; 过氧化物酶(peroxidase, POD)活性采用愈创木酚法测定[16]; 抗坏血酸过氧化物酶(ascorbate peroxidase, APX)活性的测定参照Nakano等[17]的方法; 脱氢抗坏血酸还原酶(dehydroascorbate reductase, DHAR)活性的测定参照Krivosheeva等[18]的方法; 谷胱甘肽还原酶(glutathione reductase, GR)活性测定参照 Foyer等[19]的方法。
1.3.3 非酶抗氧化物质含量测定 粗酶液提取同1.3.2。脱氧抗坏血酸(reduced ascorbic acid, AsA)和氧化型抗坏血酸(dehydroascorbic acid, DHA)测定参照Jin等[20]的方法; 氧化型谷胱甘肽(oxidized glutathione, GSH)和还原型谷胱甘肽(reduced glutathione, GSSG)参照Gossett等[21]的方法测定。
SPSS 19.0软件进行统计分析, 采用单因素ANOVA进行分析处理, Duncan’ s新复极差法进行显著性方差分析; 采用Excel 2003进行绘图与数据处理。
与对照(CK)相比, 不同激素组合和连续诱导时间处理的匍匐翦股颖幼苗抗病性明显增强(表1)。对照(CK)的病情指数随连续诱导时间的增加呈上升趋势; EBR+SA处理的病情指数随连续诱导时间的增加表现为先降后升, 而防治效果表现为先升后降, 当连续诱导时间为7 d时, 病情指数最低, 为14.93, 防治效果达33.53%, 但与连续诱导时间为10 d的防治效果无显著差异(P> 0.05); ET+SA处理的病情指数随连续诱导时间的增加呈上升趋势, 而防治效果表现为先升后降, 当连续诱导时间为5 d时, 病情指数最低, 为15.93, 但与连续诱导时间为7 d的病情指数无显著差异(P> 0.05), 连续诱导时间为7 d的防治效果最高, 为28.70%, 但与连续诱导时间为10 d的防治效果无显著差异(P> 0.05); ET+EBR处理的病情指数随连续诱导时间的增加呈先降后升趋势, 而防治效果表现为先升后降, 当连续诱导时间为7 d时, 病情指数最低为13.36, 防治效果为所有处理中最佳, 达40.51%; ET+EBR+SA处理的病情指数随连续诱导时间的增加呈上升趋势, 最低防治效果处理为连续诱导7 d, 但3种连续诱导时间处理的防治效果无显著差异(P> 0.05)。
| 表1 外源复合激素对匍匐翦股颖抗褐斑病的诱导效应 Table 1 Effect of composite exogenous hormone on creeping bentgrass against R. solani |
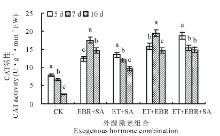
2.2.1 对SOD活性的影响 随着连续诱导时间的增加, 对照(CK)处理的幼苗叶片SOD活性略有下降(图1), 但无显著差异(P> 0.05); EBR+SA处理的幼苗叶片SOD活性随着连续诱导时间的增加呈先上升后下降趋势, 当连续诱导时间为7 d时, 叶片SOD活性最大, 为124.78 U· g-1 FW, 较CK提高了70.58%; ET+SA处理时当连续诱导时间为5 d时, 叶片SOD活性达109.49 U· g-1 FW, 较CK提高了49.70%, 之后随连续诱导时间的增加呈下降趋势; ET+EBR处理的SOD活性随着连续诱导时间的增加表现为先升后降, 当连续诱导时间为7 d时, 叶片SOD活性最大, 为168.63 U· g-1 FW, 较CK提高了130.53%, 当诱导时间为10 d时, SOD活性显著下降, 较7 d处理下降了45.18%; ET+EBR+SA处理的SOD活性变化与ET+SA处理相同, 随连续诱导时间的增加呈下降趋势, 当连续诱导时间为5 d时, SOD活性最大, 为112.39 U· g-1 FW, 较CK提高了53.66%。
