







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
不同品种高粱幼苗在干旱复水过程中的生理生态响应
[刘婷婷1, 2  , 陈道钳
, 陈道钳3 , 王仕稳1, 3, * , 殷俐娜1, 3 , 邓西平1, 3 ]
, 陈道钳, 殷俐娜|
|


作者简介:刘婷婷(1990-),女,河南郑州人,在读硕士。E-mail:liutingtingUCAS@126.com
干旱适应能力不仅包括干旱胁迫期间的抗旱能力,也包括水分胁迫解除之后的恢复能力。以8个高粱品种为材料,采用盆栽称重控水法对高粱幼苗进行干旱及复水处理,通过测定生物量、相对含水量、水势、渗透势、光合参数等生理指标的变化分析不同品种高粱抗旱能力和旱后复水恢复能力的大小及其与干旱适应性的关系,并筛选出能有效指示干旱适应能力的生理指标。结果显示,干旱胁迫条件下,“辽杂21”生物量与对照相比无显著差异,抗旱能力较强;“甘蔗籽”和“Moench.cv.Gadambalia”高粱生物量分别下降38%、34%,抗旱能力较差。各品种高粱的相对含水量、水势、渗透势、叶绿素含量、净光合速率、蒸腾速率、气孔导度等均有所下降,复水后,这些生理指标恢复到对照水平,各品种开始恢复生长。由各生理指标与抗旱能力、旱后复水恢复能力和干旱适应能力之间的相关分析结果表明,高粱的干旱适应性与复水恢复性呈显著正相关( r=0.85),与抗旱性的相关系数是0.46,复水恢复能力在高粱干旱适应性中起着更重要的作用。高粱的抗旱性和旱后复水恢复性具有不同的生理基础,在干旱胁迫期间维持较高的净光合速率、较低的蒸腾速率有助于提高抗旱性,维持较高叶片相对含水量有助于提高旱后复水的恢复能力,叶片相对含水量、净光合速率可以作为筛选高粱干旱适应性的生理指标。
Adaption to drought in plants includes drought resistance during the drought period and post drought recovery. The aim of this study was to investigate the relationships between drought resistance and drought recovery and to determine the physiological parameters which could be used to indicate drought adaptability. Eight sorghum cultivars grown in pots were exposed to drought and re-watering treatments during the seedling stage. Biomass, relative water content, water potential, osmotic potential, and photosynthetic parameters were measured. The results showed that “LZ21”“ had the highest drought resistance; biomass production under drought stress was not different to the control (well watered), while “GZZ”“ and “Gadambalia”“ had poor drought resistance producing 38% and 34% less biomass under drought