




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
异源表达 WZY2-1基因提高拟南芥植株抗旱性
[强治全1  , 杨文博
, 杨文博2 , 张帅1 , 于正阳1 , 史学英1 , 王鑫1 , 朱维宁3 , 张林生1, * ]
, 杨文博]
|
|


作者简介:强治全(1990-),男,甘肃庆阳人,在读硕士。E-mail:qiangsir0934@qq.com
脱水素是受细胞失水相关环境诱导表达的一类蛋白家族。所有的脱水素都含有保守的K片段,这段保守序列可以形成两亲性的α-螺旋,这种结构在脱水素发挥功能中起主要作用。小麦 WZY2-1基因含有9个K片段,属于K9型的脱水素,能够被低温、干旱和盐胁迫诱导表达。在本实验中,纯化了WZY2-1蛋白,通过体外LDH酶保护实验发现,WZY2-1蛋白具有保护LDH酶活性的功能。通过亚细胞定位分析发现,WZY2-1蛋白主要定位于细胞核和细胞膜。为了进一步说明 WZY2-1基因的功能,获得了转 WZY2-1基因的拟南芥植株,通过干旱胁迫条件下转基因植株的生理指标和表型分析,发现转 WZY2-1基因的拟南芥植株具有较强的抗旱能力。因此,推测小麦 WZY2-1基因在植物干旱胁迫条件下具有重要功能。
, YANG Wen-bo
Dehydrins (DHNs) belong to a protein family whose expression may be induced or enhanced by environmental stresses that lead to cell dehydration. All DHNs contain at least one motif, or K-fragment, which can form an amphipathic helix. This structure play an important role in DHN function. The wheat gene WZY2-1, a K9-type DHN gene, can be activated by low temperature, drought and salt stress. In this work, the WZY2-1 recombinant protein was purified. In vitro lactate dehydrogenase (LDH) assays revealed that WZY2-1 had a potent protective effect during freezing-thawing temperature stresses. Subcellular localization showed that WZY2-1 was localized in the nucleus and cell membrane. To further elucidate the function of WZY2-1 in plants, transgenic Arabidopsis plants that overexpress WZY2-1 were tested. The transgenic Arabidopsis lines showed a better tolerance to drought stress than unmodified. This suggests that the WZY2-1 gene may also play an important role in drought tolerance in wheat.
