{kind=link}
{kind=link}
冬小麦套种草木樨土壤中根瘤菌分布与土壤理化性质的相关性分析
[赵涛1 , 马春晖2 , 王栋1 , 景永元3 , 席琳乔1, *  ]
]
]
|
|


作者简介:赵涛(1991-),男,甘肃武威人,在读硕士。E-mail: zhaotaocn@163.com
对新疆阿克苏地区冬小麦套种草木樨地土壤中根瘤菌数量动态规律与土壤理化性质的相关性进行了研究。采用平板稀释法对根瘤菌进行计数,测定土壤理化性质包括水分、pH、容重、有机质、全氮、碱解氮、速效磷和速效钾。结果表明:1)从冬小麦返青也是草木樨播种时期(4月)到刈割期(9月),根瘤菌数量分布为10~20 cm>0~10 cm>20~40 cm土层,0~20 cm土层根瘤菌数量7月达到最大值,为6.89 lg cfu·g-1,20~40 cm土层根瘤菌数量8月达到最大值,为6.46 lg cfu·g-1;非共生期根瘤菌比共生期增加7.33%~10.63%。2)4-9月土壤pH下降0.23%~1.40%,全盐下降3.45%~37.69%;0~40 cm土层4-8月有机质、全氮、碱解氮、有效磷呈下降趋势,但在9月草木樨刈割时较8月有所增加,其中0~40 cm土层的有机质增加7.65%~26.98%,全氮增加16.67%~44.12%;0~20 cm土层碱解氮增加20.27%~160.68%;10~20 cm土层有效磷增加167.74%;0~20 cm土层的有效钾含量提高10%。3)根瘤菌数量与土壤理化性质的线性回归分析,0~40 cm土层全盐与根瘤菌数量相关性最高( R=-0.81, P<0.05)。此外,0~20 cm土层水分还与根瘤菌数量相关性高(-0.81≤ R≤-0.73, P<0.05),20~40 cm土层全氮的相关性高( R=-0.88, P<0.05);通径分析中,土壤全氮、全盐、有机质与根瘤菌数量相关性最高,其中与有机质呈正相关( P5=0.56),与全氮、全盐呈负相关( P6=-0.50, P4=-0.64),有机质是促进根瘤菌的主要因子,全氮、全盐是制约主要因子。因此,在栽培过程中应该增加有机肥,适当减少氮肥施量,降低土壤盐分,促进根瘤菌增殖,提高固氮量。
The correlation between rhizobia distribution and the physical/chemical properties in soil of winter wheat intercropped with sweet clover in the Aksu region of Xinjiang was determined. Rhizobia numbers were counted by dilution plate method. Soil physical and chemical properties including pH, soil bulk density, total salt, organic matter, total nitrogen, alkali-hydrolyzale nitrogen, available phosphorus and available potassium were also measured. The results showed that the number of rhizobia was highest at 10-20 cm soil depth compared with 0-10 cm and 20-40 cm from sowing to cutting of sweet clover. The number of rhizobia was highest at 0-20 cm depth (6.89 lg cfu·g-1) in July and at 20-40 cm soil depth in August (6.46 lg cfu·g-1). Soil pH and total salt decreased 0.23%-1.40% and 3.45%-37.69% between September and April, respectively. At 0-40 cm soil depth, organic matter, total nitrogen, available nitrogen and available phosphorus decreased from April to August. However, compared with September (sweet clover harvest) organic matter and total nitrogen in August was increased 7.65%-26.98% and 16.67%-44.12%, respectively. Alkali-hydrolyzable nitrogen at 0-20 cm was increased by 20.27%-160.68%, available phosphorus by 167.74% at 10-20 cm and available potassium by 10% at 0-20 cm soil depth. The correlation between total salt and the number of rhizobia at 0-40 cm soil depth was strong and negative ( r=-0.81, P<0.05. The correlations between moisture content and rhizobia numbers at 0-20 cm soil depth were also strong and negative (-0.81≤ r≤-0.73, P<0.05). There was also a strong negative correlation between total nitrogen content and rhizobia numbers at 20-40 cm soil depth( r=-0.88, P<0.05). Path coefficients for total salt, organic matter and total nitrogen were generally strongly correlated with rhizobia numbers, and organic matter was positively correlated ( P5=0.56) but total nitrogen and total salt negatively correlated ( P6=-0.50, P4=-0.64). Organic matter had the greatest positive influence on rhizobia, whereas total nitrogen and total salt reduced rhizobia.
