


{kind=link}
{kind=link}
{kind=link}
蘑菇圈形成对高寒草甸群落植被结构及稳定性的作用
[李佳琪, 赵敏, 魏斌, 胡廷花, 于应文*  ]
]
]
|
|


作者简介:李佳琪(1996-),女,湖南嘉禾人,在读本科。E-mail:lijq14@lzu.edu.cn
蘑菇圈作为青藏高原高寒草甸一种常见生态学现象,也是草地群落植被更新重要场所。通过对青藏高原高寒草甸蘑菇圈区域群落植被特征和经济类群、生活型及演替顶极成分功能群生物量构成、Raunkiaer频度系数、演替度及Spearman秩相关特征进行分析,明晰蘑菇圈不同区域草地植被分异特征,解析蘑菇圈形成对草地植被变化和群落稳定性的作用。结果表明:1)蘑菇圈形成明显改变物种构成,圈上群落优势种为垂穗披碱草和赖草,而圈上和圈外群落皆无绝对优势种;2)蘑菇圈环的形成显著提高了圈上禾草、地下芽植物和降低种的生物量比例,降低其物种多样性;3)蘑菇圈圈上群落处于更高演替阶段,其植物种重要值与经济类群和演替顶极成分的生物量构成变化一致;4)圈上群落主要植物种对正负关联比为0.89,多数种对的种间联结松散,群落稳定性较低;圈内和圈外群落主要植物种对的正负关联比皆>1,多数种对间关联性较强,群落稳定性较强;且蘑菇圈群落中显著关联种对发生在亚优势种与伴生种或伴生种与伴生种之间。研究认为,蘑菇圈形成促进高寒草甸由莎草/杂类草群落向禾草/杂类草群落演替,但降低高寒草甸稳定性。
Fairy rings is a common ecological phenomenon in alpine meadows on the Qinghai-Tibet Plateau and is an important site for vegetation regeneration. In order to explore the effects of fairy ring formation on vegetation change and community stability in the Plateau’s alpine meadows, we studied the vegetation characteristics of different ring zones, functional group biomass composition by economic category, the life form and succession climax component, the Raunkiaer frequency coefficient, succession degree and Spearman rank correlations within the plant community surrounding the fairy ring zone. The results indicated that: 1) fairy ring formation significantly changed species composition. The dominant species on the fairy ring were Elymus nutans and Aneurolepidium dasystachys but there were no clearly dominant species either within and outside the rings; 2) fairy ring formation significantly increased the biomass ratios of grasses, geophytes and decreaser on the fairy ring, but reduced its species diversity; 3) the community on the fairy ring was at a higher successional stage, and the importance value of plant species accorded with their economic category and succession climax component type’s biomass composition; 4) on the fairy ring, the positive to negative associations ratio of the main species-pairs was 0.89, the interspecific association of most species-pairs was not significant and the community was in a lower stable state; while within and outside the fairy rings the ratios were both >1, the association between most species-pairs was stronger and the community was more stable. The respective associated species-pairs occurred mainly between sub-dominant species or companion species and the companion species of fairy ring communities. This research shows that fairy ring formation can encourage alpine meadow succession from sedges/forbs communities to grass/forbs communities, but that they also reduce the stability of alpine meadows.
