
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
激动素对盐胁迫下老芒麦幼苗端粒酶活性及生理特性的影响
[孙守江 , 师尚礼, 吴召林, 何丽娟, 金鑫, 祁娟
, 师尚礼, 吴召林, 何丽娟, 金鑫, 祁娟* ]
, 师尚礼, 吴召林, 何丽娟, 金鑫, 祁娟]
|
|


作者简介:孙守江(1990-),男,甘肃景泰人,在读硕士。E-mail: 931620185@qq.com
以2015年采自青藏高原高寒草地野生老芒麦为对象,研究了不同程度NaCl盐胁迫下激动素对老芒麦幼苗端粒酶活性及生理特性的影响,为土壤盐渍化地区的农业生产以及老芒麦人工草地改良提供科学依据。采用营养液砂培法,待幼苗生长至第3片叶抽出并刚展开时,对老芒麦幼苗进行不同浓度(0、50、100、150、200 mmol·L-1)的NaCl盐胁迫处理168 h,随后用不同浓度(0、5、10、20、30 mg·L-1)的激动素叶面喷施处理240 h并测定幼苗的端粒酶活性以及生理指标:叶绿素、游离脯氨酸(Pro)、丙二醛(MDA)、可溶性糖(SS)、可溶性蛋白(SP)含量。结果表明,随着盐胁迫的增强,端粒酶活性呈现出先上升后下降的趋势,在50 mmol·L-1的NaCl处理下,端粒酶活性达到最大值。当处理浓度大于50 mmol·L-1时,随着盐胁迫的进一步增强,端粒酶活性逐步减小,渗透调节物质游离脯氨酸、可溶性糖、可溶性蛋白含量均上升。可溶性糖在250 mmol·L-1盐浓度下增加量最显著( P<0.05),与对照相比增加了101.5%。0 mmol·L-1盐胁迫下经10 mg·L-1 的激动素处理后,MDA含量下降64.9%。较高浓度激动素对盐胁迫的缓解效果显著( P<0.05),250 mmol·L-1盐胁迫下经20 mg·L-1激动素处理的植株,叶绿素含量较对照增加了17.3%,可溶性糖在100 mmol·L-1盐胁迫下经20 mg·L-1激动素处理,其含量较对照增加了165.6%。结果表明外源激动素对盐胁迫具有一定的缓解作用,适宜浓度的盐胁迫诱导了细胞端粒酶活性,高浓度盐胁迫可能造成了老芒麦细胞的氧化损伤,最终导致端粒酶活性下降。
A study has been undertaken to investigate the effects of kinetin on the telomerase activity and physiological characteristics of wild Elymus sibiricus seedlings under different levels of salt stress in order to provide a scientific basis for agricultural production in areas with severe soil salinization and the improvement of artificial grassland. Wild E. sibiricus were collected from Qinghai-Tibet Plateau alpine grassland in 2015 and used as test material in sand culture experiments. When the third leaf of the seedlings extracted and unfolded, the seedlings were treated with sodium chloride (NaCl) solution at different concentration levels (0, 50, 100, 150, 200 mmol·L-1) for 168 hours. The seedlings’ leaves were then sprayed with kinetin solutions of different concentrations (0, 5, 10, 20, 30 mg·L-1) for 240 hours and measurements taken of telomerase activity and physiological parameters: chlorophyll, free proline (Pro), malondialdehyde (MDA), soluble sugar (SS), and soluble protein (SP) content. The results showed that with the increase of salt stress, telomerase activity firstly increased and then decreased, peaking under the 50 mmol·L-1 NaCl treatment. When NaCl concentrations were higher than 50 mmol·L-1, telomerase activity decreased and the content of free proline, soluble sugar and soluble protein increased with increasing salt stress, among which the SS increment (101.5%) of seedlings under 250 mmol·L-1 NaCl was significantly different from that of the control ( P<0.05). Following spraying with 10 mg·L-1 kinetin, the MDA content of seedlings treated with 0 mmol·L-1 NaCl decreased by 64.9%. High concentrations of kinetin can significantly relieve the salt stress of seedlings ( P<0.05), with chlorophyll content (under 250 mmol·L-1 NaCl stress) and SS content (under 100 mmol·L-1 NaCl stress) increasing by 17.3% and 165.6% respectively after spraying with 20 mg·L-1 kinetin.These results indicate that exogenous kinetin alleviates salt stress to some extent. Suitable NaCl concentrations induced telomerase activity, whereas high concentrations of NaCl stress may cause oxidative damage to E. sibiricus cells and result in decreased telomerase activity.