冬小麦(Triticum aestivum)套种草木樨(Melilotus suaveolens)种植模式, 在1955年就有相关研究, 在生产中得到了广泛应用, 尤其在“ 一季有余, 两季不足” 的地区[1]。豆/禾套种模式在时间和空间上高效利用土地光热等资源, 冬小麦套种草木樨还充分发挥出草木樨抗性优势和种间互助优势, 达到了“ 藏粮于草” 、“ 粮草丰收” 的目的[2, 3]。近几年来, 新疆大力发展林果业和棉花, 连作重茬严重, 土壤贫瘠, 饲草不足, 影响畜牧业发展, 套种草木樨解决了当地饲草料不足的问题, 提高土壤肥力, 实现了增产增收[4, 5, 6]。豆科植物中的根瘤菌能将空气中氮素转化为植物所需的氮素, 减少氮肥用量, 降低耕地污染风险, 减少对土壤生态环境的破坏及土壤养分失衡的危害, 有利于农业的可持续发展[7, 8, 9, 10, 11]。在果园小麦、向日葵(Helianthus annuus)、玉米(Zea mays)、棉花(Gossypium spp.)等与草木樨套种, 均能提高土壤有机质及氮素含量, 实现增产[12, 13, 14, 15]。豆科植物接种根瘤菌, 能显著改善土壤理化性质, 增加土壤肥力, 提升产量和改善品质[16, 17, 18, 19]。另外, 通过增施肥料或改善土壤物理性状也可提升根瘤菌数量及固氮效率, 优质土壤更利于根瘤菌的增殖和固氮[20, 21, 22, 23]。在冬小麦套种草木樨研究中, 套种处理能提高更深层次养分的利用率, 减少浅土层养分损失, 抑制盐碱化, 改良土壤, 且草木樨根系分泌物能促进微生物增殖, 提升微生物数量[2, 24]。套种草木樨可以分别提升土壤有机质, 全氮, 碱解氮12.1%~84.8%、32%~41%、12.2%。冬小麦套种草木樨可实现小麦每hm2增产16.0%~37.5%, 下一季玉米平均增产8.7%以上, 草木樨干草4500~11250 kg, 还能大幅度提高土壤保水保肥的能力[1, 2, 25, 26, 27]。目前, 国内对草木樨根瘤菌动态分布与土壤理化性质的相关报道还较少, 本研究测定4-9月冬小麦套种草木樨样地0~40 cm土层中根瘤菌的分布情况, 并测定该土层土壤理化性质, 分析其动态规律及二者间的相关性, 为冬小麦套种草木樨模式在时间、空间上合理施肥和接种根瘤菌提供理论基础, 为农业节能减排提供数据支撑。
试验田为冬小麦套种草木樨种植模式, 该种植模式是在上一年10月播种冬小麦(行距为15 cm), 来年3月麦田化冻施肥时将草木樨与氮肥一并条播进麦田中(草木樨之间行距为15 cm, 冬小麦与草木樨之间行距为7.5 cm), 在4-6月形成冬小麦与草木樨共生期, 在7月刈割冬小麦后, 7-9月草木樨迅速生长, 形成冬小麦与草木樨非共生期。试验田位于新疆阿克苏地区拜城县察尔其镇, 年均日照2789 h, 年均降水量171 mm, 年均无霜期133~163 d。属于温带大陆性干旱气候, 冬寒夏凉, 光照充足, 四季分明。试验地地理位置N 41° 37'36″, E 81° 23'51″, 海拔1410 m。