青藏高原是世界上面积最大、海拔最高、植被类型最特殊的草地生态系统之一[1], 是我国重要天然牧场, 对我国草地畜牧业发展和生态安全维持具有重大意义。高寒草甸是青藏高原隆起和长期低温共同作用而形成的具有水平和垂直地带性的特殊植被[2], 为青藏高原广泛分布的典型植被类型, 约有0.7 hm2, 约占青藏高原草地面积的一半[3], 在青藏高原生物多样性保护、生态系统维持、区域经济发展等方面发挥重要作用[4]。
蘑菇圈斑块是指同种或异种蘑菇呈圈带状分布的一种生态学现象, 是由地下蘑菇菌丝体蔓延并向外辐射生长而形成的[5]; 其在青藏高原高寒草甸雨季普遍存在, 是植被更新的重要场所。国外学者较早对草地、林地蘑菇圈现象进行研究, 并按照蘑菇圈对植被的影响将其分为促进生长、抑制生长、无影响3类[6, 7, 8]。我国诸多学者亦对蘑菇圈区域草地植被和土壤特征进行较系统研究, 集中于典型草原和高寒草甸的生产力、物种构成、牧草养分[9, 10], 土壤理化性质和微生物特征[11, 12, 13, 14], 蘑菇圈的生态学特征[10]等。这些研究表明, 蘑菇圈形成促进圈上草地牧草生长, 提高牧草和土壤养分, 改变群落物种构成及土壤微生物结构特征, 此为蘑菇圈与草地植被互作的深入研究提供一定实践基础。
在群落尺度中, 蘑菇圈作为一种自然选择性干扰, 通过蘑菇菌丝体的生长代谢活动对所处植物-土壤生态系统产生影响, 往往表现出“ 去除物种” 而为适应干扰的先锋物种释放生态位[15], 蘑菇圈此种效应可改变草地群落种间竞争共存关系, 导致圈上草地群落结构发生改变, 进而影响群落稳定性和演替方向。在草地生态系统中, 植物物种种间关系和功能群特征常被用作群落稳定性和演替趋势的关键研究内容。种间关联是指各物种在生境中的相互关系, 常由群落生境的差异影响物种分布而形成, 当群落趋于演替顶极阶段时, 物种组成和群落结构将逐步趋于完善和稳定, 种间关系也将趋向正关联, 以求物种间的稳定共存[16]。因此, 种间关联不仅反映现阶段群落结构动态, 还体现群落稳定性和演替趋势, 是群落基本特征之一[17]; 而功能群构成特征作为研究物种随环境变化的基本单元, 简化了对物种研究的复杂程度, 用功能群构成特征来解释物种多样性格局和生态系统间功能关系更具说服力[18]。我国学者依据植物光合作用途径、物种生活型、物种生态类型、经济类群等对草地植物功能群进行划分, 并通过功能群构成及其相互作用关系, 对物种多样性[19]、群落生物量[20]、生态系统稳定性机制[21]等进行揭示。但以往蘑菇圈群落植物功能群研究, 多分析蘑菇圈植被的经济类群, 认为禾草是蘑菇圈群落中优势功能群, 蘑菇圈形成对草地植物生长具明显施肥效应, 其效应因植物种不同而变化[12, 14, 22]。然而, 因蘑菇圈植被无固定优势种, 这对蘑菇圈区域植被结构动态演替机制的深入研究造成极大不便。且以往草地蘑菇圈植被分析, 有关群落植被构成与稳定性的整合研究亦相对缺乏。基于此, 本研究从种间关系和多功能群角度, 探讨高寒草甸蘑菇圈区域群落植被构成和稳定性特征及其关系, 明晰蘑菇圈不同区域草地植被分异规律, 解析蘑菇圈形成对草地植被变化和群落稳定性的作用, 为小尺度草地植被异质性形成机制的深入揭示提供一定理论依据。
研究区位于青藏高原东北缘的甘肃省天祝县抓喜秀龙乡境内的甘肃农业大学天祝高山草原试验站附近的高寒草甸冬季牧场上, 地理坐标为E 102° 40'-102° 47', N 37° 11'-37° 14', 海拔2960 m。年均温-0.1 ℃, 1月气温最低为-18.3 ℃, 7月气温最高为12.7 ℃, > 0 ℃年积温1380 ℃; 多年平均降水量416 mm, 无绝对无霜期, 仅分冷热两季, 土壤类型为亚高山草甸土。草地植物种主要有垂穗披碱草(Elymus nutans)、矮嵩草(Kobresia humilis)、线叶嵩草(Kobresia capillifolia)、洽草(Koeleria cristata)、西北针茅(Stipa krylovii)、珠芽蓼(Polygonum viviparum)、扁蓿豆(Melilotoids ruthenica var. inschanica)和球花蒿(Artemisia smithii)等。
