
{kind=link}
{kind=link}
{kind=link}
{kind=link}
草原灌丛化入侵:过程、机制和效应
[王迎新 , 陈先江, 娄珊宁, 胡安, 任劲飞, 胡俊奇, 张静, 侯扶江
, 陈先江, 娄珊宁, 胡安, 任劲飞, 胡俊奇, 张静, 侯扶江* ]
, 陈先江, 娄珊宁, 胡安, 任劲飞, 胡俊奇, 张静, 侯扶江]
|
|


作者简介:王迎新(1990-),男,内蒙古克什克腾旗人,在读博士。E-mail: wangyx12@lzu.edu.cn
草原灌丛化已成为全球性问题,也是生态学、畜牧学研究的热点问题。紧扣草原灌丛化过程、发生机理及其生态学后效等关键问题,介绍了国内外的最新研究进展。灌木入侵途径包括有意识引进入侵和自然入侵2类,入侵过程分为传播到达、定居建群和扩散入侵3个阶段。放牧、火烧、温室气体增加,干旱以及降水格局改变等均可导致草本向木本的转化,且机制复杂,主要有影响草本,木本植物盖度、水分利用效率、化感作用等。木本植物入侵打破了草原生态系统的稳定,表现出沃岛效应,影响动植物的分布以及生态系统的功能和服务。阐明草原“草本-木本”群落演替机制,可为我国研究草原灌丛化和管理提供理论借鉴。
, CHEN Xian-jiang, LOU Shan-ning, HU An, REN Jin-fei, HU Jun-qi, ZHANG Jing, HOU Fu-jiang
Woody-plant encroachment in grasslands has been a much-studied topic in ecology and animal husbandry over the past few decades. This paper clarifes the causes and consequences of this encroachment by reviewing recent progress in the study of invasion stages, invasion mechanisms, the impacts of this encroachment on grassland ecosystems and their aftereffects. The means of woody-plant encroachment include both anthropophyte and natural invasion. In general, this encroachment divides into three stages: