

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
种质及变温对羊草种子萌发的影响
[杨伟光1, 2, 3*, *  , 刘盼盼
, 刘盼盼1, 2*, * , 袁光孝1, 2 , 毛培胜4 , 齐冬梅1 , 董晓兵1 , 刘辉1 , 李晓霞1, * , 刘公社1, * ]
, 刘盼盼, 袁光孝, 刘公社]
|
|


作者简介:杨伟光(1983-),男,内蒙古巴彦淖尔人,副研究员,在读博士。E-mail: anda580@163.com;刘盼盼(1990-),女,河南周口人,硕士。E-mail: a947148754@163.com.**共同第一作者These authors contributed equally to this work.
针对羊草种子萌发率低的问题,对43份不同种质的羊草种子,在两种变温条件下的萌发率进行了研究,并对恒温、变温、变温时长、变温时段对羊草种子萌发率的影响进行了测试。结果表明,羊草种子萌发率受种质影响较大,不同种质的羊草种子萌发率变幅为10%~95%,差异极显著。此外,羊草种子萌发亦受温度调控,恒温条件下(16、20、22、28、37 ℃)种子萌发率均较低,变温28 ℃(12 h)/16 ℃(12 h)则显著提高了种子萌发率,增幅为50%~90%。且变温处理对种子萌发有“时间剂量累加效应”,即变温处理时间越长,萌发率提高越大。在第1、2天时变温处理的种子萌发率与恒温对照相比,达到极显著差异,且在24 h内接收低温信号的剂量时长在2~16 h,高于16 h萌发率不随时间增加,低于2 h萌发率提高不显著。此外还发现,萌发24 h之内,羊草种子对变温处理有一个敏感的“窗口期”,即8~12 h,在这个时段内变温处理对种子萌发率提高最为显著。
, LIU Pan-pan, YUAN Guang-xiao, LIU Gong-she
To clarify the reasons for low seed germination rate, germination of 43 different germplasm lines of sheep grass was studied at constant temperatures of 16, 20, 22, 28 and 37 ℃, and under a variable temperature regime of 28 ℃ (12 h) and 16 ℃ (12 h). It was found that seed germination percentage of sheep grass differed between germplasm lines, with values ranging from 10% to 95%. Seed germination was low under constant temperature conditions, while the alternating 28 ℃/16 ℃ treatment increased the seed