采用“ W” 5点取样法, 在冬小麦返青期(4月)到草木樨收获期(9月)采集冬小麦与草木樨行间的土样, 每月一次, 分3个土层0~10 cm、10~20 cm和20~40 cm, 装入无菌自封袋贴好标签, 带回实验室分析。将带回实验室的土样, 立即进行根瘤菌计数、容重、水分测定。测定完成后, 将土样经自然风干, 各样品分别过0.25、1 mm孔径筛网后装袋贴好标签, 测定样品土壤理化性质。
1.3.1 根瘤菌概况计数 使用改良的YMA固体鉴别培养基(BTB试验)、平板稀释梯度法测定[28, 29]:取10 g土样, 加入50 mL灭菌蒸馏水振荡混匀, 定容至100 mL移至三角瓶中, 150 r· min-1, 28 ℃, 振荡30 min, 制成分散均匀的土壤悬浊液, 静置15~20 min; 稀释至10-3, 10-4, 10-5, 取50 μ L悬液均匀涂布至YMA平板, 28 ℃静置培养3~5 d, 记录BTB试验产酸产碱的菌落数, 7~10 d如有新菌落产生则再记录一次数据, 5次重复。
1.3.2 土壤理化性质的测定 物理性质测定:水分用烘干法(105 ℃)测定; pH用电位法测定(液土比为2.5∶ 1); 容重用环刀法测定。
化学性质测定[30]:土壤水溶性盐用质量法测定; 土壤有机质用重铬酸钾-硫酸消化法测定; 全氮用重铬酸钾-硫酸消化-蒸馏法测定; 碱解氮用碱解扩散法测定; 有效磷用碳酸氢钠浸提-钼锑抗比色法测定; 有效钾用乙酸铵浸提-火焰光度法测定。
采用Microsoft Excel 2007进行基本数据统计分析和绘图; 采用SPSS 17.0软件One-way ANOVA进行方差分析, Duncan法进行显著性检验; 对土壤理化性质与根瘤菌数量的相关性进行线性回归分析; 使用DPS v7.55软件的通径系数分析法综合分析土壤理化性质(X1为水分、X2为容重、X3为pH、X4为全盐、X5为有机质、X6为全氮、X7为碱解氮、X8为有效磷、X9为有效钾)与根瘤菌数量的关系; 根瘤菌计数结果取lg值; 冬小麦与草木樨共生期土壤根瘤菌数量取4-6月的均值, 非共生期土壤根瘤菌数量取7-9月的均值; 所有数值以平均值± 标准误表示。
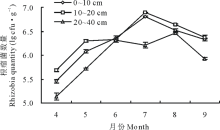
冬小麦套种草木樨根瘤菌数量动态变化结果(图1)表明, 0~20 cm土层中7月根瘤菌数量达到最大值, 为6.89 lg cfu· g-1(P< 0.05); 20~40 cm土层中6-8月根瘤菌数量较多, 其中8月达到最大值, 为6.46 lg cfu· g-1(P< 0.05), 0~10 cm土层中6、9月间差异不显著, 10~20 cm土层中5、6月间差异不显著, 其余各剖面土层在时间上差异均显著(P< 0.05); 整体上0~20 cm土层中的多于20~40 cm土层, 其中10~20 cm土层最多, 6-8月各土层变化幅度较小。
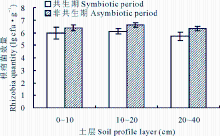
冬小麦套种草木樨4-6月为共生期, 7-9月为非共生期, 0~40 cm土壤根瘤菌数量非共生时期均高于共生期(图2), 0~10 cm、10~20 cm、20~40 cm土层分别增长了7.33%、8.68%、10.63%, 其中20~40 cm土层增长最多。
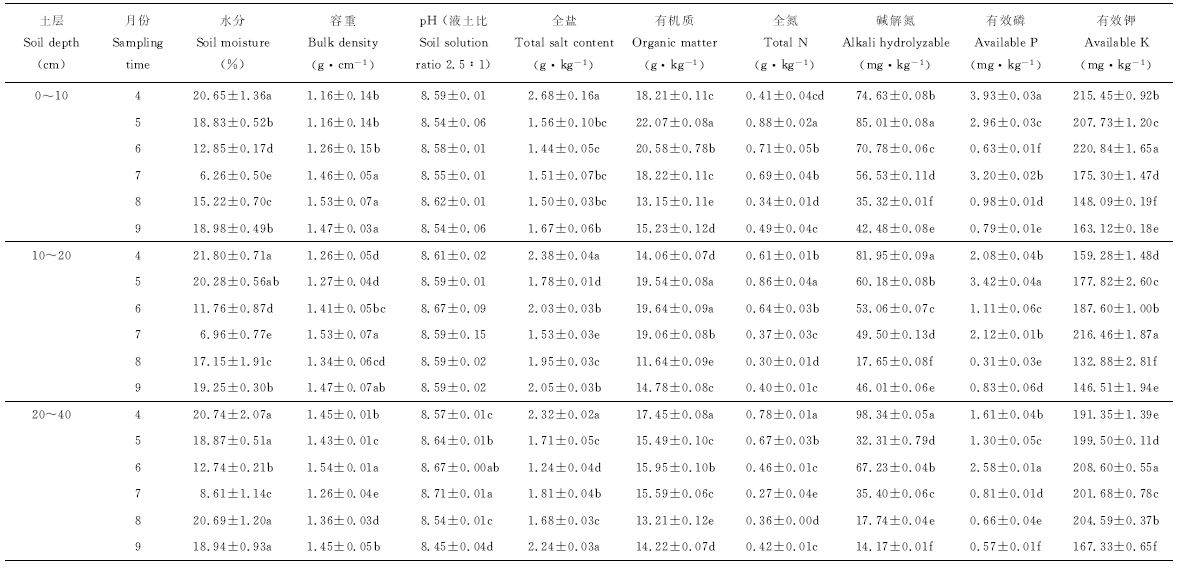