dispersal, colonizing and establishing and diffusing. The causes of encroachment have been identified as fire suppression, climate change, changes in nutrient availability and over grazing. These factors effect encroachment through changing plant coverage, soil water use efficiency and allelopathy. We conclude that woody-plant encroachment changes many interfaces in grassland ecosystems and influences the ecological and hydrological processes that lead to grassland degradation and desertification.
草原灌丛化(woody-plant encroachment in grassland)是全球性问题。过去150多年全球草原最主要的变化之一, 就是木本植物扩张[1]。全球草原区域约占陆地总面积的41%, 其中有10%~20%的地区发生了灌丛化, 且大多以放牧业为主[2]。在北美, 灌丛化的非林地面积约占3.3× 108 hm2, 每年以0.2%~0.5%的速度扩张[3]。在南非, 约有1.3× 107 hm2稀树草原发生了灌丛化[4]。在澳洲, Mesquite植物以大约每年0.4%~1.2%的速度入侵[5]。在中国, 草原灌丛化现象也有大量报道[6, 7], 其中, 内蒙古草原的小叶锦鸡儿(Caragana microphylla)灌丛化现象最为典型, 约有5.1× 106 hm2的草原出现了灌丛化[6]。青藏高原东南麓, 从1990年到2009年至少有39%的高山草甸已被灌丛草地取代[8]。
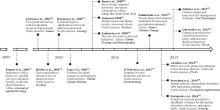
草原灌丛化一直受到各国科学家的高度关注, 在国际生态学顶级期刊阐述了其重要研究进展(图1), 其中, Sala等[1]组织发表在《Journal of Ecology》上的灌木入侵特刊格外引人注目。研究认为干旱以及降水格局改变[12, 14, 19]、放牧[17, 20]、火烧[9, 13, 21]、温室气体增加[10, 18]等均可导致灌木入侵。灌丛化打破了草原生态系统的稳定, 影响生态系统的功能和服务[11, 15, 16, 22, 23]。本研究紧扣草原灌木化的过程、机制、效应及其防控等关键问题, 总结了国内外学者对这一问题的最新研究动态, 旨在阐明草原群落演替机制, 为我国的草原灌木入侵研究和防控提供理论借鉴。