脱水素(dehydrins, DHNs)属于胚胎发育晚期丰富蛋白(late embryogenesis abundant protein, LEA)第二家族[1]。该类蛋白具有3个保守的区域:K、Y和S片段。其中, K片段(EKKGIMDKIKEKLPG)是脱水素共有的, 它可以形成兼性α -螺旋, 在植物失水情况下, α -螺旋的形成能够防止细胞内蛋白变形和细胞膜损伤[2]。此外, 有些脱水素K片段之间富含极性氨基酸, 称作Φ 片段, 它可能参与了细胞质组分的相互作用[3]。S片段由一段串联的丝氨酸(SSSSS)组成, 可以被蛋白激酶磷酸化, 与脱水素的入核有关[4]。Y片段的保守序列为(VTDEYGNP), 目前关于Y片段的功能尚不清楚。根据脱水素的K、Y和S片段组合, 将脱水素分成5个亚型:YnSKm、Kn、KnS、SKn和YnKm型[5]。
近些年, 在植物抗逆分子机理研究中, 脱水素已成为植物耐逆程度的分子指标。目前已经从玉米(Zea mays)[6]、大麦(Hordeum vulgare)[7]、高粱(Sorghum bicolor)[8]、水稻(Oryza sativa)[9]等多种农作物中克隆和分离出脱水素基因。脱水素的亚细胞定位多种多样, 但主要分布于细胞质和细胞核。通过对仙人掌(Opuntia stricta)脱水素OpsDHN1亚细胞定位, 发现OpsDHN1蛋白主要定位于细胞核和细胞质[10]。Yang等[11]研究发现小麦WZY2蛋白的亚细胞定位受外界环境条件的影响, 在正常情况下, 小麦(Triticum aestivum)WZY2蛋白主要分布在细胞核和细胞质, 但在低温处理下, 一些WZY2蛋白向细胞膜聚集。许多实验证明脱水素可以作为酶的保护剂, Drira等[12]对小麦DHN5及突变体研究表明, 低温胁迫下DHN5及突变体可以保护乳酸脱氢酶(lactate dehydrogenase, LDH)活性, 并且认为K片段可能起重要作用。与此类似的还有WZY2[11]、WDHN1[13]等脱水素。脱水素在植物抵御各种逆境胁迫中具有重要作用, Saibi等[14]将大麦的脱水素基因DHN5转化拟南芥(Arabidopsis thaliana), 获得的转基因株系具有较强的抗盐性; Liu等[1]将番茄(Lycopersicon esculentum)的脱水素基因ShDHN过表达, 增强了番茄对干旱、低温、盐等多种胁迫的耐受性; 相反的, Chen等[15]将辣椒(Capsicum annuum)的脱水素基因CaDHN1缺失, 获得了对低温、盐和渗透胁迫敏感性植株。
小麦WZY2-1基因全长1740 bp, 编码579个氨基酸, 分子量约60 kDa, 含有9个保守的K片段, 属于典型的Kn型脱水素。研究表明, WZY2-1基因受干旱、低温和盐渍胁迫诱导[16]。本研究对WZY2-1蛋白进行了亚细胞定位, 研究了温度胁迫下WZY2-1蛋白对LDH酶活性的影响, 并获得了转WZY2-1基因的拟南芥植株, 对转基因植株干旱耐逆性进行了分析。