germination rate by 50%-90% ( P<0.01). Furthermore, the longer the daily high temperature exposure, the higher germination rate was. The seed germination rate was increased very significantly ( P<0.01) on 1 day or 2 days of variable temperature treatment, compared with the constant temperature control. However, on the first day of germination, variable temperature less than 2 hours cannot significantly promotes germination, and germination rate improves significantly in 8-12 hours treatments, which considers as the “window period”.
种子是裸子植物和被子植物特有的繁殖体, 大多数有花植物通过有性生殖和产生种子繁衍后代, 种子的活力和成功萌发、成苗决定着植物种群的繁衍和生存, 直接影响作物的产量。
羊草(Leymus chinensis)是我国重要的多年生禾本科牧草资源, 由于其野生性较强, 种子存在着发芽率偏低的现象[1], 严重影响了羊草的生产应用。目前关于羊草萌发率低的主要原因无统一的定论, 有抑制物引起的生理性休眠[2], 有因种子本身基因型或收获地的土壤及气候类型[3]、发芽温度、物理阻碍和胚乳阻碍[4, 5, 6, 7]等说法阻碍萌发。研究表明, 除热水浸种不利于羊草种子萌发外, 其余方法如浓硫酸处理、冷层积处理、PEG处理、硝酸钾溶液处理以及清水浸种等方法都能在一定程度上破除羊草种子休眠, 提高发芽率[8, 9, 10, 11, 12, 13]。另外, 普遍研究认为变温能够促进羊草种子萌发, 而恒温则不利于其萌发。但以上这些研究尚无系统性的结论, 主要原因是材料有限, 均以1份或几份羊草材料进行研究, 并未开展不同基因型之间的比较研究。虽然变温促进羊草种子萌发已证实, 但变温处理的窗口期及处理最短时长还不清楚。因此, 需要对这些问题开展系统和深入的研究, 进一步揭示羊草种子萌发的规律, 为生产实践提供理论基础。
本研究主要以43份羊草种质为材料, 对其萌发规律进行研究, 包括羊草不同基因型间种子萌发差异、羊草种子萌发的最优条件、变温对种子萌发的影响等, 旨在阐明不同基因型羊草种子萌发的规律、初步探究变温调控羊草种子的萌发规律, 为羊草种子萌发研究提供基础理论数据, 同时对羊草新品种选育提供参考资料。