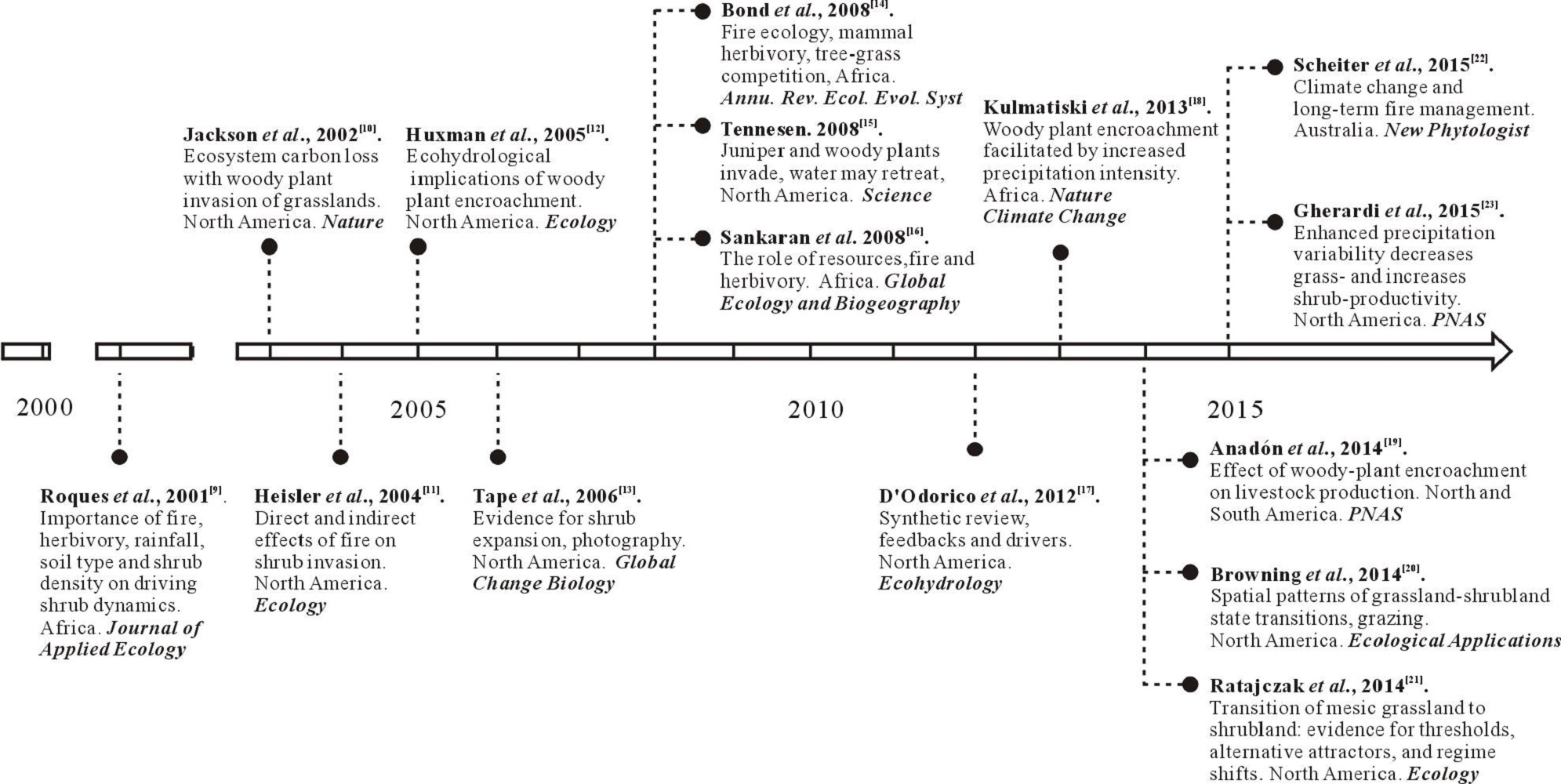 | 图1 灌木入侵的重要研究-问题及分布Fig.1 Key questions and distribution of selected important studies related to woody-plant encroachment |
灌木入侵途径包括有意识引进入侵和自然入侵2类。其中, 有意引进入侵是指人类出于食用、观赏、饲料、引种、环境治理等目的在草原上种植灌木种, 该灌木在新的生态环境内生长, 定殖并迅速扩散而成为优势种群, 对该生物境内的其他种群构成生存威胁。自然入侵指生物靠自身的扩散传播能力或借助于自然力(如鸟类、哺乳动物、蚂蚁和风)入侵[24]。
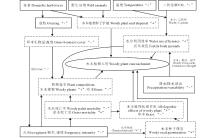
草原灌木入侵过程分为传播到达(dispersal)、定居建群(colonizing and establishing)和扩散入侵(diffusing)(图2):Ⅰ ) 传播到达阶段:外来灌木在自然因子和人类活动的影响下到达其自然演化区域以外的地区, 并成活。Ⅱ ) 定居建群阶段:外来灌木在自然状态下通过与当地生态因子相互作用实现定居和建群。Ⅲ ) 扩散入侵阶段:外来灌木通过自身繁殖和当地物种的竞争等, 不断扩大分布区[25, 26]。
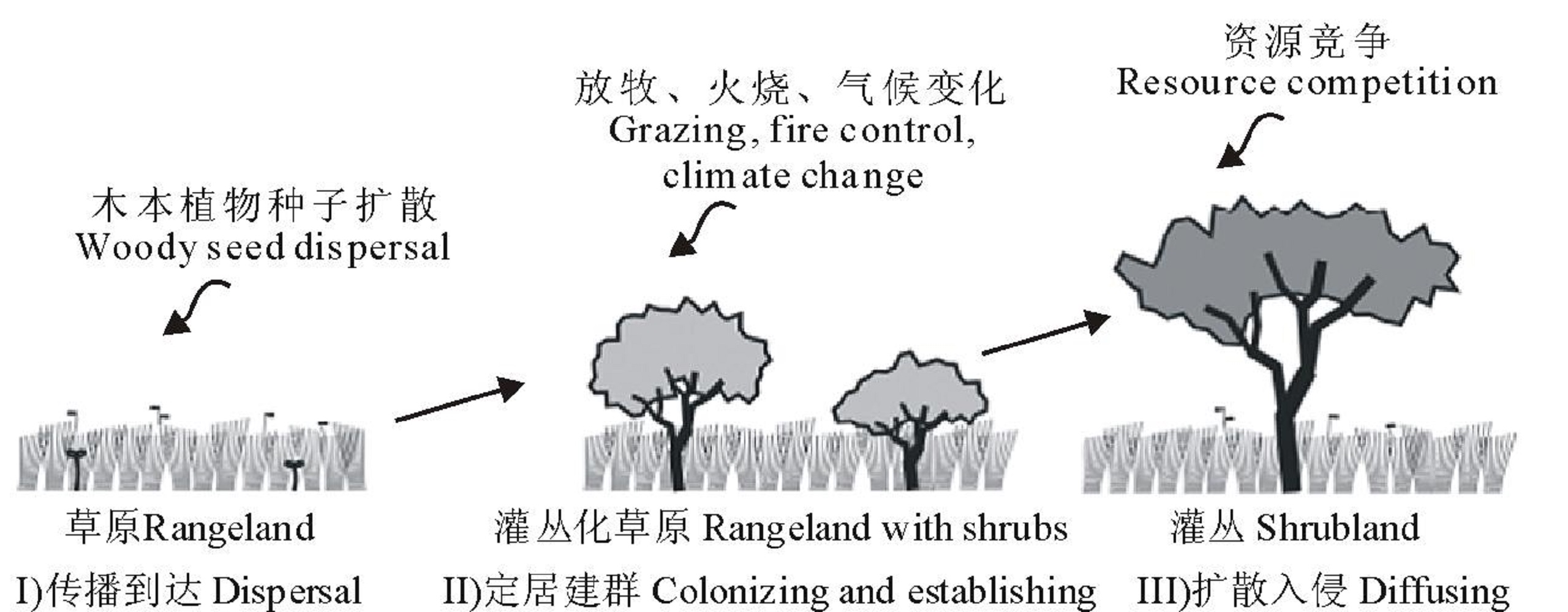 | 图2 草原木本植物入侵过程木本植物在新的环境中入侵成功, 机制非常复杂, 与植物自身属性、功能(图3)和外界环境(降水格局改变、温室气体增加、气温升高、放牧以及火烧等)息息相关。Fig.2 Stages of woody-plant encroachment |
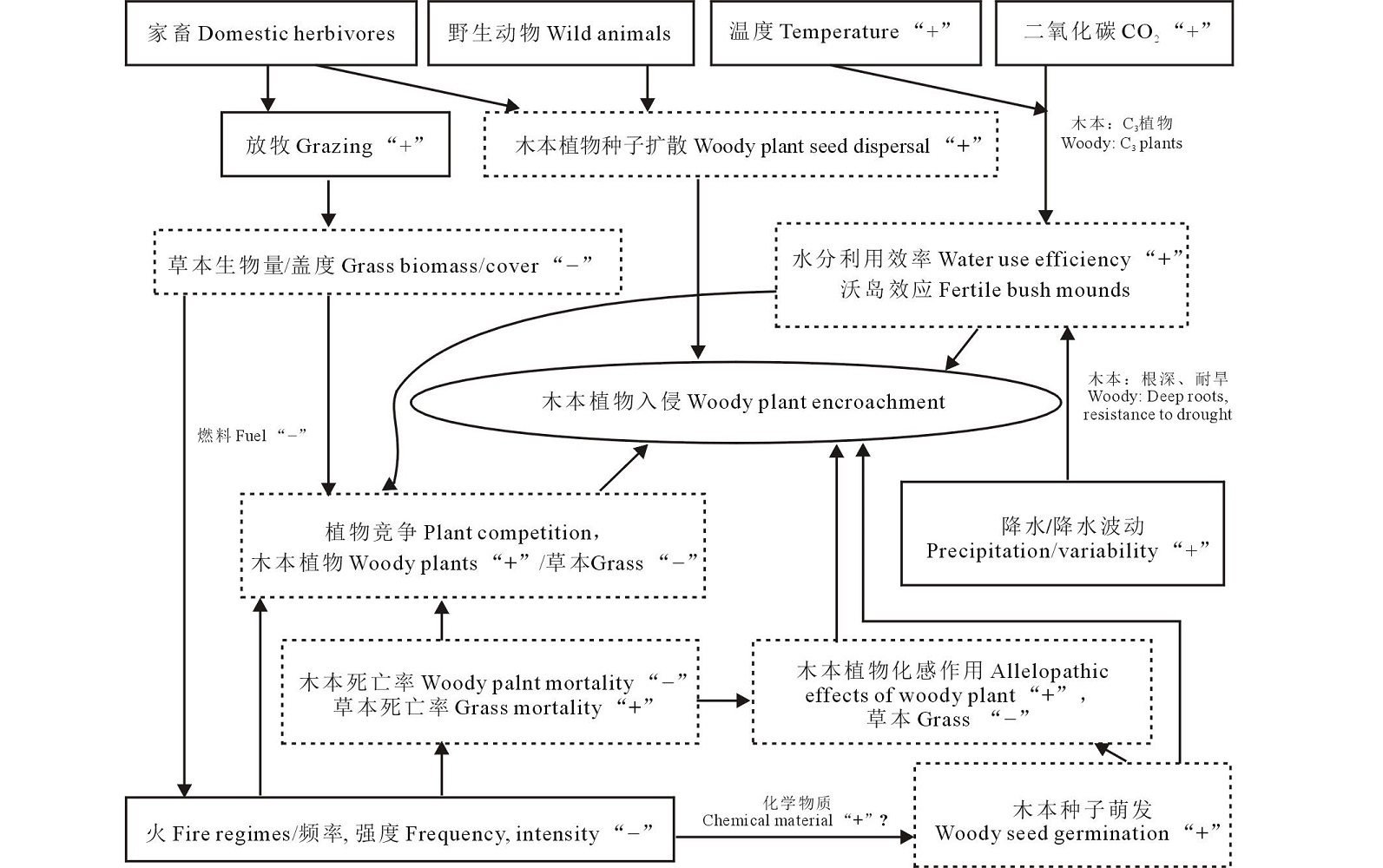 | 图3 木本植物入侵机制 实线矩形代表驱动因子, 虚线矩形代表影响进程, “ +” :增加; “ -” :减少Fig.3 Mechanisms of woody-plant encroachment Solid rectangle represents the driving factor, dotted rectangles represent influence the process, “ +” : increase; “ -” : decrease. |