2.2.2 对CAT活性的影响 与对照(CK)处理相比, 不同激素组合和连续诱导时间处理的匍匐翦股颖幼苗叶片CAT活性均不同程度上升(图2); 对照(CK)处理的幼苗叶片CAT活性随连续诱导时间的增加而显著下降, 连续诱导10 d时的CAT活性较5 d处理的下降了67.69%; EBR+SA处理的CAT活性呈先升后降趋势, 当连续诱导时间为7 d时, CAT活性最大, 较CK提高了166.82%; ET+SA处理CAT活性随连续诱导时间的增加呈逐渐下降趋势, 连续诱导5 d时CAT活性最大, 达13.45 U· g-1· min-1 FW; ET+EBR处理的CAT活性随连续诱导时间的增加表现为先升后降, 连续诱导7 d时CAT活性最大, 达19.45 U· g-1· min-1 FW, 较CK提高了197.40%; ET+EBR+SA处理的CAT活性变化呈下降趋势, 连续诱导5 d时CAT活性最大, 为18.68 U· g-1· min-1 FW, 7 d处理和10 d处理的CAT活性差异不显著(P> 0.05)。
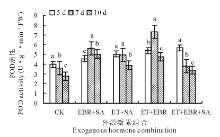
2.2.3 对POD活性的影响 随着连续诱导时间的增加, 对照(CK)的POD活性呈下降趋势(图3), 连续诱导10 d时的POD活性较5 d处理的下降了30.90%; EBR+SA处理的POD活性变化趋势同SOD和CAT, 呈现上升后下降趋势, 连续诱导7 d时POD活性最大, 较CK提高了56.23%; ET+SA处理的POD活性随连续诱导时间的增加呈逐渐下降趋势, 连续诱导5 d时POD活性最大, 为5.01 U· g-1· min-1 FW, 但与连续诱导7 d时的POD活性无显著差异(P> 0.05); ET+EBR处理的POD活性随着连续诱导时间的增加表现为先升后降, 当连续诱导时间为7 d时, 叶片POD活性最大, 为7.28 U· g-1· min-1 FW, 较CK提高了101.66%; ET+EBR+SA处理的POD活性变化呈下降趋势, 连续诱导5 d时POD活性最大, 为5.68 U· g-1· min-1 FW。
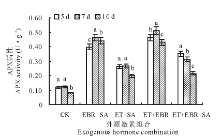
2.3.1 对APX活性的影响 与对照(CK)相比, 喷施激素处理的幼苗叶片APX活性均显著升高, 并随连续诱导时间的增加, 不同激素组合的幼苗叶片的APX活性变化趋势各不相同(图4)。对照(CK)处理中, 连续诱导时间为5和7 d时APX活性无显著差异(P> 0.05), 但连续诱导10 d时的APX活性显著下降(P< 0.05); EBR+SA处理的APX活性呈先升后降趋势, 连续诱导时间为7 d时APX活性最大, 与CK相比提高了276.42%; ET+SA处理在连续诱导5和7 d时的APX活性差异不显著(P> 0.05), 分别较CK提高了119.33%和115.45%, 当在连续诱导10 d处理时, APX活性显著下降; ET+EBR处理在连续诱导7 d时APX活性提高幅度最大, 较CK提高了316.26%; ET+EBR+SA处理的APX活性变化呈下降趋势, 连续诱导5 d时APX活性最大, 达0.351 U· g-1, 与CK相比提高了194.96%。
 | 图1 不同诱导处理对匍匐翦股颖幼苗SOD活性的影响Fig.1 Effect of different inducement treatments on SOD enzyme activity in creeping bentgrass |
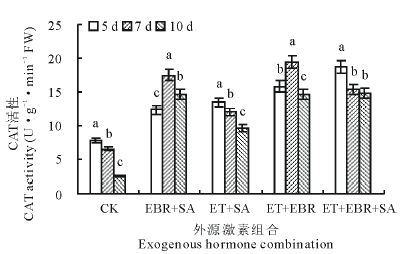 | 图2 不同诱导处理对匍匐翦股颖幼苗CAT活性的影响Fig.2 Effect of different inducement treatments on CAT enzyme activity in creeping bentgrass |
同一处理不同时间下小写字母表示在0.05水平上差异显著。下同。Different lowercase letters in same treatment and different time indicate significant difference at 0.05 level. The same below.