供试拟南芥(生态型 Columbia)用70%乙醇表面灭菌1 min, 然后用10%的NaClO溶液消毒5 min, 用无菌水冲洗3次。将无菌的拟南芥种子铺在0.5 MS固体培养基上(pH 5.8), 4 ℃处理3 d, 移至光照培养箱[16 h 光照, 8 h 黑暗, (22± 1) ℃], 生长7 d后, 移栽至9 cm的栽培盆中, 于培养室[16 h 光照, 8 h 黑暗, (22± 1) ℃]中培养。
通过对pBI121载体改造, 转化大肠杆菌TOP10, 涂板, 37 ℃培养, 挑单克隆, 摇菌, 菌液PCR检测, 用Xho I和Sal I双酶切检测, 阳性克隆测序(上海生工生物工程有限公司)。提取构建的亚细胞定位载体pBI121-WZY2-1-GFP(天根公司), 转化农杆菌GV3301, 涂板, 28 ℃培养, 挑单克隆, 摇菌, 菌液PCR检测, 获得含有pBI121-WZY2-1-GFP载体的农杆菌。
将含有pBI121-WZY2-1-GFP农杆菌接种含有50 μ g· mL-1卡纳抗生素和利福平的YEB培养基中, 28 ℃培养至OD600到0.6, 2800 r· min-1离心10 min, 收集菌体, 用含有200 mmol· L-1的乙酰丁香酮、10 mmol· L-1的MgCl2和0.02%的Sillwet-77的0.5 MS液体培养基悬浮, 室温放置4 h, 作为浸染液。取新鲜的洋葱切开鳞茎, 用解剖刀挑取1 cm2的洋葱内表皮细胞, 置于0.5 MS固体培养基上, 28 ℃暗培养24 h。将预培养的洋葱表皮细胞在浸染液中浸泡30 min, 滤干菌液, 平放于0.5 MS固体培养基上, 置于光照培养箱(16 h 光照, 8 h 黑暗, 25 ℃)培养2 d。取培养2 d的洋葱表皮细胞, 用1 mg· mL-1的DAPI溶液染色5 min, 制片, 荧光显微镜观察。
取20 μ L将乳酸脱氢酶(lactate dehydrogenase, LDH)(EC1.1.1.27, Sigma)分别与WZY2-1蛋白、BSA等体积混合, 对混合样品进行不同胁迫处理。冷融胁迫:将样品用液氮速冻30 s, 25 ℃水浴解冻5 min。45 ℃处理:将样品经45 ℃处理30 min。取40 μ L胁迫处理的样品加入到710 μ L的反应体系[10 mmol· L-1 Na3PO4, pH 7.4, 10 mmol· L-1丙酮酸, 0.2 mmol· L-1 还原性辅酶I(nicotinamide adenine dinucleotide, NADH)]中, 用SpectraMax M2酶标仪测定A340吸光值的减少值。测定程序:25 ℃水浴30 s, 每隔15 s测定1次, 持续测定5 min。以未处理的LDH酶活性为100%, 实验重复3次。LDH酶残留活性计算:LDH酶残留活性=(处理Δ 340/未处理Δ 340)× 100%。