木本较草本植物根深, 耐旱且水分利用方式及对水分的需求程度不同。Darrouzet-Nardi等[27]在Sierra Nevada山地草甸研究发现大部分的灌木更多利用深层土壤水分, 但也会利用 10%~30%的浅层土壤水分(< 30 cm), 而依赖浅层土壤水分的草本则因降水量减少而逐渐衰落。降水强度的增加可以促进深层次土壤水分的储存, 促进木本植物的生长和分布[28, 29, 30]。相反, 研究发现山地草甸在干旱胁迫作用下, 草本盖度因灌木去除显著增加, 随着土壤水分增加呈线性增加趋势[31]。另外, 在极端降水天气下, 地表径流常常会导致土壤表层N和其他营养元素的淋洗, 从而更有利于深根性的木本植物生长[32]。降水波动(precipitation variability)可以提高木本植物种间竞争、定居能力及丰富度, 引发入侵[26]。Holmgren等[33]分析遥感数据发现, 在全球尺度下, 降水波动对木本盖度的影响因区域各异, 南美显著增加, 澳洲显著降低。Heisler等[11]通过北美6年的模拟实验研究得出, 降水波动显著影响生态系统初级生产力, 多年生草本下降了81%, 木本增加了67%。究其原因, 草本植物的根系分布和吸水能力显著低于木本植物, 而木本植物根系在湿季能储存足够的水分以备旱季所需。
灌木的生长受到低温的限制。气候变化导致低温升高、冻害减少, 降低灌木的死亡率, 为灌木的扩张和定居创造了条件。北美洲西南部荒漠的典型植物Larrea tridentata能耐受的极端低温为-20~-18 ℃左右[34]; 南非稀树草原也发现气候变暖降低了对极端低温敏感的灌木死亡率[35]。气候变暖会加速极地土壤C、N循环以及凋落物分解, 进而刺激微生物活动并促进苔原灌丛化的发生[36]。与此类似, 气候变暖也会导致高寒地区灌木向高海拔入侵。如西班牙中部山脉受气候变暖的影响(最低温和最高温均升高), 原本生长在低海拔地区的刺柏属(Juniperus)灌木植物逐渐取代以羊茅属(Festuca)植物占优势的草本植物群落[37, 38]。
另外, CO2浓度增加也可促进木本入侵。木本是C3植物, CO2可以通过改变水分利用效率、光合速率以及光合养分的利用率来影响植被生长[39]。大气中CO2浓度升高会增加灌木胞间CO2浓度与大气中CO2浓度梯度, 增强灌木的水分利用效率, 使得灌木比草本的竞争力更强[40]。