从草木樨播种到收获时期0~40 cm土层土壤含水量呈“ V” 型变化, 4月土壤水分最高(P< 0.05), 7月最低, 正是冬小麦的收获期(表1)。土壤容重在0~40 cm土层从整体上呈上升趋势; 共生时期20~40 cm> 10~20 cm> 0~10 cm; 在非共生期高于共生期, 其中20~40 cm土层较0~20 cm土层下降; 9月较8月0-10 cm土层下降4.36%。土壤pH 为 8.54~8.67, 非共生期较共生期下降了0.70%; 9月较4月下降0.23%~1.40%。土壤全盐在0~40 cm土层呈下降趋势, 0~20 cm非共生期较共生期下降了10.66%~17.61%; 9月较4月降低3.45%~37.69%。有机质在非共生期较共生期下降12.01%~23.43%, 全氮下降24.00%~49.29%; 0~20 cm土层中有机质、全氮含量在5月最高(P< 0.05); 20~40 cm均在4月最高(P< 0.05); 在草木樨收获时, 9月较8月有机质增加7.65%~26.98%、全氮增加16.67%~44.12%, 得到一定的补充。碱解氮含量0~40 cm土层整体呈下降趋势; 4-8月持续下降, 8月较4月下降52.67%~81.96%; 但在9月草木樨收获时0~20 cm土层较8月增加了20.27%~160.68%; 有效磷含量整体呈下降趋势, 9月草木樨收获时较4月下降8.02%~24.29%。6月0~40 cm土层共生期土壤有效钾含量较4月提高6.30%~17.78%, 其中20~40 cm> 10~20 cm> 0~10 cm, 且共生期较非共生期高4.31%~24.46%; 非共生期0~20 cm土层在草木樨收获时有效钾含量提高10%, 20~40 cm土层降低18.21%。
| 表1 冬小麦套种草木樨土壤理化性质 Table 1 Soil physical and chemical properties of winter wheat interplanting sweet clover |
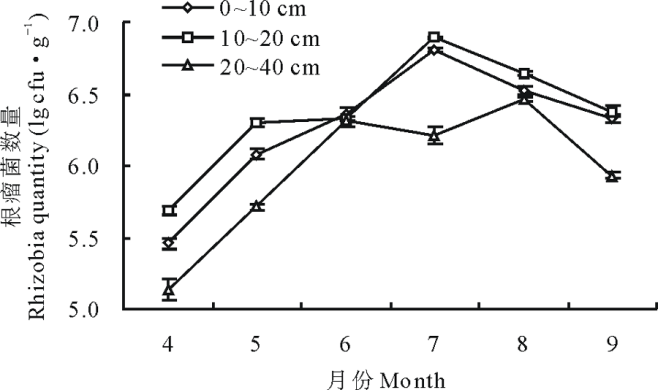 | 图1 冬小麦套种草木樨0~40 cm土壤中根瘤菌数量的变化Fig.1 Rhizobia quantity changes of winter wheat interplanting sweet clover in the 0-40 cm soil |
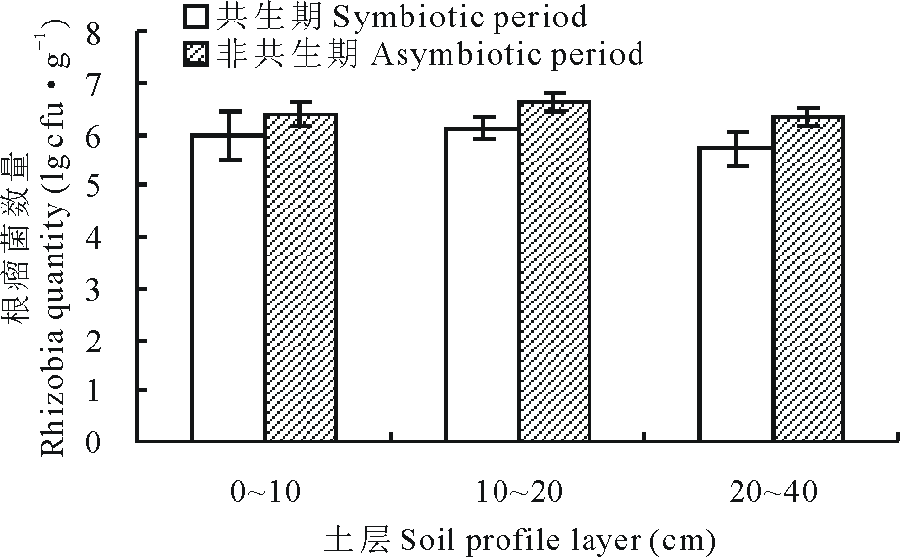 | 图2 冬小麦套种草木樨共生期与非共生期根瘤菌变化Fig.2 Rhizobia dynamically during the symbiotic period and asymbiotic period winter wheat interplanting sweet clover |