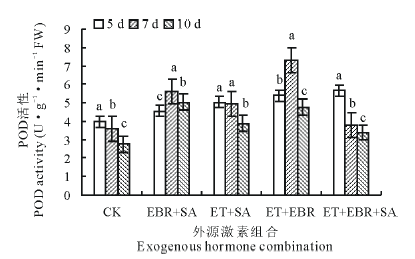 | 图3 不同诱导处理对匍匐翦股颖幼苗POD活性的影响Fig.3 Effect of different inducement treatments on POD enzyme activity in creeping bentgrass |
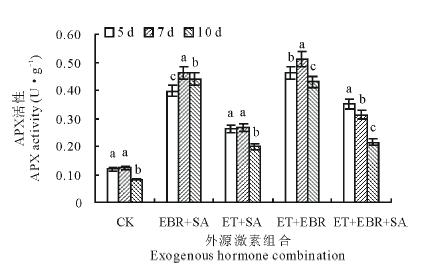 | 图4 不同诱导处理对匍匐翦股颖幼苗APX活性的影响Fig.4 Effect of different inducement treatments on APX enzyme activity in creeping bentgrass |
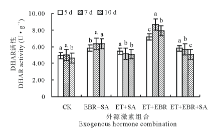
2.3.2 对DHAR活性的影响 由图5可知, 对照(CK)的幼苗叶片DHAR活性在连续诱导5和7 d处理时差异不显(P> 0.05), 当连续诱导10 d时, DHAR活性呈下降趋势; EBR+SA处理的DHAR活性呈上升趋势, 连续诱导7 d时DHAR活性达最大, 但与10 d处理时无显著差异(P> 0.05); ET+SA处理的DHAR活性随连续诱导时间的增加呈下降趋势, 且连续诱导7和10 d处理时差异不显著(P> 0.05), 各诱导时间处理较CK相比提高程度不明显; ET+EBR处理的DHAR活性同APX活性的变化, 呈先升后降趋势, 在连续诱导7 d时达最大, 较CK提高了41.95%; ET+EBR+SA处理的DHAR活性变化随连续诱导时间的增加呈下降趋势, 连续诱导5 d时DHAR活性最大, 达5.789 U· g-1, 与CK相比提高了14.67%。
 | 图5 不同诱导处理对匍匐翦股颖幼苗DHAR活性的影响Fig.5 Effect of different inducement treatments on DHAR enzyme activity in creeping bentgrass |
2.3.3 对GR活性的影响 随连续诱导时间的增加, 对照(CK)的GR活性呈先升后降趋势, 在连续诱导7 d时GR活性最大, 为0.356 U· g-1(图6); EBR+SA处理的GR活性由连续诱导5 d增加到7 d时略有上升, 但差异不显著(P> 0.05), 连续诱导10 d时GR活性显著下降; ET+SA处理的GR活性在诱导5 d时最大, 达0.489 U· g-1, 但与7 d处理的差异不显著, 10 d处理时显著下降; ET+EBR处理的GR活性随连续诱导时间的增加呈先升后降趋势, 在7 d处理时达最大, 较CK提高了80.34%, 提升幅度为各处理最大; ET+EBR+SA处理的GR活性随连续诱导时间的增加呈下降趋势, 但连续诱导5和7 d处理的差异不显著(P> 0.05), 当连续诱导10 d时其活性显著下降。
 | 图6 不同诱导处理对匍匐翦股颖幼苗GR活性的影响Fig.6 Effect of different inducement treatments on GR enzyme activity in creeping bentgrass |