stress. Relative water content, water potential, osmotic potential, chlorophyll content and photosynthetic parameters decreased after imposition of drought but all parameters recovered fully after re-watering. Correlation analysis showed that drought adaption is more closely related to drought recovery ( r=0.85) than drought resistance ( r=0.46). Maintaining higher photosynthetic and lower transpiration rate contributed to drought resistance while maintaining high relative water content improves drought recovery. These results demonstrated that drought recovery ability is more important for drought adaption than drought resistance. The physiological basis of drought resistance and drought recovery are clearly different. Relative water content and photosynthetic rate could be used to select drought adapted cultivars in sorghum.
植物在长期适应干旱环境过程中, 形成了4种适应机制:逃旱性(drought escape)、御旱性(drought avoidance)、耐旱性(drought tolerance)和复水恢复性(drought recovery)[1], 其中, 御旱性和耐旱性被称为抗旱性(drought resistance)[2]。抗旱性是指植物遭受水分胁迫后仍能保持正常生长、开花以及获得产量的能力。复水恢复性指植物经历一段时间干旱在水分条件改善之后恢复正常生长的能力[2, 3]。在实际作物生产中, 作物往往处于干湿交替环境中, 因此, 作物在干旱环境下最终的产量不仅仅受其在遭受干旱胁迫过程中对干旱的抵御和耐受能力影响, 还应包括水分条件改善之后作物从先前损伤当中重新恢复正常生长的能力。因此, 复水恢复能力是作物适应干旱环境的一个重要内容, 也是作物适应干旱胁迫的一个重要机制。例如, 当作物苗期遭受干旱时, 复水恢复能力强的禾本科作物可以通过复水后增加分蘖来保证最后的产量[1, 4]。也有研究表明, 作物不同品种在干旱复水之后的恢复能力并不相同, 认为筛选抗旱性强、复水后恢复速度快的品种对于实际生产具有重要的意义[5, 6]。为了更好地衡量作物对干旱胁迫的反应, Chen等[7]提出了干旱适应性(drought adaptation)概念, 即将作物在干旱和复水过程中的整体表现称为干旱适应性, 其既包括了抗旱能力又包括了复水后的恢复能力[8], 并通过对多个玉米(Zea mays)品种的干旱适应性研究表明干旱适应性受恢复能力的影响更大。