设计含有Xba I和Sac I酶切位点的引物(TGF: 5'-GCTCTAGAATGCACGACGCCGAC-3'; TGR: 5'-CGAGCTCTTAGTTCAGTCCAGGC-3'), PCR扩增WZY2-1基因的cDNA全长序列, 回收PCR产物。将PCR产物连接至pBI121载体上, 转化大肠杆菌TOP10, 涂板, 37 ℃培养, 挑单克隆, 摇菌, 菌液PCR检测, 阳性克隆测序(上海生工生物工程有限公司)。将过表达载体pBI121-WZY2-1转化农杆菌GV3301, 28 ℃培养, 挑单克隆, 摇菌, 菌液PCR检测, 获得含pBI121-WZY2-1载体的重组菌。利用花絮浸染法转化拟南芥, 收取T0代种子。
将收取的T0代种子铺在含有30 μ g· mL-1卡纳抗生素的0.5 MS培养基(pH 5.8)上培养, 将绿色幼苗转移到营养土培养20 d, 提取叶片基因组DNA(天根公司)。上游引物包含pBI121载体本身序列, 下游引物位于WZY2-1基因序列内部, 引物序列JCF: 5'-AGCTGCCTGGACTGAACT-3'; JCR: 5'-GCCTCTTCGCTATTACGCCAGGC-3'。以提取的T1代拟南芥基因组DNA为模板, 进行PCR检测, 收取阳性植株种子, 继续培养筛选, 直到获得T3代纯合转基因株系种子。
将消毒的野生型拟南芥和3个株系的T3代过表达WZY2-1的转基因纯合体种子铺种在0.5 MS培养基上, 4 ℃培养3 d后, 于光照培养箱培养2周。按照TRIzol试剂(Invitrogen)的操作说明, 提取野生型和T4代转基因植株幼苗总RNA。以提取的RNA为模板, 按照反转录试剂盒PrimeScript RT reagent Kit with gDNA Eraser(TaKaRa)的操作说明反转录成cDNA。以拟南芥的Actin2基因为内参, 引物序列ActF: 5'-TATCGCTGACCGTATGAG-3'; ActR: 5'-CTGAGGGAAGCAAGAATG-3', 用WZY2-1基因特异性引物(WF: 5'-CGGAGTGACCGATAAGG-3'; WR: 5'-TGCCAGTTGTTTCGTTGT-3')进行RT-PCR分析, PCR反应体系25 μ L, 包括2× SYBR Premix 12.5 μ L, 模板2 μ L, 上、下游引物各1 μ L。反应程序为95 ℃预变性30 s, 然后95 ℃ 5 s, 55 ℃ 20 s, 39个循环。
取同一时间种植和收获的野生型拟南芥和T3代转WZY2-1的拟南芥3个株系种子在4 ℃放置3 d, 种子消毒后, 铺于0.5 MS培养基上培养, 萌发后移栽至小花盆, 转移到培养室培养。正常条件生长20 d后, 脱水处理14 d后, 观察野生型和转基因植株表型。
将生长20 d的野生型和T3代转WZY2-1基因的拟南芥, 干旱胁迫36 h, 取成熟叶片, 测定其生理指标。试验方法见文献[14, 17-18]。采用酸性茚三酮法测定脯氨酸含量; 采用分光光度法测定叶绿素和MDA的含量; 相对含水量按照公式:相对含水量=[(鲜重-干重)/鲜重]× 100%, 实验重复3次。
对pBI121载体进行改造, 获得pBI121-WZY2-1-GFP融合蛋白表达载体, 转化至大肠杆菌感受态细胞TOP10中。对阳性克隆测序, 测序结果表明, 亚细胞定位载体构建成功, 命名pBI121-WZY2-1-GFP(图1A)。