放牧家畜选择性采食削减了草本植物的竞争力[41], 践踏和排泄物改变了土壤碳库、氮库及分配[42]。过度放牧, 尤其是长期过度放牧后, 减低牧压甚至禁牧的情况更利于木本植物的建立[43]。Christensen等[44]模拟了14种不同放牧率下植被动态, 结果表明过度放牧增加木本植物的分布, 取代草本植物成为优势种, 而适度放牧可以促进草本生长和营养物质循环。Allred等[45]也发现放牧降低了北美草原多年生禾草的组分, 重牧下的木本植物的盖度高于轻牧和对照。牲畜可以作为木本种子扩散的载体。在澳洲, 绵羊和有袋动物是桉树(Eucalyptus spp.)最主要的种子扩散载体[46]; 在北美, 家畜传播扁担杆属(Grewia)灌木种子的距离远远超过自然因素的限制[47]; 在非洲, 捻角羚(Tragelaphus strepciseros)以采食-排泄的方式来扩散滨藜属(Atriplex)植物的种子[48]。
火烧(fire regimes)包括火烧频率、强度、大小及其时空组合, 对维持群落 “ 草本-木本” 的平衡起到重要的作用[49]。木本植物耐火性好, 木质素丰富且分蘖点较高[50], 而草本植物短期恢复能力强。轻度、低频火烧对木本植物损伤较小, 却降低了作为燃料的草本植物及其枯落物的生物量, 有利于木本的生长[51]; 而重度、高频的火烧对木本和草本都会造成损害, 木本植物生长周期长, 恢复速率低, 在一定程度上限制了木本的扩张[52]。另外, 火烧产生的次生物质(如氰化物)促进灌木种子的萌发[53], 过火后土壤微生物的群落结构和营养食物网的改变, 有利于入侵植物竞争资源[54]。
各因素与植被具有复杂的相互作用, 降水量决定植被类型和土壤属性, 土壤属性可以被火烧改变, 而火烧与放牧的相互作用可以决定植被的变化, 植被的变化转而会影响火烧和放牧机制[55, 56, 57]。通过利用Lefkovitch阶段分类矩阵模型研究美国San Carlos东部灌木石南茄(Fabiana imbricata)在火烧和降水共同作用下的种群动态表明, 夏季火烧和春季丰富的降水最有利于石南茄种群扩展, 并通过模型预测, 石南茄种群在火烧频率为每4年1次到每100年1次的范围内都能增大, 在火烧为每6~10年1次的频率下, 石南茄种群的增长率最大[58]。Ratajczak等[59]在美国Kansas东北部生态试验站, 利用当地不同年份的放牧率以及火烧频率数据进行研究分析。研究表明, 在每年1次的火烧频率下, 放牧地灌木盖度有小幅度增加; 在火烧频率为每年1~3次的放牧地, 在前17年(1980-1997年)中, 灌木盖度以每年小于0.5%的幅度缓慢增加, 1998-2001年灌木盖度以每年5%~10%的幅度增加, 2001-2012年灌木盖度以每年1.4%~2.7%的幅度增加。