于2016年7月末, 在草地利用率约90%且蘑菇圈频繁出现的高寒草甸冬季牧场上, 随机设置3个面积0.3~0.5 hm2的样地。
1.3.1 群落特征 2016年7月末-8月中下旬, 在已设置的各样地内选择5个呈弧形草带的直径约为5 m的典型蘑菇圈斑块, 圈上部分宽70~90 cm, 各蘑菇圈斑块之间距离20 m以上, 3块样地共选择蘑菇圈斑块15个。由于蘑菇圈植被呈现明显圈带现象, 圈内和圈外植被较圈上低矮, 故将各蘑菇圈斑块植被划分为圈内、圈上和圈外3个部分。同期, 在各蘑菇圈斑块的圈内、圈上和圈外各设置2个0.5 m× 0.5 m的样方, 其中, 圈外样方应设置在相应蘑菇圈5 m外, 且植被分布较为均匀并未被真菌感染的草地上, 每个蘑菇圈斑块共设置配对样方6个。测定各样方内草层及所有植物种的高度和盖度, 然后齐地刈割收获地上生物量, 将其按绿色物质(活物质)和死物质分开, 再将绿色物质按不同植物种分开, 于65 ℃下烘干称重, 测定分种生物量。
以样方植物种的高度和盖度数据为基础, 按公式IV=(RH+RC)/2计算群落植物种重要值。式中, IV为植物种重要值(importance value), RH和RC分别为植物种的相对高度(relative height, 样方内某植物种的高度/样方内所有植物种的高度之和)和相对盖度(relative coverage, 样方内某植物种的盖度/样方内所有植物种的盖度之和)。并按公式β w=(S/ma)-1, 计算物种β 多样性指数(β w)。式中:S为群落中物种数; ma为各样方或样本的平均物种数[23]。
1.3.2 功能群生物量构成 按植物种经济类群、生活型和演替顶极成分3种方法, 分别统计不同功能群植物种生物量比例。其中, 将样方生物量按经济类群分为禾草、莎草、豆科、其他和不可食植物5类; 按生活型分为1年生植物(含越年生植物)(therophyte, TP)、地面芽植物(含地上芽)(hemicryptophyte, HP)和地下芽(geophyte, GP)植物3类; 按演替顶极成分分为降低种(decreaser), 增加种(increaser)和入侵种(invader)3类, 降低种是草地上价值最高的优良草种, 随放牧加重, 相对产量减少, 增加种是指草地上产量低、适口性差的植物种, 随放牧加重, 相对产量增加, 入侵种一般是在放牧加重或原始植被生活力下降后出现的饲用价值很低的杂草[24]。
1.3.3 草地Raunkiaer频度系数和演替度 Raunkiaer把频度系数划分为5个等级, A(级)=1%~20%, B(级)=21%~40%, C(级)=41%~60%, D(级)=61%~80%, E(级)=81%~100%。频度系数R=(n/N)× 100%, 式中:n为某一个种在全部取样中出现的次数; N为全部取样数[24]。Raunkiaer频度直方图越接近反J型, 群落越稳定; 草地演替度(degree of succession, DS)越大, 草地群落越趋于顶极阶段。
式中:e为植物种的寿命; d为植物种优势度(重要值); N为各重复样地内群落总植物种数; μ 为植被覆盖率。
1.3.4 植物种间Spearman秩相关 以植物种重要值数据为依据, 采用SPSS 16.0中Correlate中的Bivariate模块, 计算蘑菇圈圈内、圈上、圈外主要植物种间Spearman秩相关系数及显著性检验。以进一步分析蘑菇圈不同位置群落相对稳定性及群落内主要植物种间关系。