高粱(Sorghum bicolor)是一种公认的综合抗旱性较强的作物, 不仅体现在由御旱、耐旱、避旱等多种机制形成的较强抗旱能力[9], 而且体现在受旱的高粱在复水后, 能够得到不低于水分充足时的产量, 甚至产量超过充分供水水平, 即表现出旱后复水很好的恢复能力[10, 11]。前人对高粱抗旱性的评价研究较多, 同时对高粱抗旱性与相关生理指标, 如叶片水分状况、渗透调节能力、抗氧化能力等之间的关系做不少研究[12, 13], 而对高粱的复水恢复能力及其和相关生理指标的关系关注较少。同时, 是否可以采用干旱适应性定义来衡量高粱在干旱复水过程中的整体响应也缺少相关的研究。因此, 本实验以8个不同高粱品种为材料, 研究高粱苗期干旱适应性与其抗旱能力和复水后的恢复能力的关系, 并进一步探索其干旱适应性、抗旱性、旱后复水恢复能力及其相关生理基础。通过完成本研究, 以探索不同高粱品种其干旱适应性和抗旱能力及恢复能力的关系, 以及影响干旱适应性, 抗旱能力和恢复能力的生理机制。研究结果将为旱区作物品种选择和作物的抗旱育种提供借鉴。
从前期21个高粱品种栽种实验中筛选出8个生育期相对一致且抗旱性和复水恢复性差异较大的高粱品种为试验材料, 供试品种分别为“ Moench. cv. Gadambalia(M.G)” 、 “ 千斤红(QJH)” 、“ 遭烘萼(ZHE)” 、“ 饭高粱(FGL)” 、“ 高八尺(GBC)” 、“ 甘芝甜(GZT)” 、“ 甘蔗籽(GZZ)” 、“ 辽杂21(LZ21)” 。
盆栽试验于2013年5-7月在中国科学院水利部水土保持研究所干旱大厅人工气候室内进行, 试验所用塑料盆规格为直径30 cm, 高28 cm, 每桶装入14 kg黄绵土, 土壤田间持水量为20%, 测得土壤含水量为8.97%。高粱种子用1%次氯酸钠消毒20 min, 之后用蒸馏水冲洗, 选取大小一致的种子点播于塑料盆中。塑料盆置于移动遮雨棚下, 降水时遮上, 除了降水, 其他生长条件均与大田相似。当高粱幼苗第3片叶完全展开时, 定苗为2株每盆。
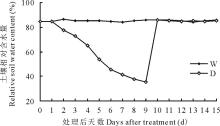
在高粱第8片叶完全展开时, 开始进行干旱-复水处理。实验设置2个水分处理:对照组充分供水(W)和处理组(自然干旱8 d后复水, D)。采用称重控水法于每天下午5点称重测定土壤含水量, 并将同一处理补水至相同含水量值。实验对照组的土壤含水量维持在田间持水量的85%~90%; 处理组土壤经自然干旱, 第8天时土壤含水量下降至田间持水量的35%, 处理第9天复水至对照水平。分别在处理前、处理第8天、复水后第5天采样, 采样时间为每天上午8点到9点, 采样部位为高粱第8片叶。实验过程中对照组(W)和处理组(D)的土壤含水量变化如图1所示。
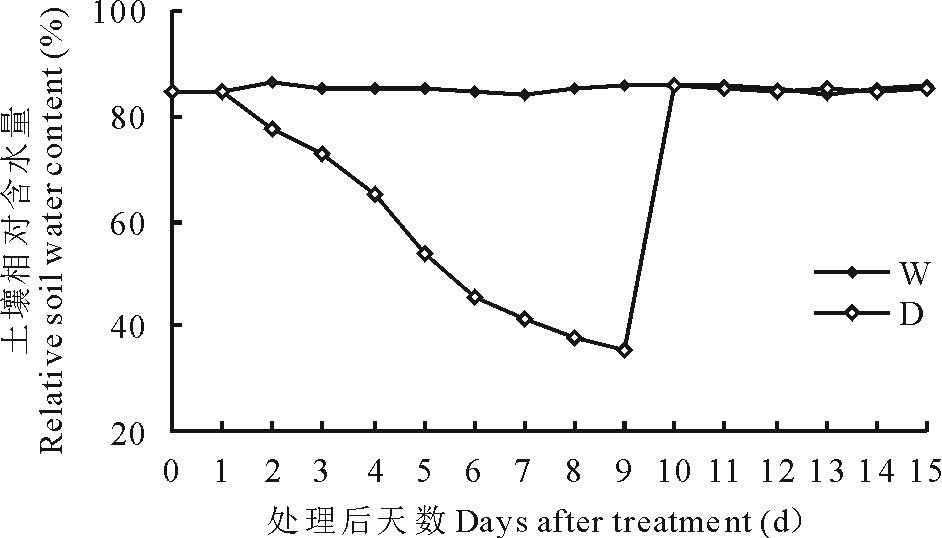 | 图1 对照组(W)和处理组(D)的土壤含水量变化Fig.1 The changes of soil water content during drought stress and re-watering process |
1.2.1 生物量 分别在干旱处理前, 干旱处理8 d后, 复水5 d后采样, 将植株从地表处剪断装入纸袋, 80 ℃烘至恒重, 每个处理分别测定10株。
1.2.2 渗透势 取第8片叶0.5 g装入0.5 mL离心管中, 液氮冷冻30 min, 室温解冻, 将离心管底部扎眼, 放入1.5 mL离心管中, 4 ℃、4000 r· min-1离心5 min, 收集上清液, 用美国Wescor露点渗透压仪5600进行测定[14]。
渗透势计算公式为:Φ =icRT/106
式中:Φ 为渗透势; R为常数, R=8.314; ic为渗透势仪读数(mmol· kg-1); T为绝对温度, T=273+室温(℃)。
1.2.3 水势 于上午6:00采取高粱第8片叶, 用3005型压力室(SEC公司, USA)测定叶水势, 每个处理测定5株。