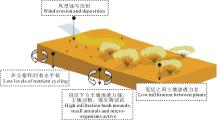
灌木通过改变土壤结构、微生物的生物量、土壤湿度和小气候, 并将有机物质集中于其树冠层之下, 从而对养分的空间分布和循环产生影响, 这就是所谓的“ 沃岛” 效应[60, 61](图4)。首先, 气候变化、过度放牧等会破坏草原植被的均一性, 致使部分地表裸露。裸露的地表经受高温和高蒸发的影响, 阻止或延迟了有机氮的吸收、反硝化以及氨化等一系列过程, 导致养分流失[62]。风和水携带的富含养分的土壤、碎屑以及种子会聚集在灌木植物冠层下, 提高冠层下方土壤的渗透能力和肥力[63]。这些效应综合在一起, 进一步加强土壤资源在灌木植物冠层下的聚集, 使灌丛对环境扰动具有更强的抵抗力[64, 65]。其次, 灌木植物从周围吸收养分, 通过自身凋落物分解, 构成一个养分库, 增加了灌丛的自持能力[66]。最后, 土壤微生物和土壤动物在灌丛下的活动较为强烈, 进一步促进了灌丛下养分循环。而草本植物生物量的降低, 进一步加强了灌木植物的定居及稳定发展[67]。Myers-Smith等[68]在澳大利亚灌丛化草地中的观测发现, 与草地斑块相比, 灌丛斑块灌木冠层下方养分循环和水分入渗率指数都较大。Parizek等[69]得到巴塔哥尼亚东北部的灌丛化草地中灌丛斑块土壤容重低于草地斑块, 土壤水分入渗速率和土壤有机质含量高于草地斑块, 灌丛化草原中草地斑块的土壤侵蚀程度增加。Lett等[70]对北美普列里高草草原(tallgrass prairie)群落进行研究指出灌木入侵后草原的碳库和氮库储量高于入侵之前。Li等[71]比较了中国北方科尔沁地区小叶锦鸡儿灌丛斑块及其周围草地斑块土壤理化性质, 认为灌丛斑块冠层下方土壤容重较低, 土壤有机碳和全氮含量比草地斑块分别高23%~32%和14%~27%, 灌丛斑块有更强的土壤持水能力。
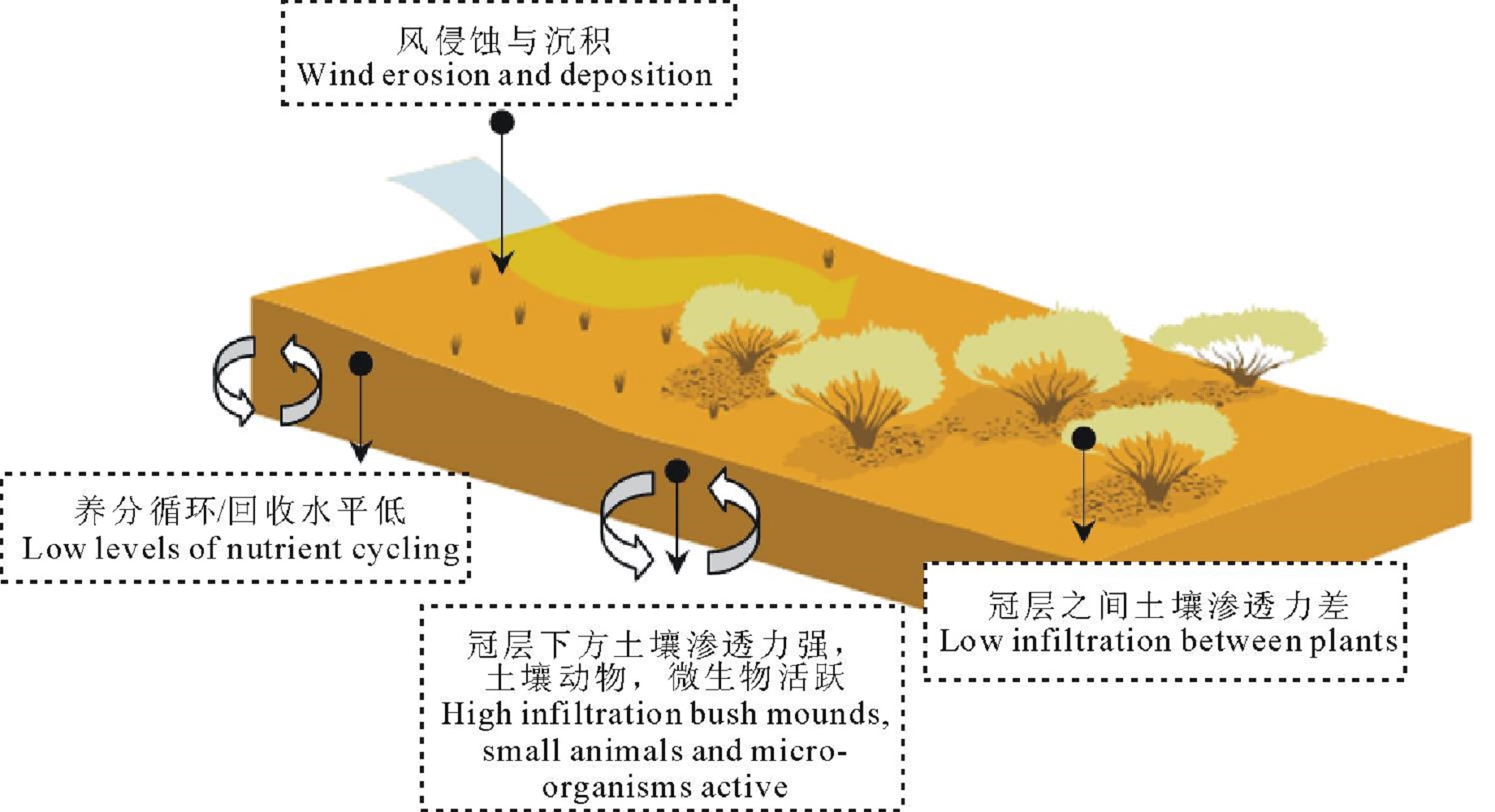 | 图4 沃岛效应Fig.4 The capture of water and nutrients by fertile bush mounds |
