{kind=link}
不同诱变处理对苜蓿叶片细胞显微和超微结构的影响
[冯鹏1, 2  , 孙力
, 孙力2 , 申晓慧1, 2 , 李如来2 , 李增杰2 , 李志民2 , 郑海燕2 , 姜成3 , 杨鹤2 , 刘俊刚2 , 郭伟2 , 张英俊4, * ]
, 孙力]
|
|


作者简介:冯鹏(1980-),男,内蒙古化德人,副研究员,博士。E-mail:fenggrass@163.com
研究60Co-γ射线、紫外线、零磁空间和EMS诱变处理下苜蓿SP1代叶片细胞显微、超微结构的变化,筛选最优诱变处理,选育适合高寒地区种植的苜蓿新品系或品种。以公农1号、Wega7F、WL319HQ、敖汉4个苜蓿品种试验材料,设置60Co-γ射线150、300、450 Gy射线强度,紫外线30、60、90 min照射时间,甲基磺酸乙酯(EMS) 0.1%、0.2%、0.4% (v/v) 溶液浓度,零磁空间处理180 d;测定苜蓿叶片细胞显微和超微结构。不同诱变处理后4个苜蓿品种叶片厚度均不同程度增加,其中公农1号EMS 0.4% (v/v)浓度下,叶片厚度最大,为446.16 μm,增厚幅度达到25.86%,同时诱变降低了叶脉突起程度;公农1号60CO-γ辐射和紫外线处理细胞结构疏松度均高于对照,诱变后Wega7F细胞结构紧密度升高;EMS处理下海绵组织厚度均降低;低剂量诱变处理下[150 Gy60Co-γ,30 min紫外线,0.1% (v/v) EMS],细胞叶绿体出现变形、基粒片层松散、类囊体解体、脂质球增多等现象,高剂量处理[450 Gy60Co-γ,90 min紫外线,0.4% (v/v) EMS]和零磁空间处理下,叶绿体膜会有部分模糊或解体,基粒片层膨胀或模糊、基粒垛叠程度变化随诱变处理种类增加或降低。4种苜蓿品种的叶片厚度诱变后叶片厚度均增加。在一定梯度范围内,60CO-γ射线和EMS诱变效率较高,突变体材料丰富,操作简单,适宜诱变育种的实际操作;150 Gy,60Co-γ射线、60 min紫外线、0.4%浓度EMS处理为苜蓿正向变异处理,有利于筛选有益诱变突变体。
, SUN Li
The overall aim of this research was to identify the optimal mutation method(s) for breeding new alfalfa varieties to cultivate in alpine regions. In this study, we investigated the effects several mutagenesis treatments [60COγ-ray irradiation, ultraviolet radiation, ethyl methyl sulfonate (EMS), and magnetic field-free space] on the microstructure and ultrastructure of the leaves of four alfalfa cultivars; Gongnong 1, Wega7F, WL319HQ, and Aohan. The mutagenesis treatments were as follows:60COγ-ray irradiation (150, 300, 450 Gy), ultraviolet radiation (30, 60, 90 min), ethyl methyl sulfonate [0.1%, 0.2%, 0.4% (v/v)], and magnetic field-free space (180 d). The microstructure and ultrastructure of the leaf were analyzed after these treatments. All of the mutagenesis treatments increased leaf thickness. The greatest increase was in Gongnong 1 under the 0.4% EMS (v/v) treatment, where the final leaf thickness was increased by 25.86 μm to 446.16 μm. The degree of leaf vein protuberance was decreased by the mutagenesis treatments. The degree of cell tightness in Gongnong 1 increased after60COγ-ray irradiation and ultraviolet radiation treatments. The cells in the leaves of wega7F were closer together after all the mutagenic treatments. The ultramicrostructure of the chloroplasts in the alfalfa leaves was also observed under different mutagenic treatments. Under lower doses or concentrations of mutagenic treatments [150 Gy60Co-γ, 30 min ultraviolet radiation, 0.1% (v/v) EMS], the chloroplasts were distorted with disrupted membranes, loose grana lamellae, disintegrated thylakoids, and increased numbers of lipid droplets. Under the highest doses or concentrations of mutagenic treatments [450 Gy60Co-γ, 90 min ultraviolet radiation, 0.4% (v/v) EMS, and 180 d magnetic field-free space] the chloroplasts became swollen, distorted, and severely damaged with fractured grana lamellae and disintegrated membrane systems. The numbers of plastids and starch grains in cells were increased under these treatments. Compared with the other treatments, the60COγ-ray irradiation and EMS treatments showed higher mutagenic efficiency, produced more mutant materials, and were simpler to operate. Therefore, we concluded that these were the best mutation methods. The 150 Gy60COγ-ray irradiation, 60 min ultraviolet radiation, and 0.4% EMS treatments were the mutagenic treatments that were most conducive to screening mutants.
