
{kind=link}
{kind=link}
{kind=link}
象草不同辐射剂量诱变系表型及遗传变异研究
[武炳超 , 张欢, 童磊, 杜昭昌, 胡家菱, 陈燚, 张新全, 刘伟, 黄琳凯
, 张欢, 童磊, 杜昭昌, 胡家菱, 陈燚, 张新全, 刘伟, 黄琳凯* ]
, 张欢, 童磊, 杜昭昌, 胡家菱, 陈燚, 张新全, 刘伟, 黄琳凯]
|
|


作者简介:武炳超(1994-),男,内蒙古巴彦淖尔人,在读硕士。E-mail: wubingchao94@163.com
以象草种茎为材料,利用10、20和30 Gy剂量的60Co-γ射线照射种茎,筛选适宜诱变剂量并研究其表型性状变异和分子水平上的遗传变异,为象草辐射诱变育种提供理论和技术基础。结果表明,辐射处理后的植株有矮化的趋势,其中分蘖数、茎节数和叶长对辐射最敏感,容易发生变异;在所有诱变系材料中,F30-39和F30-41聚为一类,与对照材料差异最大,是显著矮小化的植株。30 Gy诱变系与未辐射的材料间的遗传相似系数在0.3793~0.9655,平均为0.8276,而差异位点数在4~66个,平均为19.3个,两项指标均大于10和20 Gy诱变系群体。综合考虑辐射后植株存活率与遗传变异结果,选定30 Gy为最适宜的象草种茎辐射诱变剂量。
, ZHANG Huan, TONG Lei, DU Zhao-chang, HU Jia-ling, CHEN Yi, ZHANG Xin-quan, LIU Wei, HUANG Lin-kai
In order to provide a theoretical basis for mutation-induced breeding of Pennisetum purpureum, stems of P. purpureum were irradiated with60Co-γ rays at differing doses of 10, 20 and 30 Gy. Then, we screened the resulting mutants and defined their phenotypic variation, and identified the genetic differences at a molecular level, using SSR. The irradiated plants showed a tendency to dwarfism. The number of tillers, stems and the length of leaves were the traits most sensitive to radiation and most easily mutated. Among all the mutants, two designated F30-39 and F30-41 were significantly shortened plants, and were determined by cluster analysis to belong to a group which had the greatest genetic difference from the control material. The genetic similarity coefficient between controls and “30 Gy” mutants ranged from 0.3793 to 0.9655, with the average being 0.8276, while the number of mutation sites ranged between 4 and 66, with an average of 19.3. Both indices were greater than those of “10 and 20 Gy” mutants. Considering the survival rate and genetic mutation frequency of irradiated plants, 30 Gy was selected as the most suitable radiation dose level for mutagenesis.
狼尾草属(Pennisetum)隶属禾本科黍亚科, 蒺藜亚族, 为一年生或多年生禾本科牧草, 主要分布于热带、亚热带和温带地区, 全世界约140种, 多数原产于非洲[1]。我国人工栽培利用的品种主要有多年生的象草(P. purpureum)、一年生的美洲狼尾草(P. americanum)及二者之间的杂交种等。