盐胁迫是限制植物生长、发育和分布的主要环境胁迫之一, 对农业生产造成了巨大的损失[1], 同时也是全球面临的资源以及生态环境问题, 全球的土地遭受盐渍化侵害严重, 中国约有25%的耕地面积已发生盐渍化, 严重制约了农作物的生长发育[1]。盐胁迫扰乱植物细胞内正常的生理生化反应, 打破细胞内部离子平衡状态, 对植物造成一定的损伤[2], 如细胞膜通透性增大、叶绿素降解、光合速率降低, 最终导致大量的活性氧(reactive oxygen species, ROS)积累[3]。高浓度的活性氧破坏植物细胞基本结构, 甚至对细胞内的生物大分子, 如蛋白质造成损伤, 严重时会导致DNA断裂以及蛋白质的三级结构发生变化, 失去应有的功能[3]。特殊的端粒结构序列导致其对活性氧超级敏感, 端粒DNA序列的断裂以及丢失导致端粒缩短和功能失常[4]。
端粒(telomere)是存在于真核细胞线状染色体末端的一小段DNA-蛋白质复合体, 它与端粒结合蛋白一起构成了特殊的“ 帽子” 结构, 其主要作用是保持染色体的完整性和控制细胞分裂周期[4]。端粒、着丝粒和复制原点是染色体保持完整和稳定的三大要素[5]。通常情况下, 细胞每分裂一次都会使端粒变短, 当缩短到一定程度时就会发出应急信号, 细胞停止分裂[5]。端粒酶是染色体末端的一种特殊结构, 它是由许多简单短重复序列和端粒结合蛋白(telomere end-binding protein, TEBP)组成[4]。它的主要作用就是维持端粒有足够的长度, 保证基因信息在每个复制周期中的准确性, 以防止染色体末端遗传信息的丢失[4]。端粒酶对DNA具有一定的修复作用以及在真核细胞中能够抵御活性氧带来的细胞氧化损伤[6], 通过对人逆转录酶(hTERT)的研究发现, 一定程度的DNA损伤可以增加hTERT的表达, 使得端粒酶活性增加[7], hTERT能够增加DNA的自我修复能力以及修复速度。同时DNA末端合成能力也增强, 进一步增加了DNA轻度损伤后植物细胞的存活概率。但是, 强氧化胁迫导致植物细胞端粒酶活性降低, 最终导致细胞死亡[7]。
关于植物细胞端粒系统对非生物胁迫响应的研究报道有限。Fojtová 等[8]用镉处理烟草(Nicotiana tabacum), 发现一定程度的镉胁迫导致烟草细胞氧化损伤, 当移除镉以后, 发现烟草细胞凋亡速率降低, 是因为烟草细胞受到氧化损伤以后具有一定程度的自我修复能力, 在移除胁迫初期端粒酶活性明显升高, 由此可以看出, 端粒酶在细胞受到氧化损伤以后, 在自我恢复过程中发挥着重要作用。张徐俞等[3]将沙冬青(Ammopiptanthus mongolicus)作为研究材料, 探究不同NaCl盐胁迫程度下沙冬青细胞端粒酶活性变化与染色体DNA稳定性的关系。发现重度胁迫处理初期, 端粒酶活性急剧增大, 随着处理时间的延长, 活性逐渐减小, 但是未见DNA降解的迹象; 当移除NaCl处理后, 端粒酶活性有所增加, 通过观察发现, DNA也未发现降解迹象, 一直保持稳定。由此可以看出端粒酶在抵御植物细胞氧化损伤、保持细胞遗传物质稳定性方面发挥着重要作用。吴晓飞等[2]用不同程度盐胁迫处理沙冬青愈伤组织细胞, 发现端粒酶对氧化损伤也具有一定的延缓和修复作用。