叶片是植物体进行光合作用的营养器官, 同时也是对环境变化最为敏感的器官, 其形态结构特征最能体现环境因子的影响或植物对环境的适应[1]。结构是功能的基础, 植物生理功能的改变是由植物叶片结构的变化所致[2]。研究植物叶片形态解剖结构是对外界环境的适应性的反应, 叶片形态解剖结构对环境的响应机制是探索植物适应环境对策的基础。
诱变育种(Mutation breeding)是利用物理、化学和生物等因素诱导植物遗传性发生各种变异, 并从变异群体中选择符合育种目标的个体, 进而育成新品种或新种质的育种方法[3]。高等植物在不同逆境胁迫下, 细胞的结构也会发生不同程度的改变, 这一改变与植物所受胁迫程度和本身的抗逆性关系密切, 诱变突变体为育种提供选择材料。张文娟等[4]研究指出, 空间诱变后德福、德宝、阿尔冈金、三得利4个苜蓿品种叶片厚度增加, 叶脉突起度降低, 栅栏组织显著增厚; 与对照相比, 德福的细胞结构紧密度、细胞结构疏松度与对照差异均不显著, 阿尔冈金的细胞结构疏松度与对照差异不显著, 其他品种细胞结构紧密度、疏松度与对照均有显著差异。冯鹏等[5]研究卫星搭载对苜蓿超微结构变化的影响, 结果表明, 空间飞行后植株叶片细胞超微结构均出现了不同程度的变化, 对叶绿体的影响较大, 线粒体也出现较为明显的空洞和溢裂现象; 夏曾润等[6]通过不同浓度的甲基磺酸乙酯溶液诱变处理紫花苜蓿种子, 结果表明, EMS处理抑制了M1代种子的最终发芽率、发芽指数和胚根胚芽长度, 进而影响植株叶色, 原因可能是叶片叶绿体结构发生变化。张蕴薇等[7]发现空间诱变处理红豆草(Onobrychis viciaefolia)突变体细胞壁不规则增厚、扭曲, 细胞质稀薄, 液泡大, 叶绿体变小, 形状多不规则。叶绿体内淀粉粒细小、数量多, 基粒数减少, 每个基粒内部的基粒片层直径变小。
尽管零磁空间、60CO-γ 射线、紫外线以及甲基磺酸乙酯处理对紫花苜蓿诱变的研究已有报道, 但有关细胞显微和超微结构的报道仍较少。通过分析诱变处理后细胞显、超微结构的变化, 旨在探明不同诱变处理对苜蓿叶片细胞及亚细胞结构的影响, 为筛选最优诱变处理方法提供科学依据, 选育适合高寒地区种植的苜蓿品种。
本试验于2015年10月-2017年6月在黑龙江省农业科学院佳木斯分院和沈阳农业大学农学院实验室进行。试验地是黑龙江省农业科学院佳木斯分院牧草试验地。所在区域年平均降水量510 mm; 土壤类型为草甸黑土, pH 为6.10, 土壤有机质为2.65%、速效钾为112.76 mg· kg-1、速效磷为44.50 mg· kg-1、碱解氮为135.41 mg· kg-1、全磷为0.14%、全氮为0.19%, 全钾为1.60%。
选取公农1号、Wega7F、WL319HQ、敖汉4个苜蓿品种, 其中WL319HQ苜蓿是由北京正道生态科技有限公司提供, Wega7F、公农1号、敖汉苜蓿种子由黑龙江省畜牧研究所提供。
1.3.1 零磁空间 零磁空间采用双层磁屏蔽结构和线圈补偿方式相结合的大型26面体磁屏蔽装置, 直径为2.3 m, 磁强 ≤ 20 nT, 为地球磁强度的4× 10-4倍。将4个苜蓿品种风干种子各10 g, 3次重复, 在室温下经零磁空间处理180 d。
1.3.2 60CO-γ 射线 4个苜蓿品种种子进行60CO-γ 射线辐照处理, 剂量分别为150、300、450 Gy, 以0剂量处理作为对照, 每个处理300粒, 处理周期为7 d, 3次重复。
1.3.3 紫外线 选取籽粒饱满的苜蓿种子300粒, 将其包于纱布中, 用45 ℃的蒸馏水浸泡1 h, 用滤纸吸干种子表面的水分, 再用0.1%的氯化汞溶液消毒 10 min。无菌水清洗 3~5次, 将其置于紫外灯下40 cm处照射30、90 min。3次重复。
1.3.4 甲基磺酸乙酯(EMS) 挑选成熟的苜蓿种子900粒, 用浓硫酸处理5 min, 蒸馏水冲洗多次。在4 ℃磷酸缓冲液 (100 mmol· L-1, pH 7.0)浸泡12 h使种子完全吸水膨胀。用磷酸缓冲液将EMS分别配制成浓度为0.1%、0.4% (v/v)的溶液, 置于室温黑暗条件下处理种子15 h, 期间对药剂浸泡的种子进行轻轻翻转摇动, 然后用蒸馏水反复冲洗, 去除残留种子表面的EMS溶液。