本研究所有羊草种质材料均来源于中国科学院植物研究所种质资源圃, 共43份, 收获地点为北京植物园, 东经116° 12', 北纬39° 59', 海拔 61.6 m, 年均温度12.8 ℃, 年降水量 532.6 mm, ≥ 0 ℃积温4400 ℃, 无霜期 190~195 d, 极端最低温度-13.8 ℃, 极端最高温度 38 ℃, 土壤为棕壤土。收获年份在2014-2015年, 具体见表1。其中温室内测试出苗率所用材料为41份, 光照培养箱内测试萌发率的材料为35份, 用于变温测试萌发率的材料总计为4份。
| 表1 试验材料来源与编号 Table 1 Origin and codes of experimental |
1.2.1 温室内营养钵出苗率测定 取直径11 cm装有混合营养土(营养土: 蛭石=1: 1)的营养钵, 用平皿轻轻压平土表, 将营养钵放到盛水的托盘内吸水过夜。取出50粒人工挑选的种子均匀地撒在土表, 4次重复。用混合营养土均匀地撒在土表, 形成0.5~1.0 cm的覆盖层, 并用平皿轻轻按压覆盖层, 用喷雾壶喷湿土表。将托盘放到温室内(自然光照, 相对湿度35%~45%, 温度16~38 ℃), 计算20 d内出苗率, 出苗标准为胚芽鞘伸出土表1 mm。
1.2.2 光照培养箱萌发率测定 取铺有浸湿滤纸的玻璃平皿, 将30粒人工挑选的种子均匀地排列在滤纸上, 4次重复。将皿盖倾斜45° 盖到平皿上, 平皿和皿盖一起放入托盘内, 托盘内倒入1 cm深的蒸馏水。将托盘放到光照培养箱{光照强度40 lx, 相对湿度35%, 温度28 ℃[12 h昼(day, D)]/16 ℃[12 h夜(night, N)}中, 计算20 d内的萌发率, 萌发标准为胚根伸出1 mm。
1.2.3 不同温度处理种子萌发 1)变温与恒温处理:变温处理为28 ℃(12 h昼)/16 ℃(12 h夜), 恒温处理分别设16、20、22、28和37 ℃, 光照培养箱测定萌发率。
2)变温时长处理:在羊草种子萌发的前6 d, 设18个不同时长变温(change temperature, CT)处理, 从变温处理6 d逐步缩小到1 h, 见表2。变温处理后将种子取出立即放入恒温28 ℃光照培养箱内萌发, 对照组为一直恒温(steady temperature, ST)和变温萌发, 4次重复。
| 表2 变温时长处理 Table 2 The treatments of changing temperature for different times |
3)变温时段处理:在萌发的前6 d, 设14个处理(表3), 分别于不同萌发时段进行28 ℃/16 ℃变温处理, 其他时段均置于恒温28 ℃光照培养箱内萌发, 对照组为一直恒温萌发, 4次重复。
| 表3 变温时段处理 Table 3 The treatments of changing temperature at different times |
统计分析采用SPSS 17.0和Microsoft Excel软件完成, 使用Microsoft Excel和Origin 8.0软件作图。采用单因素方差分析和LSD最小显著差异法检验不同基因型和不同变温时长、时段处理的羊草种子萌发率的差异性。
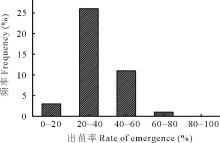