老芒麦(Elymus sibiricus)是一种适应性较强, 抗寒性较好的多年生中旱生植物, 具有粗蛋白含量高、适口性好和易栽培等优良特性, 因此在西北、华北、东北等地常常被认为是一种营养价值较高的优质牧草[9]。关于老芒麦抗旱抗寒方面已有大量报道, 在土壤遭受干旱盐渍化程度日趋严重的今天, 如何更好地利用老芒麦资源, 改良土地盐碱化仍然是目前面临的瓶颈问题。外源人工合成激动素叶面喷施对作物光合特性、保护性酶及产量的影响报道较多, 但是在牧草老芒麦上鲜有报道, 基于此, 本研究以野生老芒麦为材料, 旨在探讨不同程度盐胁迫下, 叶面喷施激动素对老芒麦幼苗端粒酶活性、膜脂过氧化物、渗透调节物质的影响, 为土壤盐渍化地区的农业生产以及老芒麦人工草地改良提供理论依据和技术指导。
供试材料为2015年采自甘肃天祝县高寒草地的野生老芒麦种子。采样地海拔2950 m; 年均温-0.1 ℃, 最冷月(1月)平均气温为-18.3 ℃, 最热月(7月) 平均气温为12.7 ℃, 大于0 ℃年积温1380 ℃; 土壤pH为7.0~8.2; 有机质含量较高, 介于8%~16%, 速效氮0.017%, 全磷0.067%。水热同期; 年均蒸发量1592 mm, 没有绝对无霜期。
试验于2018年2月在甘肃农业大学草业生态系统国家重点实验室组培室开展。选取大小一致、饱满且无病虫害的老芒麦种子, 消毒处理后将其均匀撒在一次性口杯中, 每杯50粒, 覆盖一层浅沙, 每5杯放一个塑料培养盒, 每个处理重复3次, 共75杯, 移至组培室培养, 通风透光, 每天浇灌适量蒸馏水。出苗后间苗, 每杯保留生长一致、分布均匀的幼苗30株, 待苗长至2~3 cm, 浇Hoagland营养液[10], 每5 d换一次, 每次换营养液之前用蒸馏水冲洗以防止盐分积累。
设置5个氯化钠浓度梯度(0、50、100、150、200 mmol· L-1)和5个激动素浓度梯度(0、5、10、20、30 mg· L-1, 本研究分别用K1、K2、K3、K4、K5表示)对老芒麦幼苗进行处理, 待老芒麦幼苗长至三叶期时进行盐胁迫处理, 将不同浓度的NaCl溶液浇至培养盆中, 每盆0.5 L, 每72 h处理一次, 处理时间为168 h。168 h盐胁迫处理后用不同浓度的激动素溶液在老芒麦叶面喷施, 进行恢复处理, 每72 h喷施一次, 恢复时间为240 h。测定幼苗的端粒酶活性以及生理指标(叶绿素、游离脯氨酸、丙二醛、可溶性糖、可溶性蛋白含量)。
1.3.1 端粒酶活性测定 采用1994年Kim等[4]发明的末端序列重复扩增(TRAP)法, 将0.3 g的老芒麦叶片放入预先制冷的研钵中, 加入500 μ L的TRAP buffer提取液, 快速研磨成匀浆后, 倒入1.5 mL离心管中, 再加入500 μ L TRAP buffer, 在冰上放置30 min, 12000 r· min-1离心10 min, 将上清液转移到另一个新的离心管中, 加入12%(m/V)PEG 8000, 将离心管在-20 ℃冰箱放置沉淀60 min, 离心后弃去上清, 立即进行试验或者放在超低温冰箱中保存备用。采用50 μ L的反应体系:36 μ L固相RNA清除剂处理水, 5 μ L TRAP buffer, 1 μ L dNTP mixture, 1 μ L TS21前导链引物, 5 μ L端粒酶提取物, 26 ℃反应40 min, 94 ℃变性2 min, 然后再加入1.6 μ L ACX后随链引物和0.4 μ L DNA聚合酶进行PCR 扩增。