1.2.4 叶片相对含水量 参考《植物生理学实验指导》方法[15], 从高粱植株上剪去待测定叶片, 擦净, 称重并记录(FW), 然后将叶片浸入蒸馏水中6 h后称重并记录(TW), 之后将叶片置于80 ℃烘至恒重, 称重记录(DW)。
叶片相对含水量=(FW-DW)/(TW-DW)× 100%
1.2.5 叶绿素含量 采用80%丙酮浸提法测定叶绿素含量, 每个处理5次重复[15]。
1.2.6 光合参数 采用美国LI-COR公司生产的Li-6400便携式光合仪, 每天上午测定高粱第8片叶的净光合速率(Pn), 蒸腾速率(Tr)、气孔导度(Cond)等参数。
1.2.7 抗旱能力、恢复能力及干旱适应性计算 在干旱第8天、复水第5天每个处理选取10株在80 ℃烘至恒重后, 称重(DW), 高粱的抗旱能力、旱后恢复能力以及干旱适应能力计算参考Chen等[7]。
植株抗旱能力=
植株旱后恢复能力=
干旱适应能力=
利用SPSS 18.0处理分析数据, 最小显著差异法(LSD)进行方差分析, 显著性水平设为P< 0.05。采用Excel作图, Heatmap Illustrator 1.0做聚类热图。其中, 主成分分析、Pearson相关分析均采用各指标的相对值进行分析作图。聚类热图数据分析时利用各指标的相对值的对数值(以10为底)进行作图。
如图2所示, 水分胁迫显著抑制了高粱的生长发育, 减缓了高粱生长速率, 但不同品种受抑制程度不同。经过8 d的干旱处理, 除了“ 辽杂21(LZ21)” 的地上部生物量与充分供水对照无显著性差异外, 其他品种的地上部生物量均显著降低, 其中, “ 甘蔗籽(GZZ)” 下降程度最严重, 达38%, 其次是“ Moench.cv.Gadambalia(M.G)” 高粱下降了34%。复水5 d后, 所有品种均恢复生长, 表现为生物量累积增加, 但此时的地上部生物量仍低于其对照, 且复水后不同高粱品种的生长速率不同, 表现为高八尺(GBC)> Moench.cv.Gadambalia(M.G)> 甘蔗籽(GZZ)> 饭高粱(FGL)> 遭烘萼(ZHE)> 千斤红(QJH)> 辽杂21(LZ21)> 甘芝甜(GZT), 其中 “ 高八尺(GBC)” 高粱的生物量累积超过100%, 说明其恢复速率最快, 旱后复水的恢复能力较强, “ 甘芝甜(GZT)” 恢复速率最慢, 为9.5%。说明不同的高粱品种具有不同的复水恢复能力。
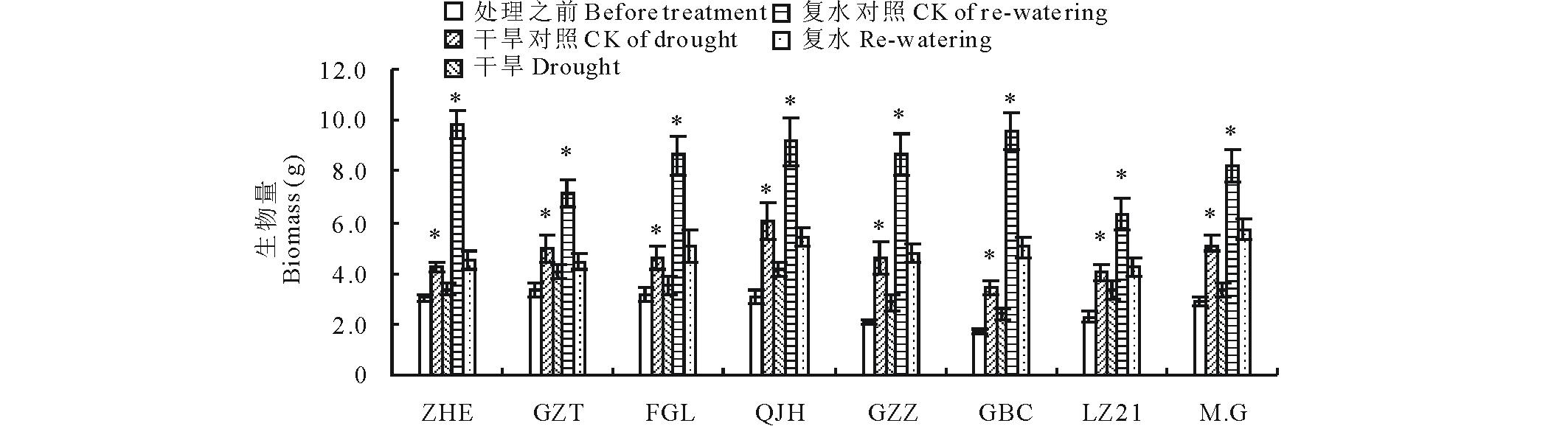 | 图2 干旱及复水对不同品种高粱地上部生物量的影响 Asterisks indicate statistically significant differences between control and treatment (P≤ 0.05).“ * ” 表示处理和对照之间差异显著(P≤ 0.05)。“ * ” means statistically significant differences between treatments (P≤ 0.05).下同 。The same below.Fig.2 The biomass accumulation during drought stress and re-watering |
从图3可看出, 干旱胁迫条件下, 各高粱品种幼苗的叶片相对含水量较对照处理低, “ 千斤红(QJH)” 和“ 饭高粱(FGL)” 的叶片相对含水量分别下降28%和25%, 为所有品种中下降程度最大, “ Moench.cv.Gadambalia(M.G)” 和“ 甘蔗籽(GZZ)” 叶片相对含水量下降程度最小, 分别为5%和4%。复水5 d后, 所有品种的叶片相对含水量均恢复到对照水平。说明干旱胁迫造成叶片发生失水, 失水程度越大, 则该品种维持叶片水分状况的能力就越弱; 复水可迅速改善叶片失水状况。