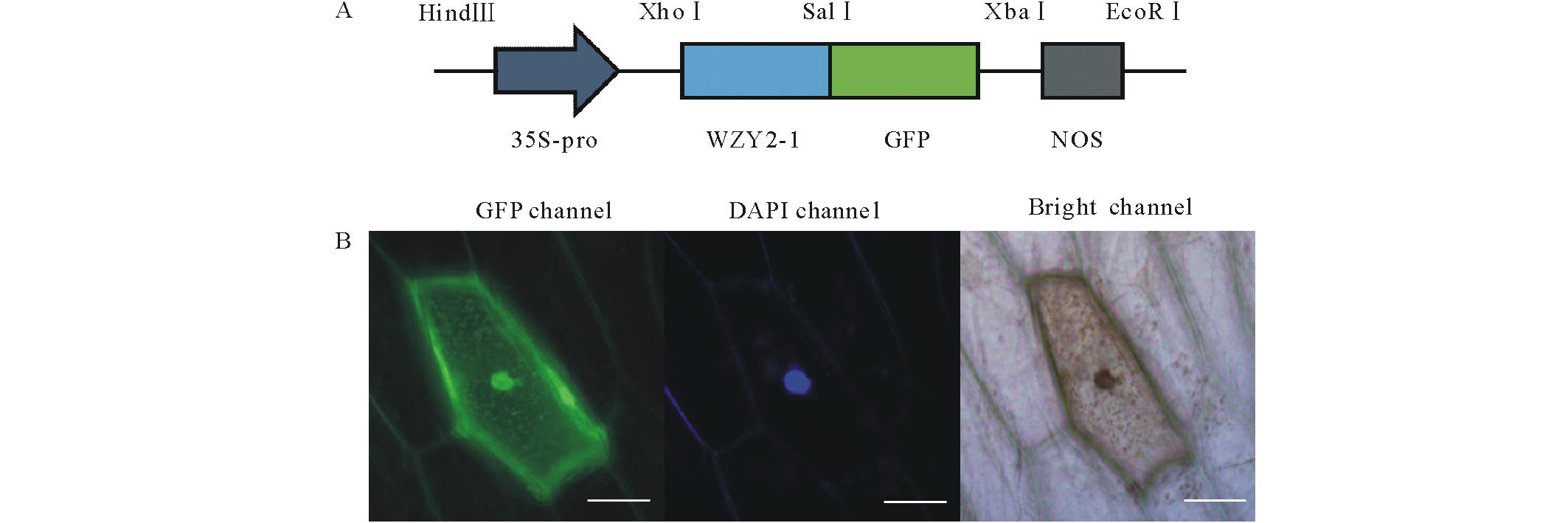 | 图1 WZY2-1蛋白的亚细胞定位 (A) 亚细胞定位载体示意图。35S-pro代表花椰菜花叶病毒的启动子; WZY2-1代表WZY2-1蛋白编码区去掉终止子; GFP代表绿色荧光蛋白; NOS代表胭脂碱合成酶基因终止序列。(B)WZY2-1融合蛋白在洋葱表皮细胞的荧光显微观测图(Bar=100 μ m)。GFP通道、DAPI通道(细胞核)和明场都进行展示。Fig.1 Subcellular localization of WZY2-1 protein (A) Diagram of the constructs of pBI121-WZY2-1-GFP used for subcellular localization. 35S-Pro, Cauliflower mosaic virus 35S promoter; WZY2-1, the WZY2-1 coding region without the stop codon; GFP, green fluorescent protein; NOS, nopaline synthase terminator. (B) Fluorescent microscopic images of WZY2-1-GFP fusion protein in onion epidermal cells (Bar=100 μ m). The GFP channel, DAPI channel (nucleus) and bright-field images were showed. |
将pBI121-WZY2-1-GFP载体转化农杆菌GV3301, 菌液PCR获得阳性重组菌。通过农杆菌浸染洋葱表皮细胞, WZY2-1蛋白在整个细胞中的亚细胞位置如图1B所示。通过对荧光信号分析, 推测WZY2-1蛋白主要分布于细胞核和细胞膜。
为了研究WZY2-1蛋白对LDH酶的保护作用, 将 LDH酶分别与WZY2-1蛋白、BSA等体积混合, 对混合样品进行不同胁迫处理后, 测定LDH酶活性(图2)。结果表明, 冻融处理后, Na3PO4缓冲液LDH几乎没有保护作用, BSA和WZY2-1蛋白分别能够维持70%和75%的LDH酶活性; 45 ℃处理后, Na3PO4缓冲液同样没有保护作用, BSA和WZY2-1蛋白分别能够维持71%和58%的LDH酶活性。说明WZY2-1蛋白在低温和高温下具有显著的LDH酶的保护作用。
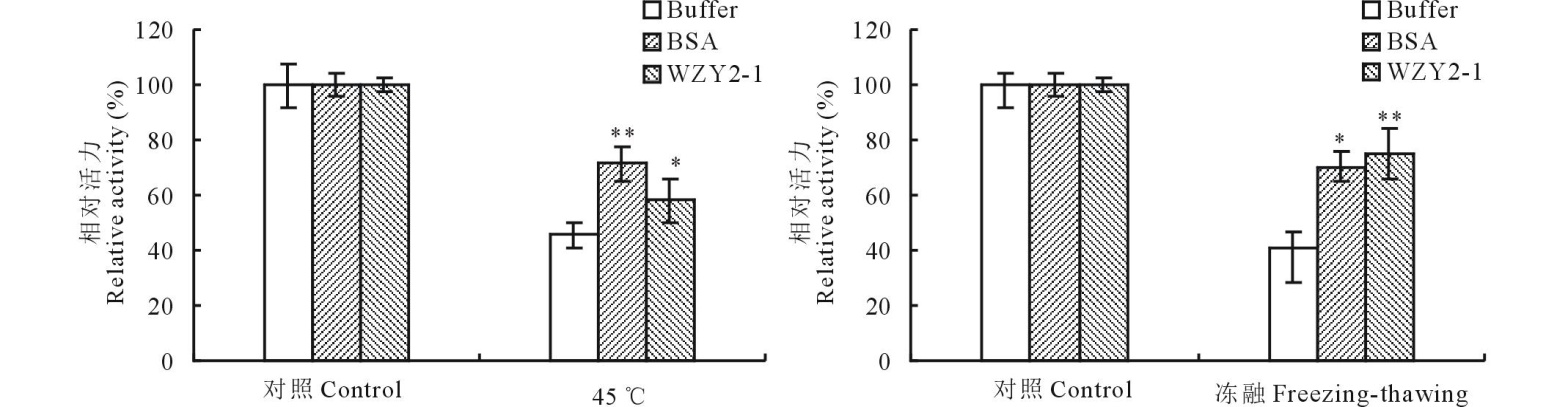 | 图2 冷融及45 ℃胁迫下WZY2-1蛋白对LDH酶活力的保护 柱状图上星号表示Buffer和BSA、WZY2-1对LDH酶活性保护的差异显著性 (* , P< 0.05; * * , P< 0.01)。Fig.2 WZY2-1 protected LDH activity from freezing-thawing and 45 ℃ stresses in vitro Different asterisk on top of bars indicate the significant differences of the LDH activity between Buffer and BSA or WZY2-1 protein (* , P< 0.05; * * , P< 0.01). |