PCR结束后对扩增产物进行12%非变性聚丙烯酰胺凝胶电泳检测, 电泳结束后, 将凝胶置于SYBR Green I 染色液中, 室温染色1 h, 用Quality one 软件分析试验结果。TRAP buffer配方为:50 mmol· L-1 Tris-HCl, pH 7.5, 5 mmol· L-1 MgCl2, 20 mmol· L-1EGTA (乙二醇双四乙酸), 0.1 mmol· L-1PMSF(苯甲基磺酰氟), 5 mmol· L-1DTT (二硫苏糖醇), 0.5%(m/V)CHAPS{3-[(3-胆酞胺丙基)-二乙胺]-丙磺酸}, 10%(m/V)甘油, 0.1 mmol· L-1benzamidine(苯甲脒), 2 mmol· L-1spermidine(亚精胺), 1.5%(m/V)PVP。
1.3.2 生理生化指标测定 叶绿素采用叶绿素含量测定仪对同一片叶的同一部位进行测定, 每个处理随机测定3片叶取平均值; 游离脯氨酸采用酸性茚三酮法测定[11]; 丙二醛采用硫代巴比妥酸(TBA)法测定[11]; 可溶性糖采用蒽酮法测定[11]; 可溶性蛋白含量采用考马斯亮蓝G-250染色法测定[11]。
采用SPSS 19.0 软件进行统计分析, Microsoft Excel 2016软件整理数据并绘图, 差异显著性定义为P< 0.05, 所有数据均为3个重复的平均值± 标准误(Mean± SE)。
| 表1 PCR 扩增过程中所用引物 Table 1 The primers used in PCR amplification |
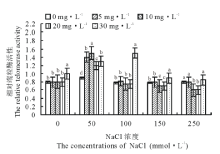
从图1可以看出, 随着NaCl浓度的增大, 端粒酶活性呈现出先上升后下降的趋势, 在50 mmol· L-1的NaCl盐胁迫处理下, 端粒酶活性达到最大值。当盐胁迫处理浓度大于50 mmol· L-1时, 端粒酶活性逐步减小。50 mmol· L-1的NaCl胁迫处理168 h, K2激动素恢复240 h, 端粒酶活性升高为对照组的1.56倍; 50 mmol· L-1的NaCl胁迫处理168 h, K3激动素恢复240 h, 端粒酶活性升高为对照的1.67倍。从整理来看, 随着激动素处理浓度的增大, 端粒酶活性逐渐上升, K5激动素恢复240 h以后, 相比对照组端粒酶活性明显升高。
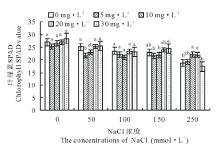
从图2可以看出, 相比对照组, 250 mmol· L-1的氯化钠盐胁迫处理下, 老芒麦叶绿素含量减少差异显著(P< 0.05), 相比对照组减少了44.2%。盐胁迫处理下, 激动素恢复以后, 抑制作用明显得到缓解, 而且随盐胁迫处理浓度的增大, 缓解效果更加明显。150 mmol· L-1盐胁迫下K2、K3激动素处理相比对照差异显著(P< 0.05), 分别减少了5.0%、4.0%, 而K4、K5激动素处理分别增长了5.0%、7.0%, 可见在同一浓度盐胁迫处理下, 随着激动素恢复浓度增大, 缓解效果较明显; 250 mmol· L-1盐胁迫下K3、K4激动素处理相比对照增加了18.7%、17.8%, K5激动素处理则减少8.2%。