2.3.4 对AsA和DHA含量及AsA/DHA的影响 与对照(CK)相比, 喷施激素处理的幼苗叶片AsA含量均显著升高, 但各处理随连续诱导时间的不同而变化趋势各不相同(表2)。对照(CK)处理的AsA含量随诱导时间的增加呈先升后降趋势, 在连续诱导7 d处理时AsA含量达最大, 为336.39 μ g· g-1; EBR+SA处理的AsA含量随诱导时间的增加呈先升后降趋势, 在连续诱导7 d处理时AsA含量达最大, 较CK提高了56.97%; ET+SA处理的AsA含量随诱导时间的增加呈下降趋势, 在连续诱导5 d处理时AsA含量达最大, 为499.65 μ g· g-1, 较CK提高了57.12%; ET+EBR处理的AsA含量在连续诱导7 d时较各处理达最大, 为654.74 μ g· g-1, 较CK提高了94.64%; ET+EBR+SA处理的ASA含量随诱导时间的增加呈下降趋势, 在连续诱导5 d处理时AsA含量达最大, 为536.41 μ g· g-1, 较CK提高了68.68%。
对照(CK)处理的DHA含量随诱导时间的增加呈先降后升趋势, 在连续诱导7 d处理时DHA含量积累最少, 为1031.44 μ g· g-1; EBR+SA处理的DHA含量呈上升趋势, 在连续诱导5 d处理时DHA含量积累最小, 但与连续诱导7 d处理时差异不显著; ET+SA处理的DHA含量呈先升后降趋势, 在连续诱导5 d处理时积累量最小, 为1014.42 μ g· g-1; ET+EBR处理的DHA随连续诱导时间的增加DHA含量积累变化较小, 各连续诱导时间处理均无显著差异, 在连续诱导7 d处理时DHA积累量最少, 为1062.88 μ g· g-1; ET+EBR+SA处理的DHA含量呈逐渐上升趋势, 在连续诱导5 d处理时DHA含量积累最小, 为1076.88 μ g· g-1。
对照(CK)处理的AsA/DHA随连续诱导时间的增加呈先升后降趋势, 连续诱导7 d处理时比值最大, 为0.33; EBR+SA处理的AsA/DHA在连续诱导7 d处理时最大, 为0.53, 较CK提高了60.61%, 连续诱导5和10 d处理的AsA/DHA无显著差异; ET+SA处理的AsA/DHA随连续诱导时间的增加而显著下降, 在连续诱导5 d处理时最大, 为0.49; ET+EBR处理的AsA/DHA在连续处理7 d时为各处理中最大值, 达0.62, 较CK提高了87.88%, 随后呈下降趋势; ET+EBR+SA处理在连续诱导5 d时AsA/DHA最大, 为0.50, 较CK提高了66.67%, 当连续诱导时间增加时呈下降趋势。
2.3.5 对GSH和GSSG含量及GSH/GSSG的影响 各激素处理较对照(CK)相比GSH含量均有所增加(表3)。对照(CK)的GSH含量在连续诱导7 d时达最大, 为9.66 μ g· g-1, 但与连续诱导5 d时的GSH含量差异不显著(P> 0.05), 连续诱导10 d时开始呈下降趋势; EBR+SA处理的GSH含量随连续诱导时间的变化呈先升后降趋势, 7 d处理时达最大, 为12.58 μ g· g-1, 较CK提高30.23%; ET+SA处理随连续诱导时间的增加, GSH含量呈下降趋势, 其在5 d处理时含量最高, 为12.07 μ g· g-1, 较CK提高了28.95%; ET+EBR处理在连续诱导7 d时GSH含量达各处理的最大值, 为13.67 μ g· g-1, 较CK提高41.51%, 随后呈下降趋势; ET+EBR+SA处理的GSH含量同AsA含量变化趋势相同, 随连续诱导时间的增加而逐渐下降, 在5 d处理时GSH含量最高, 为13.00 μ g· g-1, 较CK提高了38.89%。
| 表2 不同诱导处理对匍匐翦股颖幼苗AsA、DHA含量及AsA/DHA的影响 Table 2 Effect of different inducement treatments on contents of AsA, DHA and AsA/DHA in creeping bentgrass |