为了进一步研究WZY2-1在植物中的功能, 构建pBI121-WZY2-1转基因载体。通过花絮浸染法获得了T0代转基因拟南芥种子。通过卡纳抗生素筛选和基因组DNA的PCR检测, 获得T1代阳性植株, 收取T1代种子。继续筛选, 最终获得了T3代纯合转基因株系种子。
提取3个株系T3代转基因纯合株系的RNA, 通过实时荧光定量PCR分析WZY2-1基因的表达情况。结果如图3所示, 表明WZY2-1基因能够在转基因株系中过表达。
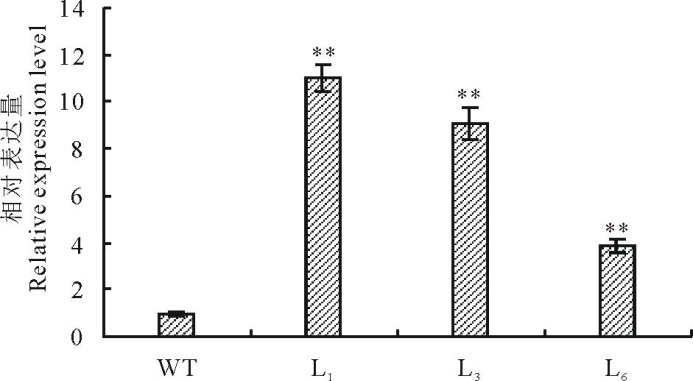 | 图3 WZY2-1基因在野生型和转基因拟南芥表达量 柱状图上星号表示野生型(WT)和转基因(L1、L3、L6)植株WZY2-1基因表达水平的差异显著性 (* , P< 0.05; * * , P< 0.01)。Fig.3 Relative expression level of WZY2-1 in the wild-type and the transgenic lines Different asterisk on top of bars indicate the significant differences of the relative expression level between wild-type (WT) and transgenic A. thaliana(L1, L3 and L6) (* , P< 0.05; * * , P< 0.01). |
将正常生长20 d的拟南芥植株脱水处理14 d后, 表型观测如图4所示, 从图看出转基因植株叶片萎蔫面积比野生型植株少, 并且转基因植株的叶片也比野生型大。说明异源过表达小麦WZY2-1基因可以提高拟南芥植株干旱胁迫的耐受性。
 | 图4 干旱胁迫下野生型和转基因拟南芥叶片萎蔫程度Fig.4 The leaves wilting degree of wild-type and transgenic A. thaliana under dehydration stress |
实验测定胁迫处理36 h后转WZY2-1基因和野生型拟南芥的相对含水量、脯氨酸、叶绿素和MDA含量(图5)。从结果可知, 正常生长情况下, 转基因植株较野生型植株生理指标差异不明显, 但在干旱胁迫36 h后, 转基因株系L1、L3、L6的相对含水量可达71.0%~71.9%, 而野生型拟南芥相对含水量只有65.6%; 转基因植物脯氨酸含量比野生型拟南芥分别高33.2%、15.7%和38.5%; 叶绿素含量比野生型分别高13.9%、10.3%和7.3%; 相反的, 转基因植株的丙二醛含量比野生型分别低33.7%、33.3%和33.5%。综上结果表明, 小麦WZY2-1基因可以提高拟南芥对干旱引起的氧化胁迫抵抗能力。