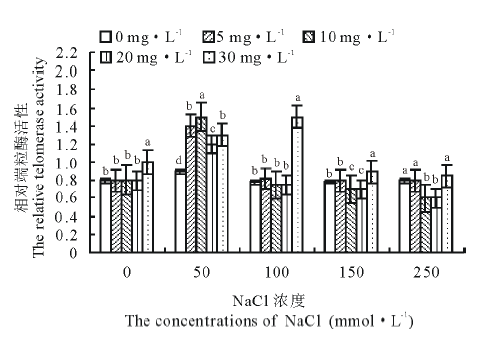 | 图1 不同浓度盐胁迫处理不同浓度激动素对老芒麦幼苗端粒酶活性的影响Fig.1 Effects of different concentrations of kinetin on telomerase activity in E. sibiricus seedlings under different salt stress |
 | 图2 不同浓度盐胁迫下不同浓度激动素恢复对老芒麦幼苗叶绿素含量的影响Fig.2 Effects of different concentrations of kinetin on chlorophyll content in E. sibiricus seedlings under different salt stress 不同字母表示处理间差异显著(P< 0.05), 下同。Different letters indicate significant difference among the treatments (P< 0.05).The same below. |
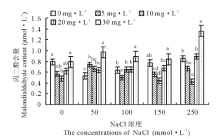
从图3可以看出, 50, 100, 150 mmol· L-1盐胁迫下老芒麦幼苗的丙二醛(malondialdehyde, MDA)含量相比CK依次减少了49.1%、23.4%、2.6%, 而250 mmol· L-1盐胁迫下却增加了7.6%, 可见适宜的盐浓度可以降低MDA的含量, 而盐浓度越高, 分泌的MDA越多。无盐胁迫下K3激动素处理下MDA相比对照显著减少64.9%(P< 0.05); 150 mmol· L-1盐胁迫下K2、K3激动素处理相比对照差异亦显著(P< 0.05), 分别减少了26.8%、42.2%; 250 mmol· L-1盐胁迫时, K3激动素处理相比对照MDA减少了50.0%。
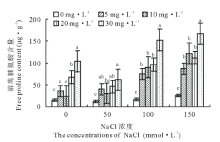
从图4可以看出, 老芒麦幼苗游离脯氨酸含量随着盐浓度的增大逐渐增加, 但各浓度处理间差异不显著(P> 0.05)。经激动素处理后发现, 0 mmol· L-1盐胁迫下K5激动素相比对照增加566.0%; 盐浓度为50 mmol· L-1时, K2、K4、K5激动素处理相比对照差异显著(P< 0.05), 分别增长了782.8%、271.4%、394.9%; 150 mmol· L-1盐胁迫下K4激动素处理增加了625.0%。表明盐胁迫下老芒麦幼苗体内的游离脯氨酸含量增加, 高盐浓度处理下其含量更高, 施加激动素处理后相比仅盐胁迫处理其含量进一步增加, 同一盐浓度处理下的高浓度激动素处理增加量显著(P< 0.05)。
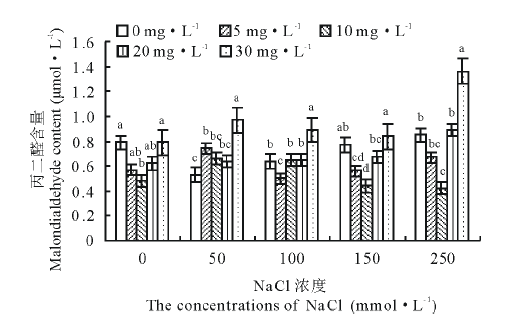 | 图3 不同浓度盐胁迫下不同浓度激动素处理对老芒麦幼苗丙二醛含量的影响Fig.3 Effect of different concentration of kinetin on the content of malondialdehyde (MDA) of E. sibiricus seedlings under different salt stress |