斑块尺度上, 灌丛自身以及冠层下草本的生产力增加, 物种组成改变。灌木入侵带来了新的生命形式, 新的物候类型(如侵入美国西部灌丛大草原的Bromus tectorum), 新的方式摄取资源(如夏威夷岛的固氮植物Myrica faya), 新的演替生态位。同时, 木本植物的适口性、植株生理特征等因素增加了动物的采食难度, 为其冠层下方的其他植物提供一定的庇护。加利福尼亚草原灌丛化过程中, 地上、地下生物量呈增加趋势, 灌丛斑块更容易维持较高的生物量[72]。内蒙古典型草原在小叶锦鸡儿入侵后, 多年生根茎禾草、多年生丛生禾草与多年生杂类草有所增加, 而其他一年生的植物则逐渐减少, 改变了草地的物种更替[7]。
群落尺度上, 在多因素(自然, 人为因素)的干扰下, 木本入侵后, 草原生产力和多样性的变化具有不确定性。美国弗吉尼亚的灌木林区叶面积指数达到峰值时, 灌木下层接受的光照仅为上层的0.7%, 草本生产力下降[73]。Bekele等[74]得出在木本植物密度(583.2± 79.3) 树· hm-2下草本植物生物量为(2625± 300) kg· hm-2, 而当木本植物密度为(2827± 276) 树· hm-2时, 草本植物的生物量只有(1176± 110) kg· hm-2。近100年来, 美国新墨西哥州的草原灌丛化过程中, 物种丰富度呈下降趋势; 在短时间尺度内, 与草本植物占优势的群落相比, 灌丛化区域的群落稳定性较低[3]。
木本化的出现, 改变动物赖以生存的生境和资源, 影响动物的生产力和习性。高山苔原灌木入侵后, 北极地松鼠(Spermophilus parryii)觅食策略发生改变, 觅食成本增加。非洲喀拉哈里牧场南部热带稀树草原灌丛化过程中, 哺乳动物的数量随灌木盖度的不同而不同, 灌丛化对非洲野猫(Felissilvestris lybica)、条纹臭鼬(Mephitis mephitis)、好望角狐(Otocyon megalotis)、灰沼狸(Suricata suricatta)产生负影响, 对黄色猫鼬的影响呈驼峰形曲线状态[75]。木本盖度的增加可以显著增加山羊(Capra aegagrushircus)的采食量和生产性能[76]。Anadó n等[19]系统研究了南北美洲草原木本植物入侵对家畜生产力的影响, 发现在高生产力草原, 增加1%的木本植物盖度会降低2.5%的家畜生产力, 而在低生产力草原, 木本植物盖度增加反而对家畜生产力有积极作用。
人类是生态系统的设计者, 管理者和受益者。木本植物入侵三阶段中, Ⅰ → Ⅱ 演替过程可以逆转, Ⅱ → Ⅲ 过程难以逆转。人类需重新思考草原“ 草本-木本” 转化的逻辑, 变输血为造血, 变对抗为兼容。