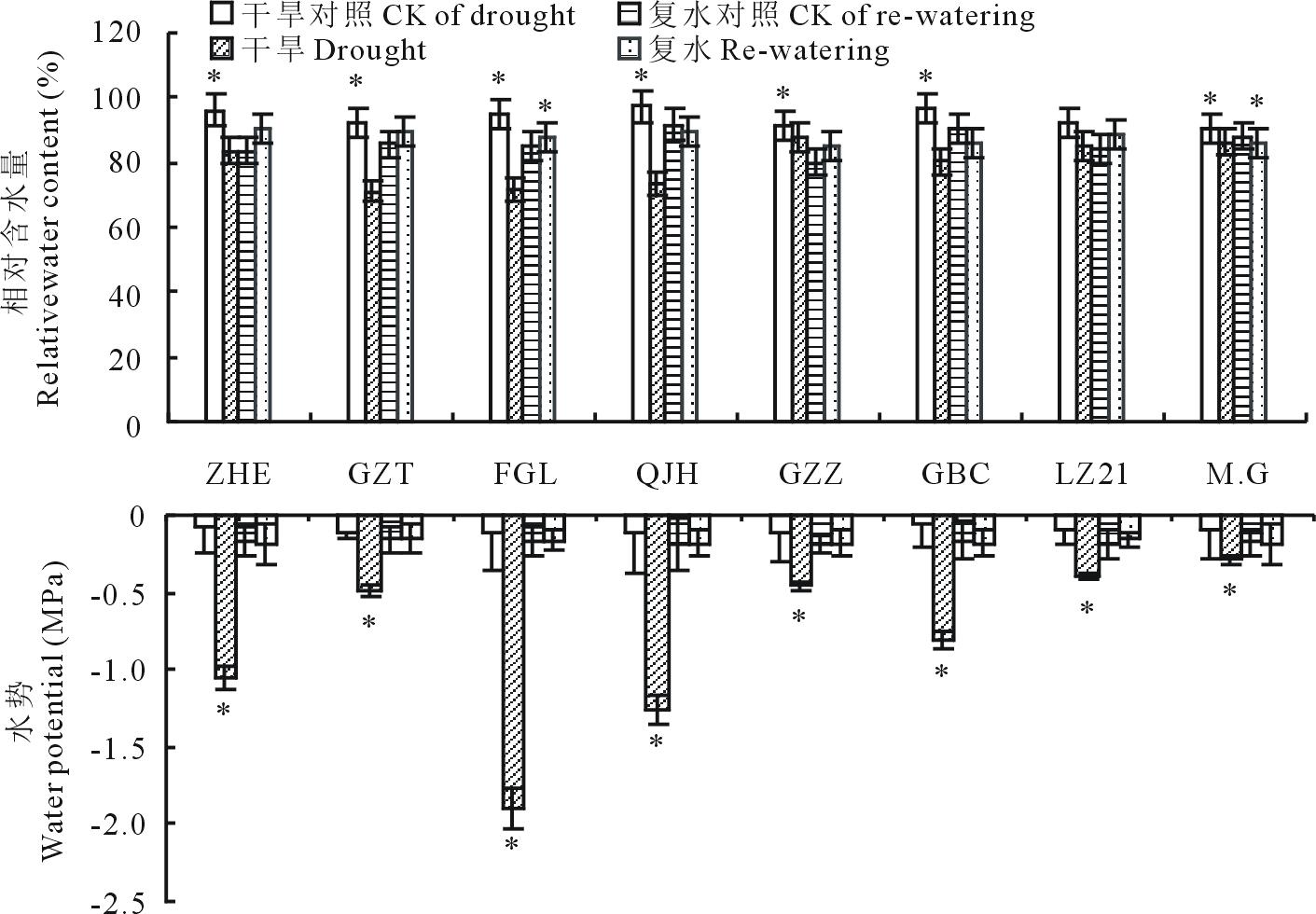 | 图3 干旱及复水对不同品种高粱相对含水量和水势的影响Fig.3 Changes in relative water content and water potential during drought stress and re-watering |
与叶片相对含水量变化相似, 干旱胁迫下, 所有品种高粱的叶水势均显著下降, 其中“ 饭高粱(FGL)” 叶水势下降程度最大, 其次是“ 遭烘萼(ZHE)” , “ Moench.cv.Gadambalia (M.G)” 下降程度最低, 而复水后所有品种的叶水势均恢复到充分供水对照水平。说明不同的高粱品种在干旱胁迫条件下的保水能力呈现出差异性, “ Moench.cv.Gadambalia (M.G)” 高粱干旱期间维持叶片较好水分状况的能力较其他品种更强, “ 饭高粱(FGL)” 维持叶片水分状况的能力最差, 但复水后所有品种高粱叶片水分状况均能恢复到正常水平。
渗透调节是植物应对干旱胁迫的一种主要调节方式。如图4所示, 干旱胁迫使所有高粱品种的渗透势均显著下降。“ 饭高粱(FGL)” 、“ 千斤红(QJH)” 分别下降144.2%、126.0%, 下降幅度较大, “ 甘芝甜(GZT)” 、“ Moench.cv.Gadambalia(M.G)” 下降幅度最小, 分别为44.5%和46.7%。复水后, 不同高粱品种的渗透势变化表现出显著差异。除“ Moench.cv.Gadambalia(M.G)” 高粱的渗透势未完全恢复到对照水平外, 其他品种的渗透势均完全恢复, 甚至显著高于对照水平。说明干旱胁迫期间不同的高粱品种具有显著不同的渗透调节能力, “ 饭高粱(FGL)” 、“ 千斤红(QJH)” 的渗透调节能力优于其他品种, 且复水后的恢复能力也较强, 而“ Moench.cv.Gadambalia(M.G)” 高粱不仅渗透调节能力较弱, 且在复水后的恢复程度也较其他品种慢。
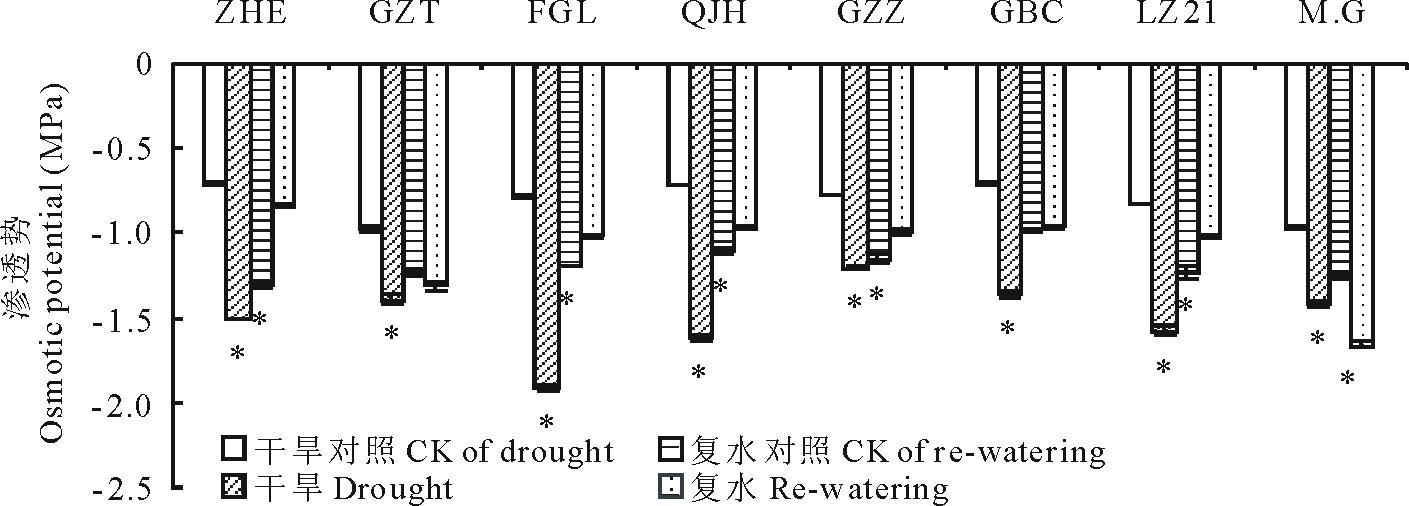 | 图4 干旱及复水对不同品种高粱渗透势的影响Fig.4 Changes in osmotic potential during drought stress and re-watering |
膨压的维持是细胞正常生长的基础, 干旱条件下较高的膨压维持能力有助于维持较好的光合作用, 于作物生长有利。由图5可知, 不同高粱品种细胞膨压对干旱胁迫的响应并不相同。“ Moench.cv.Gadambalia(M.G)” 、“ 甘芝甜(GZT)” 、“ 甘蔗籽(GZZ)” 、“ 辽杂21(LZ21)” 在干旱条件下的膨压较对照高, 具有更强的膨压维持能力, 而“ 遭烘萼(ZHE)” 、“ 饭高粱(FGL)” 、“ 千斤红(QJH)” 和“ 高八尺(GBC)” 的细胞膨压较对照有所降低, 说明干旱胁迫使得这些高粱品种叶片维持细胞膨压的能力丧失, 抗旱性较差; 复水后, 除遭烘萼(ZHE)之外, 所有品种的细胞膨压均恢复到对照水平。
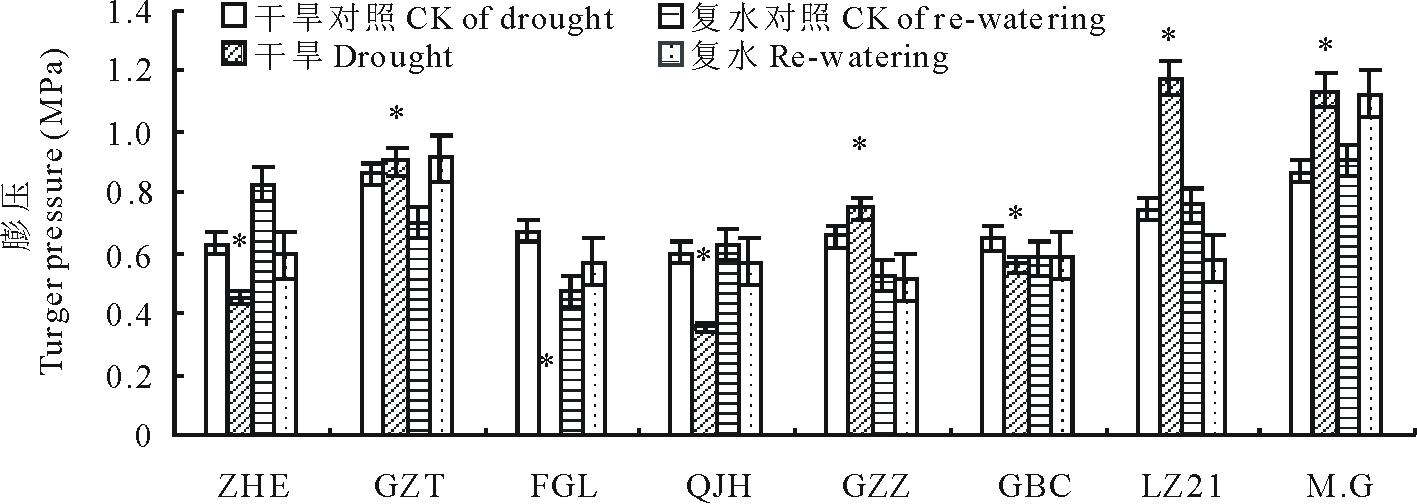 | 图5 干旱及复水对不同品种高粱膨压的影响Fig.5 Changes in turgor pressure during drought stress and re-watering |