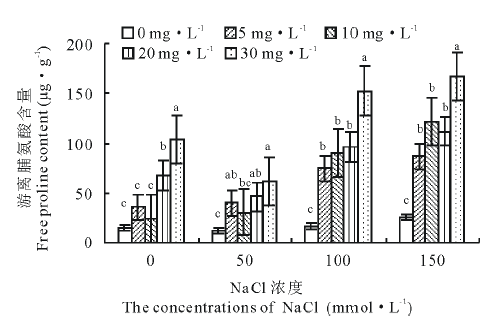 | 图4 不同浓度盐胁迫下不同浓度激动素处理对老芒麦幼苗游离脯氨酸含量的影响Fig.4 Effect of different concentration of kinetin on the content of free proline of E. sibiricus seedlings under different salt stress |
图5可以看出, 250 mmol· L-1盐浓度下可溶性糖含量显著增加(P< 0.05), 增长了101.5%, 与其他各浓度相比, 均达到了显著性差异。与盐胁迫处理相比, 激动素处理又更进一步促进了其含量的增加, 随着盐浓度的增大其促进效果先上升后下降, 以100 mmol· L-1盐浓度时激动素处理促进效果明显。同一盐浓度处理下, 随激动素浓度越高缓解效果越明显。说明适宜浓度的盐胁迫可以促进植物体内可溶性糖含量的积累, 激动素处理后促进作用更明显。具体看来, 外施激动素处理, 50 mmol· L-1盐胁迫下K2、K4激动素处理相比对照差异显著(P< 0.05), 分别增长了94.2%、77.6%; 100 mmol· L-1盐胁迫下K2、K4激动素处理差异显著(P< 0.05), 分别增长了77.1%、165.6%; 250 mmol· L-1盐胁迫下K4、K5激动素处理差异同样显著(P< 0.05), 分别增长了65.7%、66.0%。
可溶性蛋白含量在50、100、150 mmol· L-1盐胁迫处理下相比CK差异显著(P< 0.05), 分别增长了47.6%、52.4%、66.7%。表明盐浓度越高可溶性蛋白含量增加越明显。无盐胁迫处理下, K2、K3、K4、K5激动素处理相比对照差异显著(P< 0.05), 分别增长了74.6%、73.7%、68.4%、98.1%, 可见同一盐浓度处理下, 随激动素浓度的不断增大可溶性蛋白增加量先减少后增大; 50、100 mmol· L-1盐胁迫下各处理差异不显著; 250 mmol· L-1盐胁迫下K3、K5激动素处理分别增加了7.9%、12.7%。
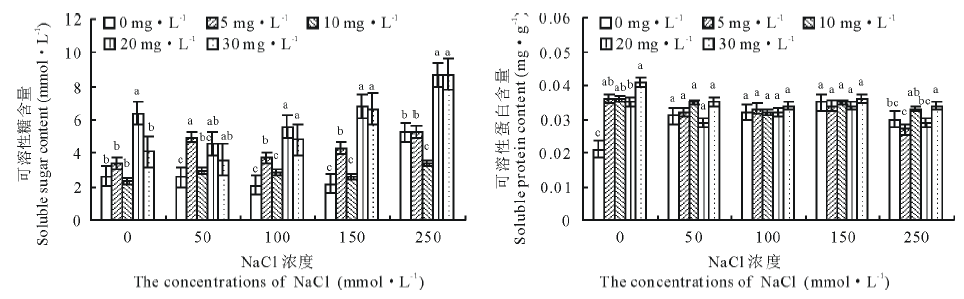 | 图5 盐胁迫下不同浓度激动素处理对老芒麦幼苗可溶性糖和可溶性蛋白含量的影响Fig.5 Effect of different concentrations of kinetin on soluble sugar content and soluble protein content in E. sibiricus seedlings under salt stress |
植物生长在盐胁迫环境下, 植物细胞内会产生活性氧(reactive oxygen species, ROS)[12], 高浓度的活性氧导致植物细胞内大分子物质如遗传物质、蛋白质以及细胞结构受到伤害[13, 14, 15], 研究结果表明, 盐胁迫处理严重影响了老芒麦幼苗的正常生长, 随着盐胁迫处理浓度增大, 阻碍作用越明显。100 mmol· L-1的氯化钠处理168 h以后, 渗透调节物质MDA含量明显升高, 游离脯氨酸大量外渗, 说明此时细胞膜已经受到破坏。氧化损伤诱导细胞程序性死亡(programmed cell death, PCD), 使细胞正常生长受到阻碍, 最终还会导致细胞直接死亡或程序性凋亡[16, 17]。