1.4.1 显微制片及观察 选取诱变当代健康植株中部功能叶片, 叶片沿中脉切取大小为5 mm× 5 mm, 用 FAA 固定液固定, 常规石蜡切片法制片, 切片厚度 5 μ m, 番红-固绿双重染色。Nikon-DIAPHOT显微镜上观察。
1.4.2 电镜制片及观察 取诱变处理和未经处理的苜蓿中部功能叶片, 注意避开叶脉, 切成长 1 cm、宽约0.5 cm 左右的小块, 迅速放入浓度 2.5%、pH 6.8 的戊二醛固定液中, 同时抽真空使材料沉入固定液, 在4 ℃下固定2 h后用 0.1 mol· L-1磷酸缓冲液(pH=6.8)冲洗2次, 每次间隔时间为2 h, 随后采用浓度为50%、70%、80%、95%、100%的乙醇进行依次脱水, 脱水后转移至100%的丙酮内, 最后用 Epon812包埋剂浸透包埋。利用 ULTRACUTE 型超薄切片机将包埋块切成50~70 nm的薄片, 醋酸双氧铀-柠檬酸铅双重染色, HITACI-II~HT7700 型透射电镜下观察, 30000倍镜下照相。
采用 Motic Image 2000 1.3软件进行显微测量, 测量叶片厚度、海绵组织厚度、栅栏组织厚度、叶脉厚度, 各显微结构观测值均为30个数值的平均值, 并计算:
细胞结构紧密度 CTR(cell tense ratio)=栅栏组织厚度/叶片厚度× 100%
细胞结构疏松度SR(spongy ratio)=海绵组织厚度/叶片厚度× 100%
叶脉突起度VPD(vein protuberant degree)=叶脉厚度/叶片厚度
2.1.1 叶片和叶脉突起度 不同诱变处理对苜蓿当代植株叶片显微结构产生不同程度的影响, 变异方向及变异程度因方法和诱变剂量不同而异[8, 9]。4种苜蓿品种的叶片厚度在诱变后变化不同, 但主要以叶片厚度增加为主。公农1号除30 min紫外线和0.1% EMS处理下叶片厚度低于对照, 其他处理均显著高于对照 (P< 0.05) (表1); Wega7F除EMS 0.1%处理下叶片厚度低于对照, 其他处理也均显著高于对照(表2); WL319HQ和敖汉苜蓿在所有处理下叶片厚度全部显著高于对照(表3, 4)。各苜蓿品种诱变处理后, 叶脉突起程度均显著小于对照(P< 0.05)(表1~4)。
| 表1 诱变处理对公农1号叶片主要结构特征影响 Table 1 Comparison of main structural features of Gongnong 1 under different mutagenic treatments (μ m) |
| 表2 诱变处理对Wega7F叶片主要结构特征影响 Table 2 Comparison of main structural features of Wega7F under different mutagenic treatments (μ m) |
| 表3 诱变处理对WL319HQ叶片主要结构特征影响 Table 3 Comparison of main structural features of WL319HQ under different mutagenic treatments (μ m) |
| 表4 诱变处理对敖汉苜蓿叶片主要结构特征影响 Table 4 Comparison of main structural features of Aohan under different mutagenic treatments (μ m) |
2.1.2 栅栏组织和海绵组织 叶片的厚度主要取决于栅栏组织厚度和海绵组织厚度[10]。叶片厚度随栅栏组织厚度增加而增加, 即栅栏组织越厚, 叶片厚度越厚。不同苜蓿品种受不同诱变处理后, 栅栏组织的厚度随着叶片厚度变化而变化[11]。与对照相比, 公农1号60CO-γ 辐射、紫外线和EMS高剂量下均高于对照处理, 同一诱变处理随着剂量的增大, 栅栏组织厚度增加, 其他处理下均低于对照(表1); 海绵组织在紫外线30 min和0.1% (v/v)EMS处理及零磁空间处理下显著低于对照(P< 0.05), 其他处理均高于对照(表1); Wega7F的栅栏组织均显著高于对照, 海绵组织多数处理条件下也显著高于对照, 但EMS处理下海绵组织厚度均低于对照(表2); WL319HQ苜蓿的栅栏组织均显著高于对照(P< 0.05)(表3)。