象草, 别名紫狼尾草。20世纪40年代初, 我国的四川、广东从印度、缅甸引入试种, 后传入湖南、江苏、福建等地, 目前我国南方各省均有栽培[2]。象草是一种茎秆粗高、含糖量大、粗蛋白和无氮浸出物含量高的优良牧草[3]。1975年, 广东、广西等地一些畜牧场大面积种植象草饲喂奶牛, 取得良好成效[4]。象草因其生长速度快、适口性好、营养价值丰富等特点被多种家畜喜食, 其多作为青刈饲料, 也可以青贮或者调制干草[5]。象草不仅仅是一种优良的牧草, 它还是一种重要的生物质能源植物。美国在20世纪80年代展开了将象草作为能源植物的研究工作, 证明了其可以用于乙醇、沼气和电能的生产[6, 7, 8, 9]。除此之外, 象草还通过其高大的植株、丰富的叶片、分蘖以及发达的根系保护了土表, 发挥着重要的生态效益[10]。近年来, 因其株型优美、抗逆性强、管理粗放、维护费用低等特点, 已成为一种新型的园林造景植物[11]。
目前我国的国审狼尾草品种中有6个是象草品种, 其中4个为引进品种, 品种资源匮乏已经成为象草进一步育种和利用的障碍。象草为4倍体, 有28条染色体[12]。由于不能形成花粉或者雌蕊发育不良, 因而一般不结实或结实率低, 种子活性低, 生产多用种茎繁殖, 种质资源遗传多样性低, 限制了其育种利用[13, 14]。诱变育种技术作为一种有效创造新种质的育种手段, 已经广泛应用于植物育种研究中[15]。其中γ 射线可以通过辐射能量使生物体内各种分子产生电离和激发, 形成自由原子或者基团, 他们相互反应并与周围大分子核酸和蛋白质起反应, 引起染色体结构变异, 主要为易位、倒位和缺失[16, 17]。为了创制新的象草种质资源, 丰富象草种质资源多样性, 本实验用60Co-γ 射线照射受体材料, 对诱变系材料表型性状变异进行初步研究, 并辅以分子标记的方法比较诱变系与对照材料之间的遗传差异, 探究辐射诱变对象草遗传变异的影响, 并确定最适宜的辐射诱变剂量, 为以后的诱变系选育和品种改良提供理论和技术支撑。
种植在四川农业大学成都崇州实验基地的象草061023003, 来自中国热带农业科学院牧草种质基因库, 均为同一无性系扩繁所得, 以保证所选材料遗传背景相同。挑选长势相近且健康的植株割取种茎, 种茎标准为含有2~3个茎节。
1.2.1 象草临界剂量与半致死剂量确定 2016年4月, 将待辐射的种茎送往四川省农科院辐射中心, 采用60Co-γ 射线进行照射, 在运送过程中通过洒水保持种茎湿润以减弱缺水对种茎的伤害。结合以前的研究, 本试验共设置了3个剂量梯度:10、20和30 Gy, 剂量率1 Gy· min-1, 以没有经过辐射处理的材料作为对照。每个剂量梯度分别辐射66个种茎。将辐射后的种茎种植于四川农业大学崇州实验基地, 茎秆倾斜插入土壤, 以土壤覆盖一个茎节为准, 株距1 m, 行距2 m, 常规大田栽培管理, 一个月后统计成活率。其中, 存活率为50%时的剂量为半致死剂量, 存活率为40%时的剂量为临界剂量[18]。
1.2.2 植株表型性状测定 等待存活的植株生长成熟后, 对每个诱变系群体随机选取15株进行形态指标测定。共测定6个指标, 包括株高, 分蘖数, 茎节数, 叶长, 叶宽及茎粗。其中叶长、叶宽两个指标选择从下至上倒数第2片叶子进行测量, 叶宽选择叶片最宽处测量。除自然高度和分蘖数以外, 其余指标均重复测量5次。
1.2.3 SSR分子标记 3个诱变系群体选择的植株同表型分析的植株, 对每个存活材料随机挑选2片健康的叶片, 采用DNA提取试剂盒DNeasy Plant Mini kit (Qigene公司, 美国)提取DNA。用分光光度计检测DNA浓度, 合格DNA样品用TE稀释至20 ng· μ L-1。PCR反应体系为15 μ L, 包括DNA 1.5 μ L(20 ng· μ L-1), Taq酶0.3 μ L, Master Mix 7.5 μ L, 上游引物和下游引物各0.6 μ L(10 pmol· μ L-1), ddH2O 4.5 μ L。PCR反应程序为:94 ℃预变性5 min, 94 ℃变性30 s, 58 ℃退火45 s, 72 ℃延伸1 min, 共35个循环; 最后72 ℃延伸7 min, 4 ℃保存。得到的扩增产物用8%聚丙烯酰胺凝胶在350 V电压下电泳155 min, 再用浓度为1 g· L-1的AgNO3染色15 min, 显影后照相保存。引物由本课题组象草转录组测序数据开发, 序列交由上海生工生物工程技术服务有限公司完成。以3个诱变系群体中形态变化差异较大的单株DNA为模板, 从50对SSR引物中筛选出20对多态性高、特异性强、条带清晰的引物。