2.3.1 不同土层理化指标与根瘤菌数量的相关性分析 土壤理化性质与根瘤菌数量进行相关性分析, 结果(表2)表明, 整体上0~40 cm土层的根瘤菌数量与全盐呈高度相关且相关性显著(P< 0.05), 与水分、碱解氮的相关性中等, 与容重、全氮的相关性较弱。在0~10 cm土层中根瘤菌数量与水分、全盐高度相关且相关性显著(P< 0.05), 与容重、碱解氮、有效钾呈中等相关, 容重与根瘤菌数量相关性显著(P< 0.05), 与有效磷相关性较弱; 10~20 cm土层中根瘤菌数量与全盐高度相关且相关性显著(P< 0.05), 与水分、容重、全氮、碱解氮呈中等相关, 水分与根瘤菌数量相关性显著(P< 0.05), 与有效钾的相关性较弱; 20~40 cm土层中根瘤菌数量与全氮高度相关且相关性显著(P< 0.05), 与全盐、有机质、碱解氮呈中等相关, 与水分、有效钾的相关性较弱。
| 表2 根瘤菌数量与土壤理化性质的相关性分析 Table 2 Correlation analysis of rhizobia quantity and soil physical and chemical properties |
2.3.2 根瘤菌数量与土壤理化性质的通径分析 土壤理化性质对根瘤菌数量分布的通径分析(表3)表明, 全盐、有机质、全氮对根瘤菌数量的直接作用较大(P4=-0.64, P5=0.56, P6=-0.50), 其余次为容重(P2=0.45)、水分(P1=-0.40)、有效钾(P9=0.39)、有效磷(P8=-0.29)对根瘤菌数量的间接作用; 水分、全盐、有机质、全氮、碱解氮、有效钾通过容重、有效磷对根瘤菌数量的影响均有促进作用, 但作用较小(P:0~0.07); 全氮、碱解氮、有效磷、有效钾通过有机质对根瘤菌数量的影响相对较大(P:0.31~0.39)。在最优回归方程中(相关系数R=0.9399; Y=15.36-0.66X3-0.76X4+0.09X5-1.16X6; 决定系数 0.8833; 剩余通径系数=0.3416), 全氮对根瘤菌数量的影响最大, 其次为全盐、pH、有机质。
| 表3 土壤理化性质对根瘤菌数量分布的通径分析 Table 3 Path coefficients between rhizobia quantity and 9 soil factors |
目前国内外相关学者在根瘤菌领域的调查研究中, 大多是对土壤根瘤菌的调查鉴定进行分子生物学方向的研究或对不同土层的豆科植物结瘤数量及其与土壤理化性质的相关性研究, 在对不同土层土壤根瘤菌的动态分布研究报道较少。本试验研究发现, 4-6月0~40 cm土层的草木樨根瘤菌数量持续增加, 到7月前后根瘤菌数量达到最高值, 之后根瘤菌数量开始减少, 但土壤根瘤菌的总体水平较高, 为105~106数量级。王海霞等[31]在宁夏苜蓿土著根瘤菌的研究中发现根据地域性、气候条件、土壤结构及理化性质的差异性, 根瘤菌的数量为103~104数量级。师尚礼[32]在苜蓿根瘤菌的有效性及影响因子的研究表明, 气候条件、土壤墒情、结构及理化性质等组合条件对土壤根瘤菌数目产生影响。