通过对41份不同基因型的种子在温室内的出苗率进行测定, 结果表明不同种质间出苗率差异较大, 出苗率从10.00%到61.33%, 出苗率集中分布在20%~30%(图1), 占所有出苗种质材料的46.34%, 表明羊草种子出苗率受种质影响较大。
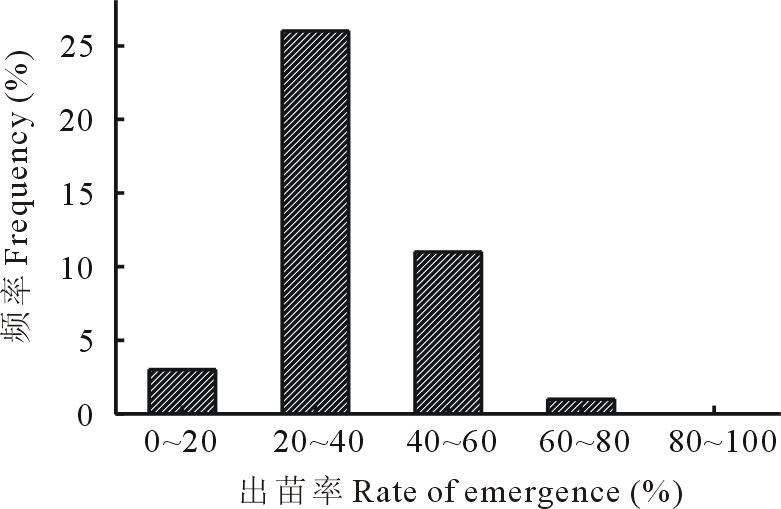 | 图1 41份羊草种质材料出苗率分布Fig.1 Distribution histogram of seed emergence rate of 41 different germplasms |
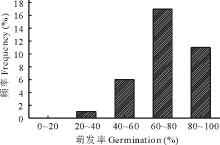
此外, 通过光照培养箱对35份羊草材料种子萌发率进行测定, 结果显示不同材料种子萌发率差异也较大, 从36.67%到95.00%, 主要集中在60%~100%, 占所有萌发种质材料的85.71%(图2), 进一步说明羊草种子的萌发率高低与种质相关。
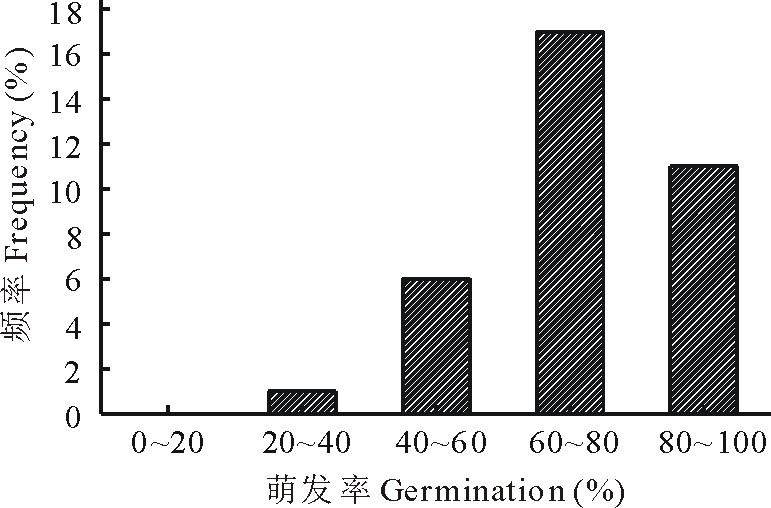 | 图2 35份羊草种质材料种子萌发率分布Fig.2 Distribution histogram of seed germination rate of 35 different germplasms |
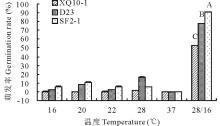
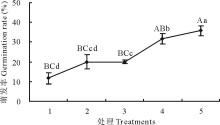
2.2.1 恒温、变温条件下羊草种子萌发规律 选取的XQ10-1、D23、SF2-1这3种羊草种质材料, 恒温处理下(16、20、22、28、37 ℃)3种材料种子萌发率均低(< 20%), 甚至萌发率为零(图3), 同一种质材料的羊草种子各恒温间的萌发率差异不显著。而变温28 ℃ (12 h)/16 ℃ (12 h), 3种羊草种子萌发率显著增加, 增幅50%~90%, 且种质间的种子萌发率也不同, XQ10-1、D23、SF2-1分别为52.50%、77.78%和90.83%, 表明变温可以显著提高羊草种子萌发率。
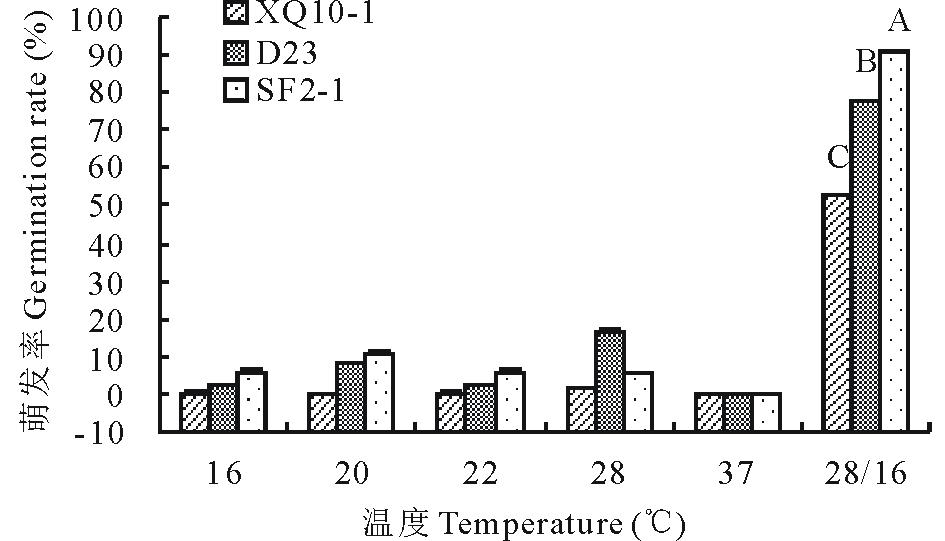 | 图3 不同温度条件下3种羊草种子的萌发率 图中的数据为平均值± 标准误, 不同大写字母或小写字母代表材料间或处理间在 P< 0.01或P< 0.05水平上差异显著, 下同。Fig.3 The seed germination rate of 3 type L. chinensis under different constant temperature and 28 ℃ (12 h)/16 ℃ (12 h) variable temperature The data in the picture are the mean± SE.The different capital or small letters indicate significant difference at the 0.01 or 0.05 level within materials or treatments.The same below. |