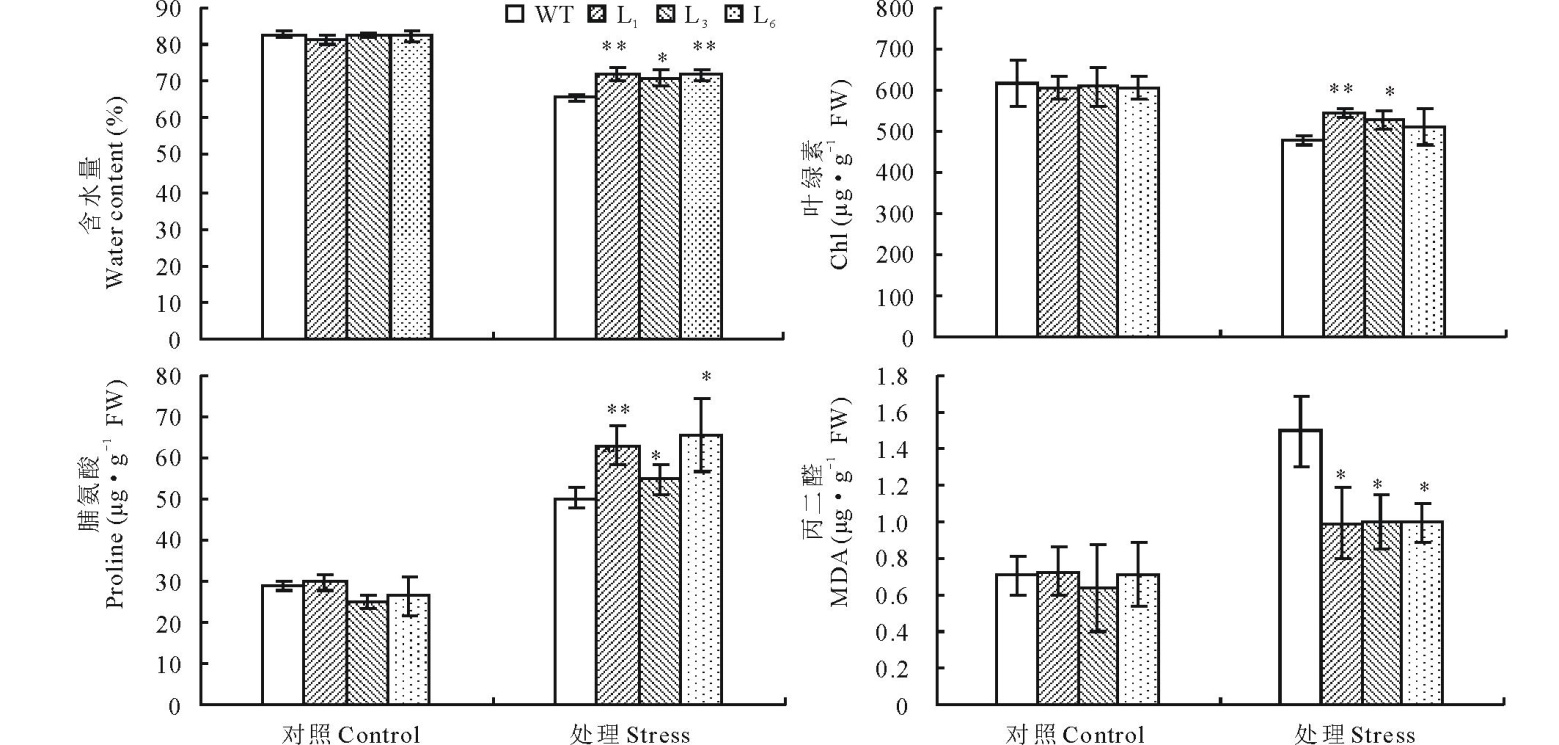 | 图5 干旱胁迫下野生型拟南芥和转基因拟南芥的生理指标 柱状图上星号表示野生型(WT)和转基因(L1、 L3 和 L6)生理指标的差异显著性(* , P< 0.05; * * , P< 0.01)。Fig.5 Analysis of physiological indexes of wild-type and transgenic A. thaliana under drought stress Different asterisk on top of bars indicate the significant differences of the physiological indexes between wild-type(WT)and transgenic A. thaliana(L1, L3 and L6)(* , P< 0.05; * * , P< 0.01). |
解析植物抗逆分子机制, 有利于获得优良抗逆性状的作物, 以提高作物的产量。干旱是影响作物生长发育和产量的主要因素之一。脱水素作为植物抗逆蛋白的重要成员, 响应干旱、低温、高温等多种逆境胁迫[19]。脱水素具有较强的亲水性, 在细胞缺水的情况下, 可以充当溶剂分子, 防止细胞内蛋白的聚集失活[5]。脱水素亚细胞定位多种多样, 但主要定位于细胞质和细胞核, 也有定位在线粒体、叶绿体和细胞膜等[10, 20, 21]。本研究将WZY2-1-GFP融合蛋白在洋葱表皮细胞瞬时表达, 观测结果发现该融合蛋白主要定位在细胞核和细胞膜。许多研究发现脱水素入核与S片段相关, 然而WZY2-1蛋白不含S片段, 这与先前研究结果一致:S片段与脱水素入核相关但不是必需的[22]。
人们研究发现脱水素可以结合金属离子和其他生物大分子, 像蛋白质、核苷酸以及带负电的磷脂分子等。认为在逆境环境下脱水素具有酶的保护剂、离子缓冲剂、膜的稳定剂及分子伴侣等功能[23]。脱水素作为酶的保护剂主要通过阻止逆境胁迫下的蛋白聚集, 但是对于其具体机制尚不清楚。LDH酶是一种冷敏感性酶, 已经被用来研究脱水素的酶保护功能参照物。本研究表明, 小麦WZY2-1蛋白具有保护温度胁迫下LDH酶活性的功能。目前普遍认为, 脱水素保护LDH酶活性的机制, 是通过在蛋白外围形成“ 分子盾” 保护蛋白的活性[20]。
植物遭受逆境胁迫会积累大量的超氧阴离子, 引起细胞膜氧化。脯氨酸作为渗透物质, 植物体内游离脯氨酸含量往往决定植物的抗旱能力[24]。对野生型和转WZY2-1基因的拟南芥植株脯氨酸含量的分析表明, 干旱胁迫下转基因拟南芥植株脯氨酸含量较高。光合作用是植物积累有机物的重要过程, 而植物进行光合作用需要大量叶绿素。在干旱条件下, 植物不仅会引起细胞膜受损, 也会影响叶绿素的合成, 导致植物光合效率降低[25]。本研究表明, 转小麦WZY2-1基因的拟南芥植株具有较高的叶绿素含量。丙二醛是膜脂过氧化的产物, 植物体内丙二醛含量是衡量植物遭受氧化胁迫程度的重要指标[14]。本实验测定结果表明, 野生型拟南芥较转基因植株具有更高的丙二醛含量。综上结果表明, 异源表达小麦WZY2-1基因可以提高拟南芥的干旱胁迫耐受性。
通过对WZY2-1蛋白的亚细胞定位发现, WZY2-1蛋白主要定位在细胞核和细胞膜。实验还分析了温度胁迫下WZY2-1蛋白对LDH酶的保护作用, 发现WZY2-1蛋白可以保护LDH酶的活性, 还获得了转WZY2-1基因的拟南芥植株, 通过表型和生理指标研究表明, 小麦WZY2-1基因可以提高拟南芥对干旱胁迫的耐受性。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|