用Excel进行数据处理及制图, SPSS16.0软件的One-Way ANOVA对蘑菇圈圈内、圈上、圈外群落植被构成及功能群生物量特征进行方差分析(F-检验)和显著性检验, 并将数据表示为均值(mean)± 标准误(standard error of mean, Sem)。
蘑菇圈主要植物种重要值显示, 圈内、圈上和圈外群落植物种构成差异较大(表1)。圈上群落以垂穗披碱草和赖草为主要优势种, 其重要值为0.201和0.189; 早熟禾为亚优势种, 其重要值为0.100; 洽草、线叶嵩草、球花蒿、秦艽等为主要伴生种。圈内与圈外群落均无绝对优势种, 圈内主要植物种有垂穗披碱草、早熟禾、洽草、球花蒿等, 圈外主要植物种有垂穗披碱草、线叶嵩草、早熟禾、矮嵩草等; 且圈内和圈外的垂穗披碱草和赖草重要值均比圈上的低。说明, 蘑菇圈形成可明显改变群落物种构成。
| 表1 蘑菇圈群落主要植物种重要值 Table 1 The importance values (IV) of main species of fairy ring communities (IV≥ 0.02) |
蘑菇圈形成对群落草层高度、总生物量、物种数、β 多样性指数β w均具显著或极显著影响(表2)。草层高度和总生物量均为圈内=圈外< 圈上(P< 0.05), 且圈上草层高度和生物量分别是圈外和圈内的3.2~3.8倍和1.7~1.8倍; 群落物种数为圈内=圈外> 圈上(P< 0.05); 而其β w为圈内> 圈外(P< 0.05), 且均与圈上相近(P> 0.05)。说明, 蘑菇圈环的形成利于草地植物生长, 降低物种多样性。
| 表2 蘑菇圈植物群落特征 Table 2 Plant community characterisitic of fairy ring communities |
蘑菇圈经济类群生物量构成显示, 禾草比例为圈上> 圈内=圈外(P< 0.05), 前者为后者的3.1和4.0倍; 莎草和其他科比例为圈上< 圈内=圈外(P< 0.05), 且后者为前者的7.9~11.1倍和6.4~10.9倍; 不可食植物比例为圈外> 圈内> 圈上(P< 0.05); 而豆科植物比例在蘑菇圈不同位置相近(P> 0.05)(图1A)。圈上群落生物量构成以禾草为绝对优势(89.7%), 不可食、莎草和其他科植物仅占2.4%~4.2%; 圈内群落生物量构成, 禾草、莎草、其他科和不可食植物各占20.9%~28.6%; 圈外群落生物量构成, 莎草(33.0%)和不可食植物(28.0%)较多, 禾草居中(22.4%), 其他科植物较少(15.2%); 蘑菇圈不同位置豆科植物极少(图1A)。说明, 蘑菇圈形成促进高寒草甸由莎草/杂类草群落向禾草/杂类草群落转化。
蘑菇圈植物生活型谱生物量构成显示, 地面芽植物(HP)为圈外> 圈内=圈上(P< 0.05), 地下芽植物(GP)为圈上> 圈内=圈外(P< 0.05), 1年生植物(TP)在蘑菇圈3个位置之间相近(P> 0.05)(图1B)。群落生物量构成以HP为主, 圈内、圈上和圈外HP比例分别为62.2%、57.3%和68.9%, GP比例分别为25.2%、33.0%和22.6%, TP仅约为10%(图1B)。表明, 蘑菇圈形成利于圈上地下芽植物生长。
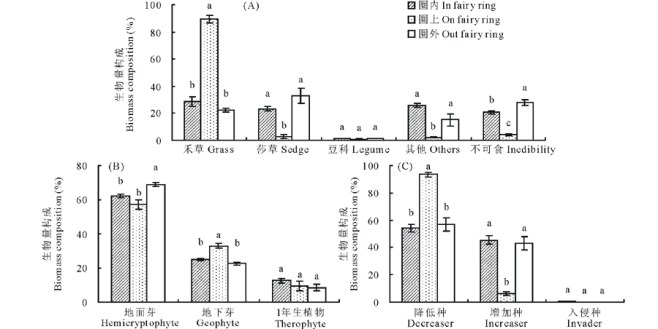 | 图1 蘑菇圈群落功能群生物量构成 (A)经济类群Economic category; (B)生活型Life form; (C)演替顶极成分Succession climax component. 不同小写字母表示在P< 0.05水平下差异显著。The different small letters mean the significant differences at P< 0.05.Fig.1 Biomass composition of different functional groups of fairy ring communities |