对于形态指标采用Microsoft Excel 2007软件处理实验数据, 采用统计分析软件SPSS 19.0进行方差分析。对电泳得到的胶片进行标准化处理:在相同迁移位置, 对稳定且清晰的条带进行统计, 有带记为“ 1” , 无带记为“ 0” , 建立原始矩阵。根据表征矩阵, 利用Excel 2007统计SSR扩增产物的条带总数(total number of bands, TNB)和多态性条带数(number of polymorphic bands, NPB), 计算多态性条带所占的比率(percentage of polymorphic bands, PPB)和引物多态性信息含量(polymorphism information content, PIC)。使用Popgene 32软件计算3个诱变系群体的基因多样性、Shannon信息指数、多态性位点及多态性百分率, 利用NTSYSpc 2.1(Version 2.10s)软件计算遗传相似系数(genetic similarity coefficient, GSC)并用Past 3绘制UPGMA聚类图, 以探究诱变系与对照材料间的亲缘性。
经过60Co-γ 射线辐射后, 得到3个诱变系群体, 植株存活率随着辐射剂量的增加而降低。当剂量为10 Gy时, 存活植株数为62株, 存活率为93.94%, 辐射剂量为20 Gy时, 存活植株数为49株, 存活率为74.24%, 而当剂量为30 Gy时, 植株存活数为28株, 存活率为42.42%, 而对照组植株存活率为100%, 预测30 Gy可能是象草的诱变适宜剂量。
在3个诱变系群体中分别随机选取15株进行形态指标的测量, 发现3个诱变系群体中均有一个或多个形态指标与对照组群体存在显著或极显著差异(表1)。其中, 10 Gy诱变系群体中, 仅有叶长一个指标与对照群体差异显著; 20 Gy诱变系群体中株高、叶长、叶宽3个形态指标与对照群体差异显著, 其中叶长和叶宽两个指标均达到极显著差异水平; 30 Gy诱变系群体中, 共有4个形态指标与对照群体差异显著, 其中株高和叶宽与对照群体差异极显著。
| 表1 不同剂量辐射诱变系形态变异 Table 1 The morphological variation of mutants by different dose of radiation |
进一步对3个群体各形态变异系数进行计算(表2), 结果表明, 除了叶长外, 其余形态变异系数均在30 Gy诱变系群体中最大, 说明该群体是差异最明显的诱变系群体。对3个群体中不同形态变异系数进行比较, 发现在10 Gy群体中最容易发生变异的形态依次为叶长、分蘖数、茎节数; 在20 Gy群体中, 最容易发生变异的形态依次为茎节数、叶长、分蘖数; 而在30 Gy群体中, 依次为茎节数、分蘖数、叶长。由此可见, 分蘖数、茎节数和叶长对60Co-γ 射线最为敏感, 容易在受到辐射后发生变异。
| 表2 各诱变系不同形态指标的变异系数 Table 2 The coefficient of variation of morphological in different mutants group (%) |
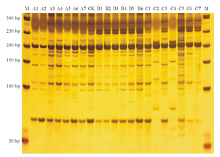
2.3.1 遗传多样性分析 筛选出的20对引物共扩增出116条清晰可见的条带, 扩增片段大小为50~300 bp(图1), 其中多态性条带91条, 多态性条带比率为78.45%。每对SSR引物扩增的条带数为3~8条, 平均条带数为5.8条, 多态性条带数为1~8条, 平均为4.6条。多态信息含量(PIC)在0.032~0.758, 平均为0.273, 其中引物c104424_g1的PIC最大, 引物c106150_g1的PIC最小(表3)。对3个诱变系群体的基因多样性、Shannon信息指数、多态性位点及多态性百分率进行比较(表4), 发现在30 Gy诱变系群体中的4项指标均高于另外两个诱变系群体, 说明30 Gy的60Co-γ 射线较另外两个剂量能引起更多的遗传变异, 更加丰富的基因多样性。