4-6月土壤根瘤菌数量持续增加可能与土壤墒情及温度有一定的相关性, 冬小麦及草木樨根系在春季活动旺盛, 根系分泌物开始积累, 对根瘤菌的增殖有促进作用; 7月冬小麦刈割对草木樨的荫蔽作用减弱, 生长抑制解除, 草木樨迅速生长, 支细根扩散部大部分分布在0~20 cm土层, 有机质、热交换条件较下层好, 根瘤及根系分泌物也较多, 因此该土层根瘤菌数量最多(0~20 cm> 20~40 cm); 8月20~40 cm土层根瘤菌数量达到最高值, 在草木樨根系结瘤的相关研究中发现, 8月草木樨主根系向下延伸, 支细根扩散, 深土层结瘤数目增加[10, 27, 33, 34, 35], 但根系结瘤量与土壤根瘤菌数量是否有直接关系, 还有待进一步证实; 9月气温转凉光照变短土壤根瘤菌生长条件变差, 可能导致了根瘤菌数量下降。因此, 根瘤菌数量在4-9月呈单峰曲线规律分布。Mahler等[36]发现国外土壤中根瘤菌在气温低时数量多, 夏末秋初数量最少; 本研究与周智彬等[29]的根瘤菌数量动态变化研究结果一致, 闫晓宁[37]研究发现根瘤的分布为0~20 cm> 20~40 cm, 吕秀华[38]在东北天然草原的相关研究中也发现其呈单峰曲线规律。
在共生期土壤受上年留茬及植物根系的腐烂分解及根瘤菌固氮作用的影响, 增加了土壤氮素余量及有机质含量, 从而减少了草木樨根部土壤区域的氮阻遏效应, 并为根瘤菌的增殖提供了优良的物质条件; 在非共生期生长抑制解除, 固氮作用达到最大化, 小麦根系对根瘤菌的阻遏作用消失, 腐烂的冬小麦根系也提供了一定的养分, 提升了根瘤菌生长的环境, 故而在非共生期根瘤菌数量较高[31, 39, 40]。
土壤水分含量的高低直接影响植物及其微生物的生长。4月气温低, 植物需水量少, 又受冬、春灌水影响, 所以此时土壤含水量最高; 6月底7月初冬小麦开始收获, 气温升高, 地表覆盖度降低, 灌水减少引起土壤含水量低; 7月以后进入草木樨管理期, 补充了灌水, 地表覆盖度增加, 水分损失减少, 此时含水量较高。共生期0~40 cm土层土壤容重由浅到深持续增加, 与赵亚丽等[41]的研究结果一致; 非共生期0~20 cm土层较高, 草木樨主根系主要分布在该土层, 对土壤的束缚和挤压作用比20~40 cm土层强, 且冬小麦在刈割时还受机械的碾压; 刘慧等[3]发现7-9月草木樨容重增加, 但0~20 cm、20~40 cm间规律不突出。草木樨根系还能分泌部分酸性物质, 引起土壤pH在非共生期较共生期下降0~0.7%[42, 43]。土壤全盐在0~20 cm土层非共生期较共生期降低10.66%~17.61%, 相关研究也表明冬小麦套种草木樨能有效降低0~40 cm土层土壤含盐量, 本试验结果与此一致[2, 24]; 全盐受灌溉水对土壤的冲洗作用及生物排盐作用的影响, 草木樨在8、9月气温降低时生长放缓, 为便于刈割, 灌溉水减少, 所以没有完全将20~40 cm土层中的盐分冲洗到下层, 但9月较4月土壤全盐仍下降3.45%。上一年留在土壤中的植物根系20~40 cm土层比0~20 cm土层少, 且20~40 cm土层易腐烂分解, 所以该土层的有机质、全氮含量在4月最高; 浅土层根系健壮, 在春灌后才完全腐败分解, 且在4月有机质、全氮消耗量较分解量少, 5月0~20 cm土层中有机质、全氮含量达到了最高; 全氮、碱解氮、速效磷、速效钾随着冬小麦、草木樨生长持续下降(除20~40 cm土层有效钾含量)。草木樨在9月刈割时较8月有机质提高7.65%~26.98%、全氮提高16.67%~44.12%、碱解氮(0~20 cm土层)提高20.27%~160.68%、有效钾(0~20 cm土层)提高10%左右; 从4、9月含量变化中, 冬小麦及草木樨生长消耗了土壤有机质、全氮、碱解氮、有效磷、有效钾。相关研究中一致发现套种草木樨能提升土壤基础肥力[1, 2, 24, 25, 26], 是因为本试验在9月采样时草木樨刚刈割, 植物根系还有生命力, 采样时间不同所致。