蘑菇圈演替顶极成分生物量构成显示, 降低种生物量比例为圈上> 圈内=圈外(P< 0.05), 前者是后者的1.6~1.7倍; 而其增加种生物量比例则为圈上﹤圈内=圈外(P< 0.05), 后者为前者的7.2~7.6倍; 3个位置入侵种生物量比例均< 1%(图1C)。说明, 蘑菇圈形成加速圈上群落演替进程, 使其趋于演替顶极阶段。
群落植物种Raunkiaer标准频度直方图结果显示, 蘑菇圈圈内、圈上和圈外群落各级植物频度皆符合Raunkiaer频度定律, 即A级> B级> C级≥ 或≤ D级< E级。圈内、圈上、圈外的A级植物频度系数分别为54.1%、48.3%和42.1%, E级植物频度系数分别为24.3%、24.1%和21.1%, 其A级和E级植物比值分别为2.2、2.0和2.0(图2); 且其频度图均呈反J型。说明, 蘑菇圈群落内部植物物种分布较均匀, 群落整体处于相对稳定状态。蘑菇圈群落演替度值结果为, 圈上(47.27± 5.40)> 圈内(30.21± 1.70)=圈外(30.73± 1.03)(P< 0.05)。说明, 圈上群落处于更高演替阶段, 与其植物种重要值及经济类群和演替顶极成分功能生物量构成结果一致。
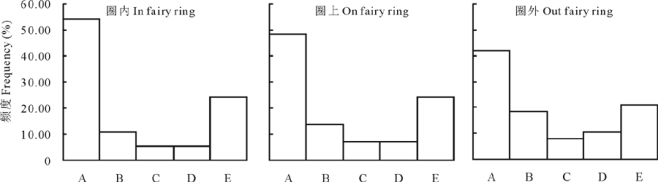 | 图2 蘑菇圈群落Raunkiaer标准频度直方图Fig.2 Standard frequency histogram of fairy ring communities A: 1%~20%; B: 21%~40%; C: 41%~60%; D: 61%~80%; E: 81%~100%. |
Spearman秩相关分析显示, 在蘑菇圈圈内、圈上和圈外群落中, 正相关种对数分别占各自总对数的53.3%、47.0%和61.7%, 负相关种对数分别占各自总对数的46.7%、53.0%和38.3%, 正负关联比分别为1.14、0.89和1.61; 其中, 显著或极显著正关联种对分别占3.3%、9.1%、2.5%, 显著或极显著负关联种对分别占4.2%、0、1.7%(图3)。说明, 蘑菇圈圈上多数植物种对的种间联结松散, 群落稳定性较低; 圈内和圈外群落多数植物种对的种间关联性较强, 群落内部各物种相互依赖, 群落稳定性相对较强。圈内群落中, 显著正相关种对为球花蒿和翻白叶委陵菜, 秦艽和二裂委陵菜, 线叶嵩草和矮嵩草; 显著负相关种对为球花蒿和蒲公英, 秦艽和唐松草, 线叶嵩草和紫花针茅、甘肃棘豆(图3)。圈上群落仅存在显著正相关种对, 为扁蓿豆和矮嵩草, 早熟禾和秦艽、洽草, 洽草和秦艽、线叶嵩草。圈外群落中, 显著正相关种对为球花蒿和早熟禾、秦艽; 显著负相关种对为早熟禾和甘肃棘豆, 矮嵩草和紫花针茅。说明, 蘑菇圈群落中显著关联种对发生在亚优势种与伴生种或伴生种与伴生种之间。
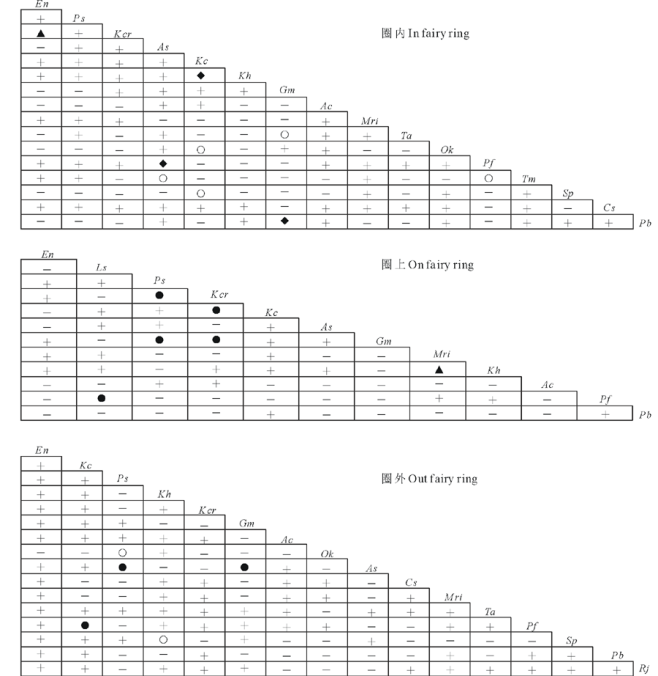 | 图3 蘑菇圈群落不同位置主要植物种对的Spearman秩相关分析Fig.3 Spearman rank correlation analysis of main species in different fairy ring communities正相关Positive correlation:▲, P< 0.001; ◆, 0.001< P≤ 0.01; ●, 0.01< P≤ 0.05; +, P> 0.05.负相关Negative correlation:△ , P< 0.001; ◇, 0.001< P≤ 0.01; ○, 0.01< P≤ 0.05; -, P> 0.05. |