2.1.3 细胞结构紧密度(CTR)和细胞结构疏松度(SR) 供试的4 个苜蓿品种叶片结构细胞结构紧密度、细胞结构疏松度变化各不相同。与对照相比, 公农1号苜蓿只在紫外线处理下CTR显著低于对照(P< 0.05), 其他处理与对照差异不显著(表1); 公农1号60CO-γ 辐射和紫外线处理SR均高于对照, 其他处理低于对照(表1); 诱变后Wega7F CTR升高; SR只是60CO-γ 的150 Gy辐射处理、紫外线处理和0.4% EMS处理高于对照; 敖汉苜蓿与对照相比, 除150 Gy处理的CTR显著高于对照外, 其他所有处理下CTR均低于对照处理, 只有EMS 0.1%处理CTR高于对照, 说明敖汉苜蓿诱变后细胞结构较为松散(表1~4)。
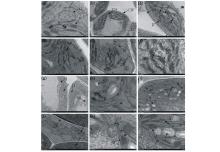
从图1可以看出, WL319HQ对照组叶肉细胞中的叶绿体靠近细胞壁整齐排列, 形状规则呈近纺锤形, 叶绿体膜完整无缺, 基粒片层结构排列整齐、均匀、紧密, 淀粉粒较小, 含量较少。在紫外线(30 min)和EMS[0.1% (v/v)]低剂量或浓度处理下, 叶肉细胞中的叶绿体形状仍较规则, 片层结构和垛叠结构排列整齐, 但部分垛叠结构开始模糊, 线粒体脊部分消失, 质体数量减少, 少量片层溶解, 但在150 Gy辐射处理下, 叶肉细胞超微结构除具有前面的其他处理低剂量的特征外, 此处理下的垛叠结构全部有肿胀现象。高剂量处理下[450 Gy 60Co-γ , 90 min紫外线, 0.4% (v/v) EMS], 多数细胞壁仍然完好, 但个别细胞壁完全溶解, 内容物全部外渗至细胞间隙, 叶绿体膜溶解, 个别叶绿体膜有向外突起现象, 质体明显增多, 片层结构逐渐溶解, 个别垛叠结构增厚, 部分有溶解模糊现象, 在完整细胞内, 胞浆向四周扩散, 淀粉粒增多, 多数线粒体脊消失。零磁空间处理下, 细胞内一些变化同高剂量处理相似, 但是细胞壁处理有完全溶解现象外, 细胞壁上还有分泌物质存在, 质体数量下降, 细胞充满内含物, 淀粉粒多, 且增大。
 | 图1 WL319HQ叶片细胞超微结构(× 30000) a: CK; b: 0.1% EMS处理; c: 30 min紫外线处理; d~e: 90 min紫外线处理; f: 150 Gy辐射处理; g: 300 Gy辐射处理; h~i: 450 Gy辐射处理; j~k: 0.4% EMS处理; l: 零磁空间处理。 CW: 细胞壁; VA: 液泡; CH: 叶绿体; M: 线粒体; S: 淀粉粒; D: 垛叠结构; P:质体。Fig.1 Ultrastructure of WL319HQ (× 30000) a: CK; b: 0.1% EMS treatment; c: 30 min UV treatment; d-e: 90 min UV treatment; f:150 Gy radiation treatment; g: 300 Gy radiation treatment; h-i: 450 Gy radiation treatment; j-k: 0.4% EMS treatment; l: magnetic field free space. CW: cell wall; VA: vacuole; CH: chloroplast; M: mitochondrion; S: starch grain; D: crib fold structure; P: plasmid. |