对照(CK)处理的GSSG含量在连续诱导7 d时积累量最少, 为1.30 μ g· g-1, 5和10 d处理时的GSSG含量积累均高于7 d处理; EBR+SA处理的GSSG积累量随连续诱导时间的增加而不断增加, 5 d处理时积累量最小, 为1.28 μ g· g-1; ET+SA处理的GSSG积累量在5 d处理时积累量最小, 为1.29 μ g· g-1, 但与7 d处理的无显著差异, 10 d处理时GSSG积累量显著增加; ET+EBR处理在连续诱导时间为5 d时, GSSG积累量最小, 随连续诱导时间的增加GSSG含量呈增加趋势, 7和10 d处理的GSSG含量差异不显著(P> 0.05); ET+EBR+SA处理的GSSG含量随连续诱导时间的增加而呈上升趋势, 5和7 d处理差异不显著(P> 0.05), 10 d处理时GSSG积累量显著增加。
对照(CK)处理的GSH/GSSG随连续诱导时间的增加呈先升后降趋势, 连续诱导7 d处理时比值最大, 为7.43; EBR+SA处理的GSH/GSSG在连续诱导7 d处理时最大, 为9.09, 较CK提高了22.34%, 但与连续诱导5 d时的GSH/GSSG差异不显著; ET+SA处理的GSH/GSSG随连续诱导时间的增加而显著下降, 在连续诱导5 d处理时最大, 为9.36, 较CK提高了37.04%; ET+EBR处理的GSH/GSSG在连续处理7 d时为各处理中最大值, 达9.88, 较CK提高了32.97%; ET+EBR+SA处理的GSH/GSSG随连续诱导时间的增加而逐渐降低, 在5 d处理时GSH/GSSG最大, 为9.56。
| 表3 不同诱导处理对匍匐翦股颖幼苗GSH、GSSG含量及GSH/GSSG的影响 Table 3 Effect of different inducement treatments on contents of GSH, GSSG and GSH/GSSG in creeping bentgrass |
植物诱导抗性是利用物理、化学及生物的方式使植物受诱导因子刺激后自身所产生的抗性, 从而达到防治植物病害的作用, 然而不同的诱导剂对同一植物的防治效果存在差异, 这主要与诱抗剂的诱导抗病机理及植物自身的代谢途径等有关, 对于某一固定植物, 存在着适合该植物的一种或几种有效诱抗剂[22]。Lee等[23]使用苯并噻二唑(benzothiadiazole, BTH)诱导匍匐翦股颖品种“ Crenshaw” 、“ Penncross” 和“ Providence” 抗币斑病, 但对“ L-93” 是没有效果的。房媛媛等[24]研究发现以不同浓度的2, 3-丁二醇(2, 3-butanediol, 2, 3-BD)和水杨酸处理匍匐翦股颖后接种尖孢镰刀菌, 水杨酸各处理中防治效果最高为11.25%, 而250 μ mol· L-1的2, 3-丁二醇的防治效果可高达78.26%。诱导剂诱导植物产生抗病性, 均需要一定时间激发寄主植物体内产生系列生理反应, 不同诱导物或同一诱导物对不同寄主和病害的诱导抗病时间持续性可能有所不同[25]。向妙莲等[25]研究表明, 采用0.1 mmol· L-1的茉莉酸甲酯(methyl jasmonate, MeJA)于接种青枯病前0~96 h喷雾处理辣椒幼苗均具有一定的防治效果, 但以接种前48 h时处理的防治效果最好。刘兴菊等[26]研究发现, 利用2, 3-丁二醇对匍匐翦股颖多次诱导后接种立枯丝核菌, 其中2次诱导处理下效果最好, 病情指数最低。本研究结果表明, 不同的复合激素及连续诱导时间对匍匐翦股颖抗褐斑病的效果不同, 其中ET+EBR连续诱导处理7 d的病情指数最低, 为13.36, 防治效果最佳, 为40.51%。分析现有研究诱导匍匐翦股颖抗性的试验结果发现, 4种外源激素组合对匍匐翦股颖褐斑病均有一定的诱导防治效果, 但诱导机制可能存在差异, 导致抗性差别显著; 另外, 植株抗病性的激发是一个相对缓慢的过程, 短时间内或较少次数的诱导对植株抗病能力而言没有明显提高, 但不同诱导物或同一诱导物对不同寄主和病害的诱导抗病时间和诱导次数也可能有所不同。