合理放牧和火烧可以驱动Ⅱ → Ⅰ 过程的发生。一些家畜(如山羊偏好采食灌木的叶, 花或种子直接降低了灌木的生物量和种子扩散的机会。火烧可直接杀死植物, 而草本植物在短期内恢复能力更强, 一定程度上抑制了木本的扩张。Roques等[9]在非洲南部的Swaziland地区, 分别于1947, 1971, 1979, 1990年, 利用航空照片对该地萨王纳草原103个固定地点进行分层抽样, 结合当地火灾频率、放牧强度、降水量等数据进行研究分析后发现, 频繁火灾、低强度的放牧以及干旱, 可以减少高大灌木的密度。在澳州北领地, 土著居民用火烧管理、更新灌木草原的办法经久不衰[77]。在北美高原草原, 大约每3~5年的火烧间隔有利于生态系统健康发展[78]。在我国内蒙古锡林河流域, 1~2年的火烧频次明显抑制了小叶锦鸡儿灌丛的发展[6]。
一旦Ⅱ → Ⅲ 过程已发生, 根除和控制灌木发展就会非常困难。根除和控制已入侵灌木的方法主要有机械法(适用于种群数量小的灌木, 包括拔除、砍倒、火烧、水淹、光照和遮阴等); 化学法(如使用专一性除草剂); 生物防治法(利用入侵灌木的天敌控制其种群密度和扩展速度); 综合管理系统(integrated brush management)[25]。
草原灌丛化已经成为全球性热点问题。灌木入侵打破了草原生态系统的稳定, 影响生态系统的功能和服务。因此, 研究木本植物的入侵格局、机制、规律和趋势, 并探索防治、管理的方法是面临的长期而艰巨的任务。首先, 尽管已从水分、温度、土壤、生物以及人为活动等自然与人为因素综合作用角度对草原灌丛化的成因进行了深入分析, 对草地灌丛化的过程和机制有了较为深入的认识, 但这些研究结果多基于对自然过程的观察和分析, 缺乏基于这些过程的大尺度、长时间野外控制实验证据的支持。其次, 由于对草地灌丛化过程和机制的理解仍不完善, 导致灌丛化草地能否恢复, 在哪种条件下可以恢复的全球性认识不完全、不一致。第三, 灌丛化是否会造成生态系统的退化仍然没有定论。Eldridge等[4]基于全球尺度的分析, 认为灌丛化并未在全球尺度上造成普遍性的生态环境退化。最后, 自然和人工方法及其组合, 哪种控制草地灌丛化的方法更为高效仍无定论。因此, 未来灌木入侵研究应在几个方面推进:1)全球尺度长期控制实验的实施与数据整合分析; 2)入侵过程和机制需进一步明确。例如, 如何综合多重因素建立木本植物入侵的生态学框架等; 3)控制管理措施科学有效, 更系统化。尤其是灌木入侵后, 应该以自下而上的视角, 复杂的思维方式, 生态文明的视野, 复合生态伦理机制, 寻找木本植物与草原生态系统各组分的共存之道; 4)灌木入侵与动物生产、人类健康的关系。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|