 | 图1 引物c100123_g1扩增产物电泳图Fig.1 The amplification product electrophoresis of primer c100123_g1 M代表Marker, A1~A7分别代表F10-41、F10-42、F10-43、F10-45、F10-49、F10-52、F10-54, B1~B6分别代表F20-40、F20-46、F20-47、F20-51、F20-53、F20-54, C1~C7分别代表F30-38、F30-39、F30-40、F30-41、F30-42、F30-43、F30-44, CK代表对照材料。M stands for Marker; A1-A7 represent F10-41、F10-42、F10-43、F10-45、F10-49、F10-52、F10-54, respectively; B1-B6 represent F20-40、F20-46、F20-47、F20-51、F20-53、F20-54, respectively; C1-C7 represent F30-38、F30-39、F30-40、F30-41、F30-42、F30-43、F30-44, respectively; CK stands for control material. |
| 表3 20对SSR引物序列及其扩增结果 Table 3 20 SSR primers used in this study and their amplification results |
| 表4 3个诱变系群体基于SSR标记的多态性分析 Table 4 Polymorphism analysis of three mutants based on 20 SSR |
2.3.2 遗传相似性及SSR标记位点差异分析 根据SSR引物扩增出的条带, 计算各诱变系群体与对照材料之间的遗传相似系数。从表5中可知, 对照材料与诱变系材料之间的遗传相似系数在0.3793~0.9741, 平均值为0.8565。3个诱变系群体与对照材料的遗传相似系数有一定差异, 在10 Gy诱变系群体中, 遗传相似系数在0.8017~0.9655, 平均为0.8856; 在20 Gy诱变系群体中, 遗传相似系数为0.7069~0.9741, 平均为0.8563; 在30 Gy群体中, 遗传相似系数为0.3793~0.9655, 平均为0.8276。说明30 Gy诱变系群体与对照材料遗传差异最大, 发生的变异最多。在所有的诱变系材料中, F30-41与对照材料的遗传相似系数最小, 为0.3793, 该材料与对照材料的遗传差异最大, 而F20-44与对照材料的遗传相似系数最大, 为0.9741, 说明与对照材料的遗传差异最小, 亲缘性最近。
| 表5 诱变系与对照材料间的遗传相似系数 Table 5 The genetic similarity coefficient between mutants and control materials |
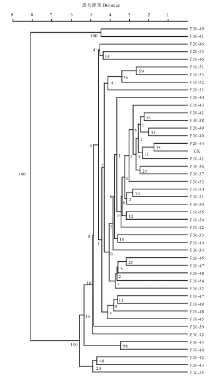
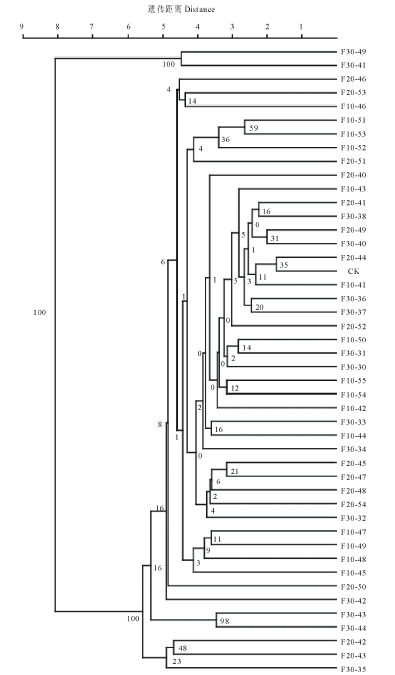 | 图2 诱变系的UPGMA聚类图Fig.2 UPGMA dendrogram of mutants |
根据扩增条带, 对诱变系材料和对照材料间的差异位点数进行统计(表6)。发现在10 Gy诱变系材料与对照材料间的差异位点数在4~23个, 平均13.3个; 20 Gy诱变系材料与对照材料间的差异位点数在3~34个, 平均16.7个; 而30 Gy诱变系材料与对照材料间的差异位点数在4~66个, 平均为19.3个。其中F30-41差异位点数达到66个, 差异位点百分率为56.9%, 说明该材料与对照材料间的遗传差异最大。其次是F30-39, 差异位点数达到65个, 差异位点百分率为56.0%; 而F20-44仅有3个差异位点, 差异位点百分率为2.6%, 说明该材料与对照材料间的遗传差异最小。该结果与遗传相似性分析结果一致, 均说明当象草茎秆受到30 Gy的60Co-γ 射线辐射时, 更易引起变异。
| 表6 对照材料与诱变系间的SSR标记差异 Table 6 The difference of SSR markers between the base material and its mutants |