如图6所示, 受干旱胁迫的影响, 所有高粱品种的叶绿素含量均较对照显著降低, 但降低程度不同, 说明干旱胁迫造成各高粱品种叶片内叶绿素不同程度的降解。其中, 降低程度最大和最小的品种分别为“ Moench.cv.Gadambalia(M.G)” 和“ 千斤红(QJH)” , 分别下降了46.4%和13.9%; 复水处理使得各高粱品种的叶片叶绿素含量开始有所恢复, 复水5 d后, 各品种的叶绿素含量已经恢复到充分供水对照水平, 甚至高于对照含量。
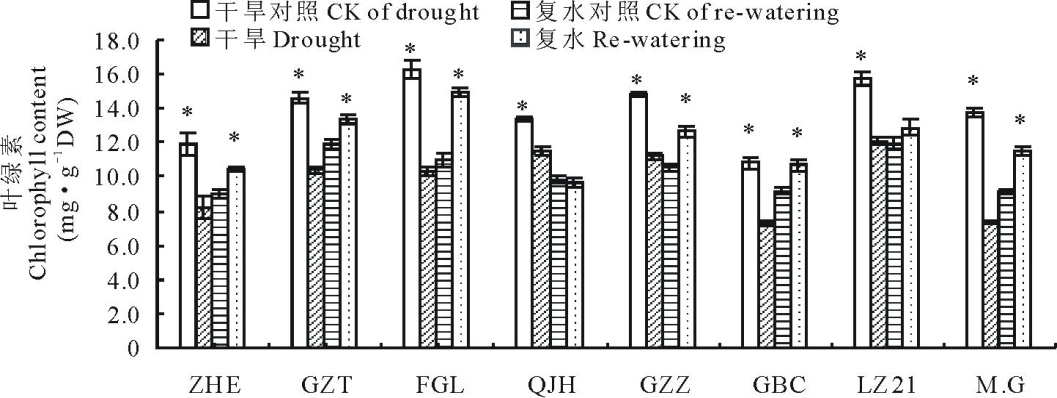 | 图6 干旱及复水对不同品种高粱叶绿素含量的影响Fig.6 Changes in chlorophyll content of 8 sorghum lines during drought stress and re-watering |
水分胁迫通常会降低气孔导度, 抑制作物的光合作用和蒸腾作用。在本实验中, 如图7所示, 胁迫后所有高粱的净光合速率、气孔导度、蒸腾速率均受到显著抑制, 尤其是“ Moench.cv.Gadambalia(M.G)” 、“ 甘芝甜(GZT)” 、“ 甘蔗籽(GZZ)” 、“ 辽杂21(LZ21)” 的净光合速率、气孔导度、蒸腾速率几乎降低为0, 说明这4个品种的光合作用受抑制程度最严重, 抗旱性较差。复水后, 不同品种高粱的净光合速率、蒸腾速率和气孔导度都有所恢复, 但恢复速度呈现品种差异。“ Moench.cv.Gadambalia(M.G)” 、“ 甘芝甜(GZT)” 的净光合速率、蒸腾速率先恢复至对照水平, 其他品种则恢复较慢。就气孔导度而言, 复水后所有高粱品种均未恢复到对照水平。
 | 图7 干旱及复水对不同株系高粱净光合速率、蒸腾速率和气孔导度的影响Fig.7 Changes in photosynthesis rate, stomatal conductance, and transpiration rate during drought stress and re-watering |
如图8所示, 将高粱在干旱胁迫期间和复水恢复后的相对含水量、水势、渗透势、叶绿素、净光合速率、蒸腾速率、气孔导度、膨压的相对值做主成分分析, 分别得到3个主成分。干旱胁迫期间, 这3个主成分一共解释了92.41%的变量, 复水之后, 提取的这3个主成分解释了88.83%的变量。干旱期后第一个主成分PC1与相对含水量、水势相关, PC2与渗透势、净光合速率、蒸腾速率、气孔导度、膨压紧密相关, PC3与叶绿素含量紧密相关; 复水之后, PC1与渗透势、净光合速率、蒸腾速率、气孔导度紧密相关, PC2与相对含水量、水势紧密相关, PC3与叶绿素紧密相关。
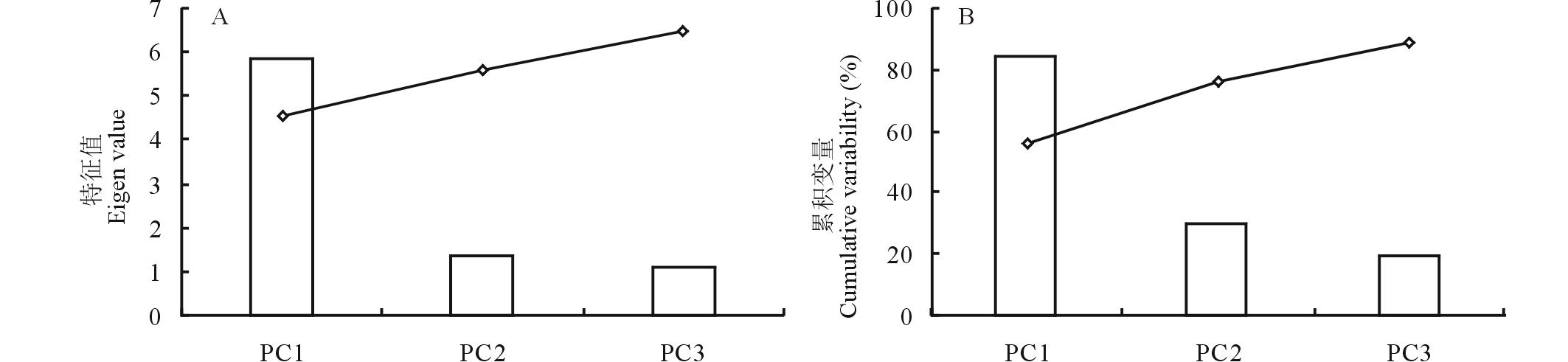 | 图8 干旱(A)及复水(B)时期各个主成分解释变量得分 柱状图代表每个主成分的特征值, 折线图代表每个主成分的累积变量。Fig.8 Scree plot of variance explained by each factor of the principal component during drought stress (A) and re-watering (B) Each bar represents corresponding eigenvalue of each principle component; line charts represent cumulative variability change. |
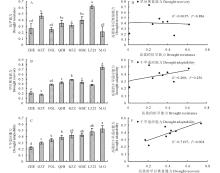