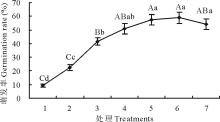
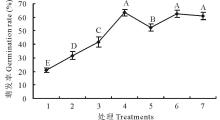
2.2.2 不同时长的变温处理下羊草种子萌发规律 选取4种羊草种质材料(D50, XQ10-1, 2015年收获的SF2-1和149), 进行6 d内不同时长变温处理后再放到28 ℃恒温条件下进行萌发, 发现4种羊草种子所有的不同时长变温处理的萌发率均极显著(P< 0.01)高于恒温对照, 且种子的萌发率在变温处理1~5 d, 随着变温时间的增加, 萌发率有显著提高(图4)。除材料D50外, 其余3种材料在变温时长≥ 4 d的羊草种子萌发率与变温对照相比差异不显著, 表明变温处理3~4 d后羊草种子萌发率达到萌发最高值。
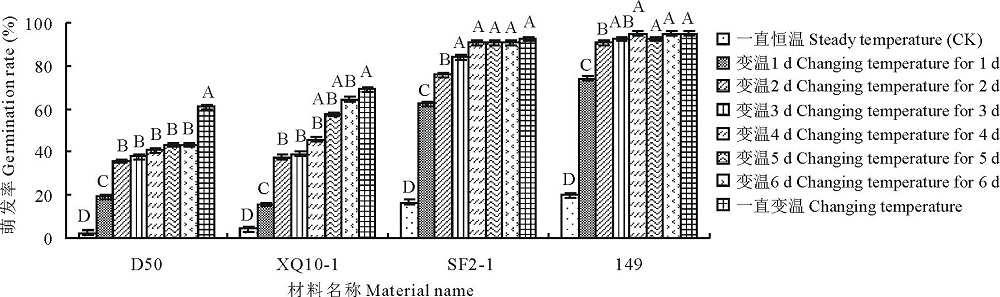 | 图4 4种羊草种质材料的种子萌发率对不同时长变温处理的响应Fig.4 The seed germination rate of 4 different L. chinensis under 28 ℃ constant temperature and 28 ℃(12 h)/16 ℃(12 h) variable temperature for various days |
此外, 与恒温萌发对照相比, 变温处理1和2 d, 种子萌发率极显著提高(P< 0.01)。变温处理2与1 d相比, 种子萌发率有显著提高; 而处理3 d以上与前一天相比, 除SF2-1之外, 其余种质材料的种子萌发率均无显著差异。因此, 与恒温对照相比, 变温处理1~2 d即可显著提高羊草种子萌发率。
上述结果表明, 变温处理1 d, 羊草种子萌发率就有极显著提高(P< 0.01)。为了进一步探究1 d内, 即24 h内变温时长对羊草种子萌发影响的窗口期, 以SF2-1为试验材料, 对萌发第1天以4 h为一个梯度进行变温处理, 结果表明6个不同梯度处理中, 羊草种子萌发率均较恒温有极显著提高(P< 0.01), 而且变温时间越长萌发率提高越大, 当变温时长大于16 h后, 萌发率不随时间变长而继续提高(图5), 其中变温8 h即可显著提高羊草种子的萌发率(P< 0.01)。
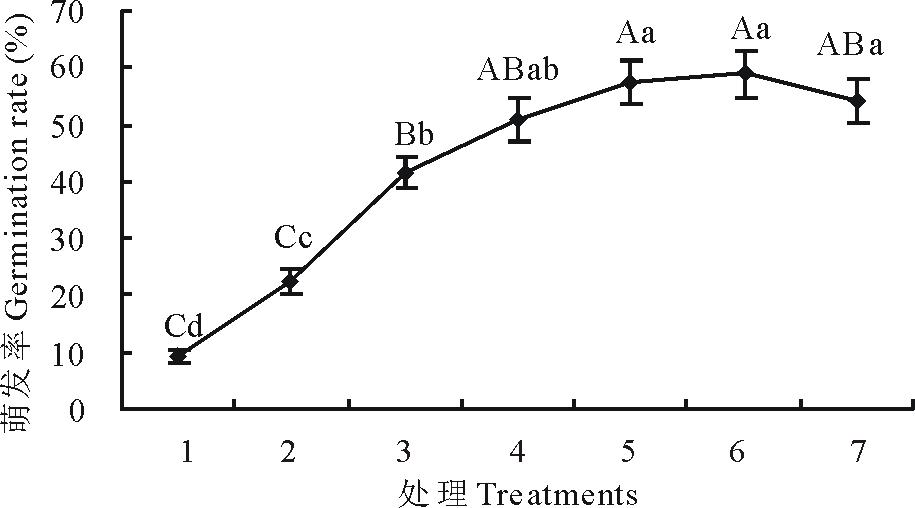 | 图5 4 h梯度不同时长变温处理对羊草种子萌发的影响Fig.5 Germination rate in different duration of variable temperature under 4 h interval |
进一步对萌发第8~12 h的1 h梯度变温处理实验表明, 当变温处理1 h, 虽然羊草种子萌发率有提高, 但与恒温对照相比差异不显著(P> 0.05)。当变温时长为2 h以上时, 羊草种子萌发率较恒温对照显著提高(P< 0.05), 变温时长为4 h时, 萌发率极显著(P< 0.05)提高, 说明变温处理2 h为羊草种子萌发率提高的窗口期, 见图6。