一般认为, 蘑菇圈形成利于草地群落植物生长和生产力提高[5, 14, 25], 本研究结果与此一致。这是由于蘑菇圈菌丝体向外辐射生长过程中, 地下菌丝逐渐消亡降解, 释放大量速效氮磷养分, 降低物种间资源竞争, 为植被生长提供物质基础[22]。同时, 蘑菇菌丝分泌代谢产物[25], 刺激圈上纤维素分解菌等细菌生长, 使有机物质迅速转化为糖类、醇类及各种有机酸, 进而促进圈上植被生长[14]。另外, 蘑菇圈圈上优势种为禾草, 其强大须根系和蘑菇菌丝交织作用, 增强草地土壤团粒结构[10], 此亦可促进圈上植物生长。但Edwards[8]认为, 蘑菇菌丝具有杀草作用, 使圈上牧草减产; 其原因可能是蘑菇菌丝生长充满土壤地下空间, 妨碍土壤表面水分自由扩散, 使植被因缺水而生长受阻所致, 也可能是菌丝产生氰化氢等毒素毒害植物根尖而降低圈上植物生长所致。
本研究蘑菇圈形成显著降低群落物种多样性。这一方面是由于圈上禾草对蘑菇圈的施肥效应敏感, 使蘑菇圈上绝对优势种垂穗披碱草和赖草的生长速率和高度高于其他物种, 由此刺激蘑菇圈植物群落渐向具有绝对优势种的方向发展, 进而使蘑菇圈上群落的异质性比圈内和圈外更高, 由此降低群落物种多样性。另一方面是由于蘑菇圈上高大优势植物对其他物种产生遮蔽作用, 降低群落下层植物透光率及物种存活率, 使下层不耐荫又低矮的物种被竞争优势物种排除[26], 由此导致物种多样性下降。这与赵吉等[9]对典型草原蒙古白蘑的研究结果一致, 但与雷茜等[22]、王文颖等[5]和王启兰等[14]对高寒嵩草草甸黄绿蜜环菌的研究结果相反。造成蘑菇圈形成对草地群落物种多样性影响不同结果的原因, 可能与蘑菇圈菌丝体种类、生境和蘑菇圈形成后的研究时期等差异有关[9, 14, 25]。
植物功能群按其性状及数量进行分类, 集中反映植物对生境的适应性及生态系统的功能特征[27]。本研究蘑菇圈形成, 提高适口性好的降低种如禾草等的生物量比例, 降低适口性差的增加种等的生物量比例, 使圈上群落由莎草/杂类草群落向趋于演替顶极阶段的禾草/杂类草群落转化。这是由于蘑菇圈圈上各经济类群中, 禾草多而莎草少, 禾草和莎草对蘑菇圈形成的响应较显著, 通过竞争释放使圈上高大禾草生物量增大, 以补偿莎草和其他科植物减少的生物量; 同时, 本研究物种多样性降低亦会释放更为多样化的生态位空间, 存留物种也会展现更广的适应性[28], 进而使草地群落盖度增高, 由此使禾草的遮阴作用和竞争优势更明显; 二者均使蘑菇圈圈上群落具有向禾草逐步演化的趋势。此与王文颖等[5]、赵吉等[9]和陈立红等[25]的报道, 蘑菇圈形成显著提高高寒草甸和典型草原禾草生物量和比例及草地趋于禾草方向演替的整体结果类似。
群落稳定性依赖于物种对生境变化的敏感性和种间关系对环境干扰的放大效应[21]。本研究Raunkiaer标准频度直方图结果为现阶段蘑菇圈圈上群落处于相对稳定状态, 但Spearman秩相关结果又显示, 蘑菇圈圈上群落稳定性较圈内和圈外弱, 群落整体趋向于微负关联; 这是由于圈上植物以活性的对施肥和干扰极敏感[17]的禾草为主, 且蘑菇圈通常出现在圈上植被的外围, 出现初期, 圈上群落因上一年蘑菇菌丝的分解可获取足量养分以供植被生长, 但随蘑菇菌丝向外扩展圈上群落因蘑菇菌丝分解而对土壤的施肥效应逐渐消除, 由此预测, 当年圈上群落将会在较短时间内演替成圈内群落并脱离蘑菇菌丝体的干扰, 而当年圈外群落则很快被向营养的蘑菇菌丝侵染发展成圈上群落而受到干扰[8]。所以, 圈上群落长期处于蘑菇圈干扰范围内, 且圈上群落的位置将随蘑菇圈的外延一直处于变化之中, 进而使圈上群落处于不稳定状态。
此外, 虽然本研究对高寒草甸蘑菇圈群落植被构成和稳定性特征进行了分析, 明确了蘑菇圈区域草地植被变化特征及演替方向, 但蘑菇圈形成与植被演变的互作过程及机制仍不明晰, 尚需将来进一步深入研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|