植物处于逆境下, 其形态学特征、生理生化特征以及细胞结构均产生变化, 细胞结构的改变既是基因变异的反映[12], 又是对环境的适应性的体现[13, 14]。所以通过对显微结构变化的研究分析可以探究其微观变异机理, 同时也可以通过对显微结构特征的了解来筛选出抗寒性强的诱变方法及有益突变体植株。通常情况下, 叶片厚度, 叶肉栅栏组织、海绵组织和叶脉等形态解剖结构变化多数是由于外部环境变化所致[15]。植物的栅栏组织厚度、海绵组织厚度、叶片组织结构紧密度、疏松度等细胞结构特性均与植物抗逆性有一定关系。由于叶肉细胞结构紧密程度主要是针对栅栏组织而言, 所以细胞结构紧密度用栅栏组织厚度占叶片厚度比例的相对值来体现, 栅栏组织也被用来作为抗寒指标进行研究[16]。Pavel等[17]认为细胞结构紧密程度越大, 该品种的抗寒能力越强。房用等[18]对茶树的细胞结构分析认为, 茶树细胞栅栏组织厚度、栅栏组织层次、栅栏组织结构排列紧密程度以及栅栏组织厚度与海绵组织厚度的比值与茶树的抗寒具有正相关关系。冯鹏[19]研究表明, 空间搭载后, 紫花苜蓿高水分含量叶片厚度增加, 海绵组织厚度和紧密程度也有所增加。张蕴微等[20]研究表明, 对于苜蓿叶片显微结构来说, 海绵组织和栅栏组织在空间诱变处理下反应效应不同, 空间诱变对海绵组织的影响大于对栅栏组织的影响, 空间搭载后, 苜蓿叶片海绵组织结构与对照达到差异显著水平, 但栅栏组织与对照差异不显著。本研究结果表明, 诱变后叶片厚度、叶脉突起度、栅栏组织厚度、海绵组织厚度和细胞组织结构紧密度等指标均会产生变异, 诱变方法和苜蓿品种不同, 变化程度也不同, 与对照相比, 供试苜蓿品种叶片厚度多数呈增加趋势, 而叶片细胞结构紧密度增加, 有利于增强其抵御逆境的能力和抗寒性的提高。公农1号苜蓿的栅栏组织厚度在450 Gy辐射诱变和90 min紫外线处理下增加, Wega7F、WL319HQ和敖汉苜蓿的栅栏组织与对照相比均呈增加趋势, 各品种的海绵组织厚度则随诱变处理的不同而增减不同, 细胞结构的紧密度与疏松度和海绵组织变化相似, 说明其变异方向及变异程度具有不确定性。本研究选取的4个苜蓿品种半同胞种子作为诱变材料, 极大地减少了种子间遗传背景的差异, 并选择叶片作为显微结构的研究对象, 可以更准确地反映植株对诱变条件的适应。
叶片对不同诱变反应都非常敏感[21]。叶片细胞显微结构和超微结构经过诱变后都会发生显著变化, 如细胞壁薄厚不均匀[22], 叶肉细胞内部分细胞器退化消失[23], 细胞液泡变大[24], 细胞器被挤到细胞壁边缘[25], 叶绿体变形或膜解体[26]。杨凤仙等[27]还观察到低温胁迫下叶绿体膜解体形似几个变态叶绿体相互粘连堆叠在一起, 大部分叶绿体的基质片层排列紊乱, 有些片层融合而变得模糊不清或完全消失。王冬梅等[28]研究表明, 番茄(Lycopersicon esculentum)叶片叶绿体在高温胁迫下会膨大变圆, 叶绿体被膜出现不同程度的断裂、解体, 类囊体片层松散, 排列紊乱。这是由于细胞受到伤害后做出的应激反应所致。本研究中, 与对照相比, 尤其是在不同诱变高剂量和零磁空间处理下, 叶绿体膜会有部分模糊或解体, 基粒片层膨胀、溶解、基粒垛叠程度下降, 个别也有增加, 有的细胞壁完全破裂, 内含物全部外泄, 线粒体脊模糊, 质体明显增多并出现聚集现象。由于质体是一类与碳水化合物合成与储藏有关的细胞器, 质体增加能够提高细胞活力, 对于提高苜蓿抗寒性具有优势作用[29, 30]。本研究还发现, 诱变处理后, 叶绿素中的个别淀粉粒变多变大, 说明光合产物淀粉在叶肉细胞叶绿体内的积累, 能够大大提高紫花苜蓿对寒冷的忍受能力, 因为增加的淀粉解体时能够释放能量, 这将更好的维持细胞较高的活性水平; 同时解体的淀粉粒还具有提高细胞质浓度、增加渗透压及吸水力以及提高其抗逆性的作用。
1)诱变处理增加了苜蓿叶片厚度, 但降低了叶脉突起程度; 细胞淀粉粒变大, 数量变多, 有助于提高苜蓿的抗寒性能。
2)在一定梯度范围内, 60CO-γ 射线和EMS诱变效率较高, 突变体材料丰富, 操作简单, 适宜于诱变育种的实际操作。
3)150 Gy 60Co-γ 射线、60 min紫外线、0.4%浓度EMS处理为苜蓿正向变异处理, 有利于筛选有益诱变突变体。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|