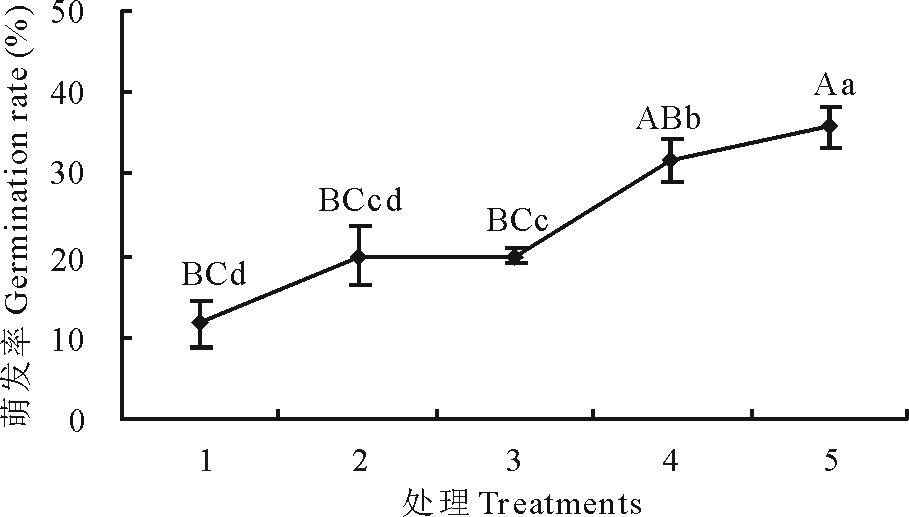 | 图6 1 h梯度不同时长变温处理对羊草种子萌发的影响 1:恒温对照Steady temperature (CK); 2:变温4 h Change temperature for 4 h; 3:变温8 h Change temperature for 8 h; 4:变温12 h Change temperature for 12 h; 5:变温16 h Change temperature for 16 h; 6:变温20 h Change temperature for 20 h; 7:变温24 h Change temperature for 24 h.Fig.6 Germination rate in different duration of variable temperature under 1 h interval |
2.2.3 不同时段变温处理对羊草种子萌发率的影响 为了探寻28 ℃/16 ℃变温处理对种子萌发作用的具体时间段, 选取4种不同基因型的种子进行了6 d内不同时段的变温处理, 即分别在第1天, 第2天, 依次到第6天进行变温处理, 结果表明, 与恒温对照相比, 第1天变温处理, 即可极显著地提高种子的萌发率(P< 0.01); 随着变温处理时间段的延后, 种子萌发率的增长越少。与变温对照相比, 羊草种子萌发期间, 越早对其进行变温处理, 种子最终萌发率提高得越多, 而较晚进行变温处理, 例如第6天进行变温处理, 萌发率提高不明显(P> 0.05), 见图7。
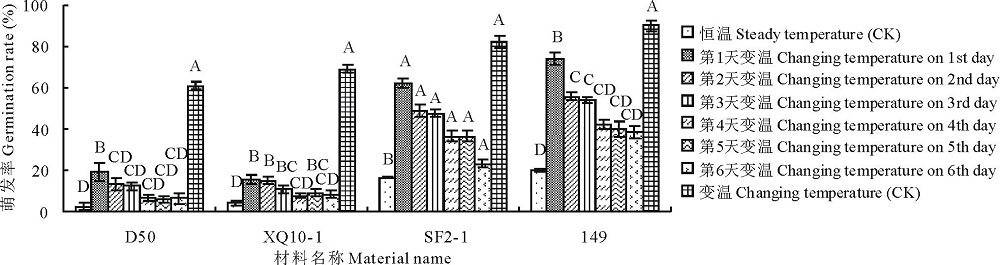 | 图7 4种基因型羊草种子的萌发率对不同时段变温处理的响应Fig.7 The germination rate of 4 genotype seeds in 28 ℃ constant temperature and 28 ℃(12 h)/16 ℃(12 h) variable temperature at different days |
在6 d内不同变温时段变温处理的研究中, 发现萌发第1 天进行变温, 萌发率极显著提高。为了进一步探究变温处理的敏感期, 选取SF2-1为试验材料, 将萌发第1天分成6个4 h的时段进行变温处理, 结果表明, 与恒温对照相比, 各时段变温处理的羊草种子萌发率均有极显著(P< 0.01)提高, 而且不同时段变温处理对萌发率提高幅度有差异, 萌发开始的第8~12 h变温处理, 萌发率提高最多, 即此时是变温处理最敏感时期, 见图8。