不同土层土壤理化性质与根瘤菌数量的相关性分析中, 全盐(R=-0.81, P< 0.05)在0~40 cm土层中对根瘤菌数量的影响最大; 0~20 cm土层水分易损失, 又因该地区盐碱含量较高, 所以根瘤菌数量受水分(-0.81≤ R≤ -0.73, P< 0.05)、全盐(R=-0.87, P< 0.05)影响较大; 20~40 cm土层透水、透气性能较上层差, 但植物对氮素的吸收利用最多(46.15%), 土壤全氮(R=-0.88, P< 0.05)含量供给不足间接地影响了根瘤菌的增殖, 成为制约根瘤菌数量的主要因素, 牛红榜等[44]发现植物对养分的消耗会制约土壤微生物的生长繁殖, 彭冠初等[45]发现该地区土壤中严重缺乏有机质、有效氮和有机磷, 肥料的施量不足阻碍了作物产量的提高。
在通径分析中, 土壤理化性质对根瘤菌数量的影响主要取决于全盐、有机质的直接作用及容重、水分、有效钾、有效磷的间接作用; 有机质、有效磷对根瘤菌数量不仅有较强的直接影响, 而且还辅助其他性质促进根瘤菌增殖; 全氮、碱解氮、有效磷、有效钾指标对根瘤菌数量的影响正负不一, 可能是因为施肥不足、施肥比例不适、植物消耗土壤养分未及时补充, 造成了土壤理化性质对根瘤菌数量影响出现差异[44, 46]。最优回归方程也表明全氮、全盐、有机质对根瘤菌数量的影响较大。
综合分析, 土壤全氮、全盐、有机质是影响根瘤菌数量最重要的因素。其中, 根瘤菌数量与有机质呈正相关(P5=0.56), 与全氮、全盐呈负相关(P6=-0.5, P4=-0.64)。有机质是促进根瘤菌的主要因子, 全氮、全盐是制约主要因子。因此, 在栽培过程中应该增加有机肥, 适当减少氮肥施量, 降低土壤盐分, 改善根瘤菌的土壤环境, 促进根瘤菌增殖, 提高固氮量, 同时还给土壤输送了植物及根瘤菌所需的能源物质, 土壤肥力也得到提升[47, 48, 49]。冬小麦、草木樨生长越好, 其根系就越发达, 根瘤菌的数量就会越多[20, 34, 50]。
1)冬小麦与草木樨套种根瘤菌数量在垂直方向上10~20 cm土层中分布最多, 在水平方向上0~20 cm土层在7月最高, 20-40 cm土层8月最高; 非共生期数量高于共生时期, 非共生时期20~40 cm土层的根瘤菌数量增长最多。
2)土壤有机质、全氮、碱解氮、有效磷、有效钾在4-8月均有不同程度的下降, 9月草木樨刈割时较8月有所增加。
3)土壤理化性质对根瘤菌数量的影响中, 整体上0~40 cm土层全氮、全盐、有机质与根瘤菌数量相关性最高, 其中与有机质呈正相关, 与全氮、全盐呈负相关; 有机质、有效磷还辅助全氮、碱解氮、有效磷、有效钾促进根瘤菌的增殖, 0~20 cm土层中水分与根瘤菌数量相关性较高, 20~40 cm土层中全氮的相关性最高。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|