干旱适应能力不仅包括植物在干旱胁迫期间的抗旱能力, 也包括复水之后的恢复能力。由图9可看出, 不同品种的干旱适应能力为Moench.cv.Gadambalia(M.G)> 辽杂21(LZ21)> 高八尺(GBC)> 甘蔗籽(GZZ)> 千斤红(QJH)> 饭高粱(FGL)> 甘芝甜(GZT)> 遭烘萼(ZHE)。“ Moench.cv.Gadambalia(M.G)” 的旱后恢复能力最强, 抗旱能力最弱; “ 辽杂21(LZ21)” 的抗旱能力最强; “ 甘芝甜(GZT)” 的旱后恢复能力最弱。由抗旱能力、旱后复水的恢复能力和干旱适应能力三者之间的相关分析可得出, 旱后恢复能力和抗旱能力之间相关系数不显著, 说明彼此之间相互独立; 和抗旱能力相比, 旱后复水的恢复能力与干旱适应性的相关系数更高, 说明在植物适应干旱复水环境中, 旱后复水的恢复能力可能起着更重要的作用。
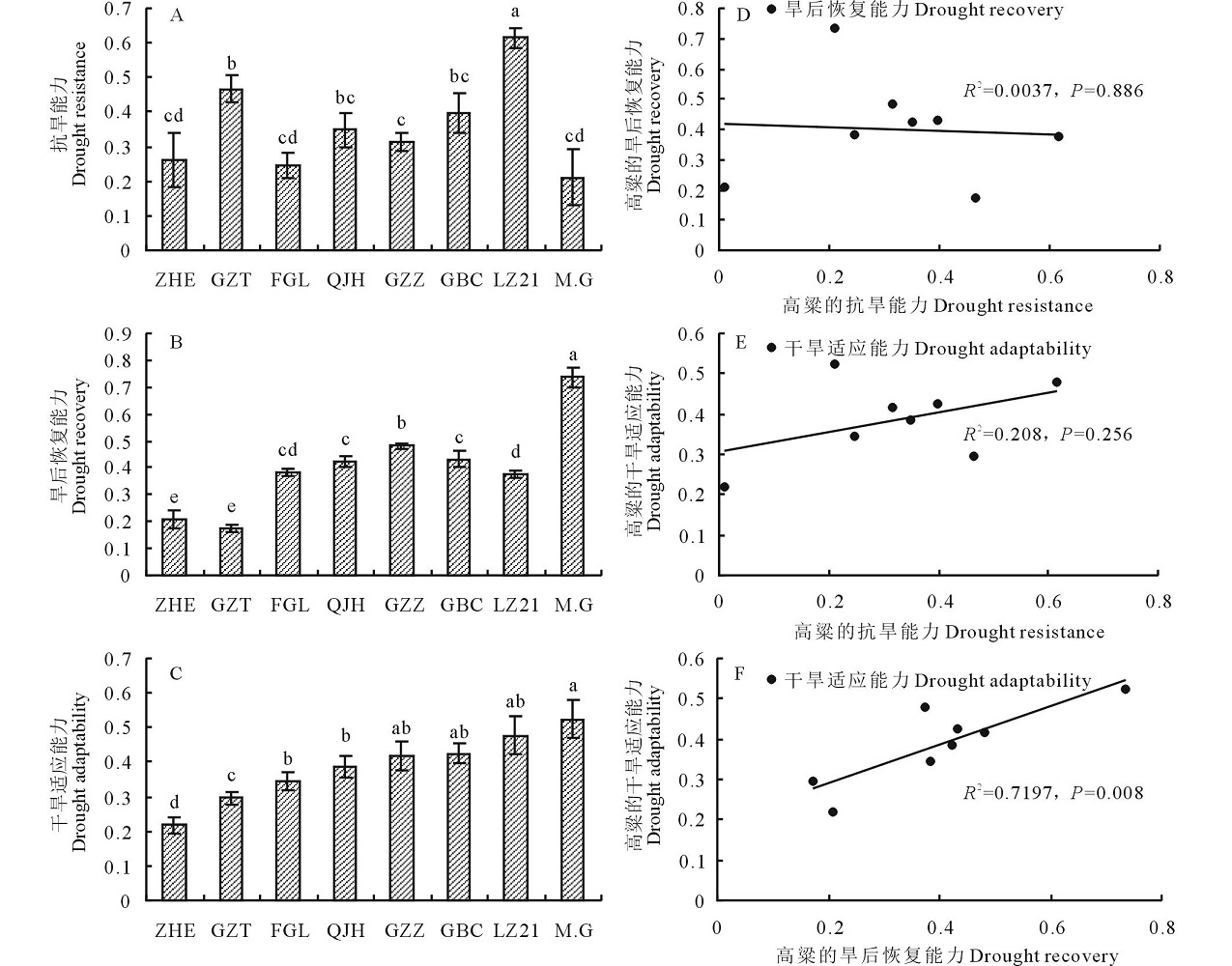 | 图9 不同高粱品种的抗旱能力(A)、复水恢复能力(B)、干旱适应能力(C)的比较以及三者之间的相关分析Fig.9 Drought-adaptive capabilities and correlations between drought-adaptive capabilities A: Drought resistance. B: Drought recovery. C: Drought adaptability. D: Correlation between drought resistance and drought recovery. E: Correlation between drought resistance and drought adaptability. F: Correlation between drought recovery and drought adaptability. |
如图10所示, 取各生理指标相对值的对数值(以10为底)做热图总结了8个高粱品种对抗旱性、旱后复水恢复性的整体生理响应。利用双变量Pearson相关系数法对干旱胁迫时期和复水恢复时期高粱的8个生理指标的相对值分别与抗旱能力、旱后恢复能力、干旱适应能力进行相关性分析, 结果如表1所示。干旱胁迫条件下, 净光合速率与抗旱能力呈显著正相关, 蒸腾速率、气孔导度与抗旱能力呈负相关, 叶片相对含水量与旱后恢复能力、干旱适应能力呈正相关, 净光合速率、蒸腾速率、气孔导度、水势与干旱适应能力呈负相关。水分胁迫解除之后, 水势与抗旱性显著负相关, 渗透势与旱后恢复能力、干旱适应性呈正相关。
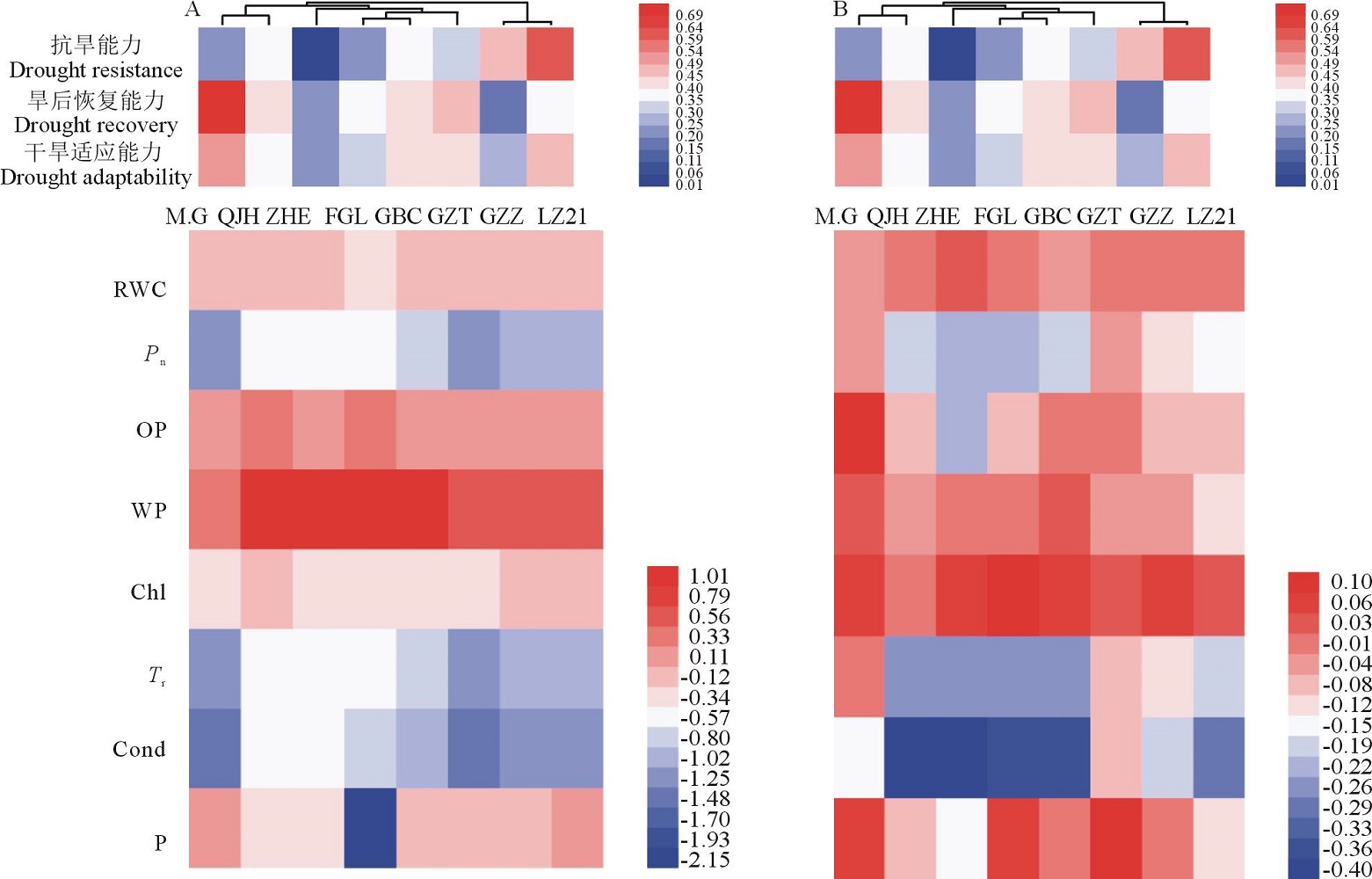 | 图10 不同品种高粱各生理指标响应干旱(A)及复水(B)的热图分析(图中数值采用相对值的对数值表示)Fig.10 Overall assessments of physiological responses in different sorghum lines during drought stress(A)and re-watering(B) |
| 表1 干旱及复水时期各生理指标相对值与高粱抗旱能力、复水恢复能力、综合抗旱性之间的相关性 Table 1 Correlations between drought-adaptive capabilities and physiological responses |