在植物的诱导抗病过程中伴随着多种物质的代谢, 而这些过程中关键因子是催化代谢反应的酶, 其中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)是与植物抗病相关的抗氧化关键酶, 植物通过调节这些抗氧化关键酶活性, 保持活性氧代谢平衡, 以减缓细胞内活性氧自由基积累和膜脂过氧化伤害[27]。SOD作为植物细胞防御酶系统中的重要成分之一, 主要的功能便是歧化负氧离子, 产生H2O2和O2, 减少活性氧对植物体细胞的伤害; CAT和POD是植物体内常见的氧化还原酶, 是细胞内重要的内源活性氧清除剂[28], 从而使植物细胞能够缓解一定程度上的氧化损伤。许多研究表明, 植物经诱导剂处理后SOD、CAT、POD等的活性都显著增加[29, 30], 并且这些酶活性的升高常被用来作为植物抗病性的重要指标[31]。而关于SOD、POD、CAT等酶活性与植物抗病性的关系也存在较多争议[32]。本研究分析比较匍匐翦股颖幼苗经不同复合激素处理后接种立枯丝核菌, 其叶片关键抗氧化酶活性的变化, 发现各复合激素可显著提高SOD、CAT、POD等酶活性, 且于最佳连续诱导时间处理下酶活性最强, 证实了SOD、POD、CAT的活性增强有助提高植株的抗病性, 说明复合激素诱导匍匐翦股颖抗褐斑病, 可能与其激发植株体内抗氧化酶活性并调控活性氧代谢平衡有关。
植物在减缓或抵御细胞伤害时常调运酶促和非酶促两类系统来清除活性氧, 其中抗坏血酸过氧化物酶(APX)、脱氢抗坏血酸还原酶(DHAR)和谷胱甘肽还原酶(GR)是抗坏血酸-谷胱甘肽循环(AsA-GSH)活性氧清除系统的重要酶组成, AsA和GSH是重要的非酶抗氧化物质[33, 34, 35]。APX在AsA-GSH循环中通过AsA来清除H2O2, 在H2O2清除过程中起主要作用, AsA被氧化形成单脱氢抗坏血酸(monodehydroascorbate, MDHA), MDHA或在单脱氢抗坏血酸还原酶(monodehydroascorbate reductase, MDAR)作用下再生形成AsA, 或通过非酶促歧化形成氧化型抗坏血酸(DHA), DHA在DHAR作用下再生形成AsA, GR通过催化GSSG生成还原态的GSH。AsA是植物体常见的一种非酶促抗氧化剂, 在AsA-GSH循环中能够有效清除H2O2, 同时还可直接清除
以EBR、SA和ET相互组合为外源复合激素, 在不同连续诱导时间下处理匍匐翦股颖幼苗, 可以降低匍匐翦股颖褐斑病的病情指数, 提高匍匐翦股颖对褐斑病的抗性, 并增强SOD、CAT、POD和AsA-GSH循环中抗氧化酶活性及AsA和GSH的再生能力, 使活性氧的清除能力增强, 降低膜脂过氧化程度, 从而维持细胞结构的稳定性, 提高匍匐翦股颖的抗病性。综合不同外源复合激素和连续诱导时间对匍匐翦股颖抗褐斑病各指标的分析, 本试验认为, 外源复合激素ET+EBR连续诱导处理7 d时对匍匐翦股颖抗褐斑病的诱导效果最好。但是, 外源复合激素诱导植物抗病的响应机制及其信号途径还较为复杂, 还需进一步深入研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|