进一步对辐射后诱变系材料进行非加权平均法(UPGMA)聚类分析, 以进一步探究不同辐射剂量所得诱变系材料与对照材料之间的亲缘关系。以所得条带原始矩阵构建亲缘关系系统树(图2)。由图可见, 所有材料可分为两类, F30-39(图3)与F30-41聚为一类且与对照材料遗传距离最远, 说明它们的变异程度最大。而F20-44与对照材料聚为一类, 说明几乎没有发生变异。而其他诱变系材料在不同距离与对照材料分开, 说明其受到60Co-γ 射线照射后, 均产生不同程度的变异。
 | 图3 突变体F30-39Fig.3 Mutants F30-39 |
不同的物种以及不同部位辐射对60Co-γ 射线的敏感性不同, 因此筛选最适宜的诱变剂量是辐射诱变育种的基础。王文恩等[19]使用60Co-γ 射线辐射野牛草(Buchloe dactyloides)干种子, 初步确定了促进野牛草干种子萌发的适宜辐射剂量为100~150 Gy, 而日本结缕草(Zoysia japonica)干种子辐射育种的半致死剂量为480 Gy[20]。以象草种茎为辐射材料进行诱变的报道较少, 采用3个剂量的60Co-γ 射线对象草种茎进行辐射, 发现30 Gy诱变系群体的存活率为42.42%, 接近临界剂量的存活率, 故推测30 Gy为象草种茎辐射诱变的适宜剂量。
象草不仅可以为草食动物提供大量优质牧草[21], 还具有一定的观赏效果[22]。对辐射诱变系与对照材料表型性状进行测定和分析是发现新品系的基础。曾捷等[23]用60Co-γ 射线辐射扁穗牛鞭草(Hemarthria compressa)后, 发现75%的诱变系叶片变小、株高变矮、茎变细。张彦芹等[24]也通过利用60Co-γ 射线辐射高羊茅(Festuca elata)分化苗, 得到了叶片变小、变细的高羊茅突变体。对狗牙根(Cynodon dactylon)进行辐射诱变后, 发现草层高度显著降低, 并且显著影响了叶宽、叶长及节间直径和密度[25]。本试验发现3个诱变系群体的株高、茎节数、茎粗、叶长及叶宽等形态指标与对照材料相比, 均有一个或多个指标显著或极显著减小, 其中分蘖数、茎节数和叶长对60Co-γ 射线最敏感, 变异系数较高。此结果与前人研究结果基本一致, 辐射诱变更倾向于产生植株变小的突变体[23, 24]。其中, F10-42和F10-49的株高和茎节数均大于对照材料, 是将来进行牧草育种的有潜力的新种质。
SSR标记是共显性分子标记, 多态性丰富、易于鉴别基因型, 被认为是最好的研究群体遗传变异的分子标记之一[26]。例如黄婷等[27]利用SSR标记对6个多花黑麦草(Lolium multiflorum)品种进行遗传差异分析并对品种进行有效的鉴定。本试验通过筛选得到的20对SSR引物, 对3个诱变系群体与对照材料的遗传差异进行了鉴定, 发现30 Gy诱变系群体的基因多样性为0.36, Shannon信息指数为0.51, 多态性位点89个, 多态位点百分率76.72%, 遗传相似系数为0.8276, 差异位点数19.3个, 均大于10和20 Gy诱变系群体, 表明该剂量更易产生丰富的遗传变异。对所有诱变系群体进行UPGMA聚类分析, 发现F30-39和F30-41聚为一类, 与对照材料距离最远, 说明其遗传差异最大, 而其他材料也与对照材料不同程度的分开, 说明不同剂量的60Co-γ 射线均有可能产生突变体。甘蔗(Saccharum officinarum)品种桂糖22号就是由60Co-γ 射线辐射诱变育成的[28]。刘天增等[29]利用辐射诱变方法筛选海滨雀稗(Seashore paspalum)突变体。甜菜(Beta vulgaris)的耐旱突变体也是由辐射得到的, 并且用ISSR分子标记进行了鉴定[30]。秦家友等[31]利用SSR标记对玉米(Zea mays)辐射诱变系进行遗传差异研究, 发现诱变系与基础材料间存在真实的遗传差异, 本研究结果与此一致。本研究的结果仅仅鉴定了不同诱变系群体与对照材料之间分子水平上的遗传差异, 确定了最适宜象草种茎辐射的60Co-γ 射线剂量, 对于突变体的突变原因以及可能发生突变的基因的鉴定还有待进一步深入研究。
不同剂量的60Co-γ 射线均可诱导象草发生变异, 其中分蘖数、茎节数和叶长对60Co-γ 射线最为敏感, 容易发生突变。结合不同剂量辐射诱变系表型及遗传变异的结果, 发现30 Gy的诱变效率最高, 可作为最适宜象草种茎辐射诱变的剂量。同时发现两个显著矮小化的突变体F30-39和F30-41, 可为象草后续育种及基因挖掘研究提供材料。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|