研究结果表明, 100 mmol· L-1氯化钠胁迫处理168 h以后, 丙二醛(MDA)含量明显上升, 此时端粒酶活性明显增强, 说明细胞端粒酶活性的提高可能在一定程度上缓解盐胁迫导致的氧化损伤, 端粒酶在氧化损伤恢复和DNA修复方面发挥着重要的作用。端粒酶在维持端粒长度稳定性方面起到重要的作用。本研究中, 与低浓度氯化钠相比较而言, 当氯化钠浓度超过100 mmol· L-1时, 老芒麦端粒酶活性降低, 结合前人的研究成果以及本研究结果可以明显看出, 端粒系统抵御逆境胁迫及氧化损伤的作用具有一定的阈值, 过度的氧化损伤可能导致端粒酶结构成分发生变化, 最终导致端粒酶失去活性。植物生长状态是内外共同调控机制作用的结果, 通过本研究可以看出, 外源物质(如激动素)也会起到一定的缓解作用。
外源物质激动素能够缓解盐胁迫对植物的伤害, 从而促进植物的生长[18], 主要通过促进植物地上, 地下部分生物量的增加[18], 清除盐胁迫下植物体内自由基, 减少活性氧、膜脂过氧化物的积累, 刺激渗透调节物的大量产生, 增强植物在不良环境下的生存能力[19]。本研究发现在盐胁迫下, 老芒麦幼苗的各指标在激动素处理下均得到了不同程度的缓解, 如在0、150、250 mmol· L-1盐胁迫下, 10 mg· L-1激动素处理显著降低了MDA含量(P< 0.05), 分别减少了64.9%、42.2%、50.0%, 而盐胁迫下对大豆(Glycine max)幼苗[20]用同浓度的激动素处理发现MDA含量也降低了, 但是效果没有老芒麦的明显, 可见适宜浓度的激动素能够缓解盐胁迫对老芒麦造成的伤害, 进一步保护了其细胞膜系统; 对苦马豆(Sphaerophysa salsula)苗期的研究发现盐胁迫下6-BA处理游离脯氨酸含量相比盐处理增加了, 其主要机制是外源物质提高了脯氨酸合成酶的活性[21]。本研究盐胁迫下, 不同浓度的激动素处理老芒麦, 脯氨酸含量明显增加, 无盐胁迫下, 尤以30 mg· L-1显著(P< 0.05), 增加了566.7%, 50、150 mmol· L-1盐胁迫下激动素各浓度增加量亦显著(P< 0.05), 而苦马豆[21]、番茄(Solanum lycopersicum)[22]的处理效果不到30%, 表明激动素在老芒麦上的处理效果更明显; 可溶性蛋白含量在无盐胁迫下, 30 mg· L-1激动素处理增加了98.1%, 这与郑平生等[23]在草莓(Fragaria ananassa)试管苗上的研究基本一致。试验结果表明100 mmol· L-1盐胁迫下, 20 mg· L-1激动素处理可溶性糖含量增加显著(P< 0.05), 表明激动素处理更能促进盐胁迫下老芒麦体内可溶性糖含量的上升, 维持细胞渗透环境。整体评价发现老芒麦在盐胁迫下经激动素处理各项抗性指标较理想, 可以考虑在土壤盐渍化地区生产以及老芒麦人工草地改良上借鉴使用。
本研究从端粒酶活性和生理特性角度探讨激动素对老芒麦幼苗适应盐胁迫的影响。研究发现随着NaCl盐胁迫程度的增大, 渗透调节物质含量均上升, 经激动素处理以后, MDA含量下降, 叶绿素含量上升, 可溶性糖含量上升, 由此可以看出外源物质激动素对盐胁迫具有一定的缓解作用。适宜浓度的盐胁迫诱导了细胞端粒酶活性, 高浓度盐胁迫可能造成了老芒麦细胞的氧化损伤, 最终导致端粒酶活性下降。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|