旱区作物的生长往往处在可变低水环境, 因此干旱环境下作物最终的产量不仅仅受其在遭受干旱胁迫过程中对干旱的抵御和耐受能力的影响, 还应包括水分条件改善之后作物从先前干旱损伤当中重新恢复正常生长的能力[16]。除抗旱能力外, 近些年, 更多研究开始关注当胁迫解除之后作物从胁迫损伤中恢复正常生长的能力, 即作物的旱后复水恢复能力[17]。因此, 衡量作物对于干旱的响应应该包括干旱和复水这两个过程。本研究采用Chen等[7]提出的干旱适应性定义来研究不同高粱品种对干旱胁迫的响应, 结果表明, 不同品种的干旱适应能力、抗旱能力和旱后复水的恢复能力均存在较大差异; 干旱期间抗旱能力最强的高粱品种, 在复水之后恢复生长的能力不一定最强, 而干旱期间抗旱能力弱的高粱, 在复水之后恢复生长的能力不一定也较弱。相关分析表明, 高粱的干旱适应能力与旱后复水的恢复能力呈显著正相关(r=0.84), 而与抗旱能力相关系数为0.45。说明采用干旱适应能力能更好地反映高粱对干旱胁迫的响应, 而旱后复水的恢复能力可能在高粱对干旱适应能力中起着更重要的作用, 如本研究中, Moench.cv.Gadambalia高粱的旱后恢复能力最强, 抗旱能力较差, 但在8个品种中却有着最好的干旱适应能力。此研究结果与Chen等[7]在玉米种的研究结果一致, 即玉米的干旱适应能力和抗旱能力、旱后复水恢复能力都有关, 而旱后复水的恢复能力可能扮演着更重要的作用。前人有研究表明, 作物的旱后复水恢复能力与抗旱性有关, 抗旱性强的品种, 同时复水后的恢复能力也较高[18, 19]。但是在本研究中抗旱能力和旱后复水的恢复能力无相关性, 说明高粱的抗旱能力和复水后的恢复能力是彼此独立的, 都是高粱响应干旱胁迫的重要途径。
干旱胁迫除了能显著造成生长抑制和产量下降等形态变化外, 也会对植物的生理、代谢等过程造成显著地影响, 这些生理指标的变化, 一方面是反映干旱抑制植物生长的内在生理机制, 同时也反映对干旱及复水响应的内在机制[12, 13, 20, 21]。因此, 探索干旱适应能力、抗旱能力及旱后复水的恢复能力和相关生理变化响应的关系, 一方面可以理解植物适应干旱的内在机制, 也可以为抗旱育种提供参考。本实验中, 干旱胁迫后各项生理指标都有所降低, 但降低幅度有所差异, 复水之后, 叶绿素含量、水势、渗透势恢复到甚至超过对照水平, 净光合速率、蒸腾速率、气孔导度有所恢复, 但大多数品种未恢复到对照水平。各项生理指标在干旱胁迫期间降低的程度, 以及复水后的恢复程度与品种有着很大关系。干旱期间维持较高的净光合速率和相对含水量有助于提高高粱的旱后恢复能力; 复水后的渗透势与旱后复水的恢复能力密切相关。而整体来看, 叶片相对含水量、渗透势等和干旱适应性密切相关。如“ Moench.cv.Gadambalia” 和“ 甘蔗籽” 高粱在干旱胁迫期间叶片相对含水量下降程度是最小的, 干旱适应能力最好。以上结果说明高粱在干旱胁迫期间抗旱性的生理基础与复水后恢复期间的复水恢复性的生理基础是不同的, 减轻高粱干旱胁迫期间的水分散失, 维持叶片水分状况, 有助于提高抗旱能力, 而维持在干旱胁迫下的良好水分状况有助于提高旱后复水的恢复能力。当水分胁迫缓解之后, 作物快速从干旱损伤中恢复的能力对于作物的生长发育有着积极的意义。因此, 复水恢复性在作物综合抗旱能力中的作用凸显的越来越重要[22]。
本研究表明高粱的干旱适应能力与抗旱能力和旱后复水的恢复能力都相关, 但旱后复水的恢复能力可能在高粱对干旱的适应能力中起着更重要的作用。而高粱在干旱胁迫期间的抗旱能力和复水后的恢复能力有着不同的生理基础, 因此, 在旱地作物品种选择和旱地育种中, 既要考虑其干旱胁迫下的抗旱能力, 又要考虑其旱后复水的恢复能力。本研究中, 由于是在苗期进行, 其结果主要适用于营养生长, 而和产量形成密切相关的生殖生长过程还缺乏相应的研究。但是在实际旱地作物生长中, 已被证明的干旱复水后产量的补偿和超补偿效应存在, 说明旱后复水的恢复能力及干旱适应性对旱地农业生产起重要作用。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|