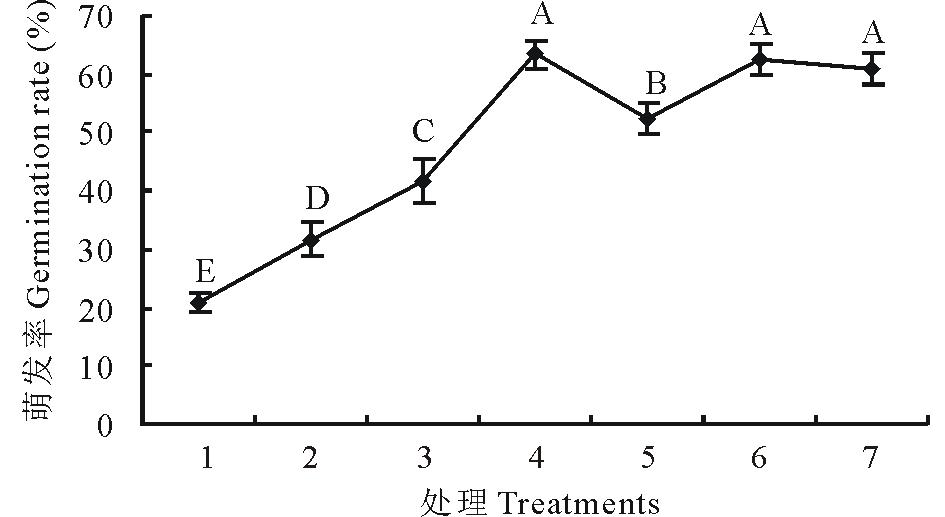 | 图8 羊草种子在萌发第1天不同时段对变温处理的响应 1:恒温对照Steady temperature (CK); 2:0~4 h变温Change temperature on 0-4 h; 3:4~8 h变温Change temperature on 4-8 h; 4:8~12 h变温Change temperature on 8-12 h; 5:12~16 h变温Change temperature on 12-16 h; 6:16~20 h变温Change temperature on 16-20 h; 7:20~24 h变温Change temperature on 20-24 h.Fig.8 The germination of seeds in 28 ℃ constant temperature and 28 ℃ (12 h)/16 ℃ (12 h) changing temperature at different time period in first day |
前期研究发现, 羊草发芽率低的主要原因是其有较强及较长的种子休眠期[2], 且羊草种子萌发率的高低不仅与种子休眠程度有关, 还与发芽温度[14]、种子活性[15]、激素种类与含量水平[8]、气候条件、土壤类型等有关[10]。且通过浓硫酸处理、冷层积处理、聚乙二醇(polyethylene glycol, PEG)处理、硝酸钾溶液处理以及清水浸种等方法均能在一定程度上提高羊草种子发芽率[6, 16]。加入氟啶酮进行羊草种子冷层积, 可完全破除羊草种子休眠, 使种子萌发率达到96%[17]; 用30%聚乙二醇处理羊草种子24 h后, 能显著提高羊草种子发芽率和幼苗活力[10]; 也有研究发现清水浸种1 d后用 30% NaOH处理60 min, 施加200 g· g-1 GA3, 可使羊草种子发芽率达到91%[8]。 以上这些研究为羊草生产实践提供了依据, 但是这些均属于外界环境因素对种子萌发的影响, 而本研究中对43份羊草种质在温室条件及光照培养箱中的萌发率进行测定, 发现不同羊草种质材料的种子萌发率差异显著, 说明羊草种子的萌发率与种质密切相关, 这一结论为今后选育具有高萌发率的羊草种质及培育羊草新品种奠定了基础, 也为下一步从分子生物学方面探究不同种质羊草种子萌发差异的分子机理奠定了理论基础。
种子萌发是一个生理生化变化的过程, 是在一系列酶的参与下进行的, 而酶的催化与温度有密切的关系。前人研究发现, 恒温不利于羊草种子发芽, 低温和变温都能显著地提高种子萌发率[2, 6, 14, 18, 19, 20]。何学青等[8]表示20 ℃(12 h)/30 ℃(12 h)可以最大限度地提高羊草种子的萌发率; 蔺吉祥等[21]在20 ℃(12 h)/30 ℃(12 h)萌发条件下, 通过对发芽率、发芽速率、开始发芽时间以及50%种子发芽天数等指标进行观察, 发现变温处理可以显著提高羊草种子的萌发率。本研究发现恒温(16、20、22、28、37 ℃)条件下, 3种基因型种子萌发率均低(< 20%), 甚至萌发率为零, 而变温28 ℃ (12 h)/16 ℃ (12 h), 3种羊草材料种子萌发率显著增加, 增幅50%~90%, 这一结论与前人研究结果相类似[18], 但是不同种质材料的种子对变温的响应是不同的, 例如本研究中选用的XQ10-1、D23、SF2-1这3种材料, 在28 ℃ (12 h)/16 ℃ (12 h)变温条件下的萌发率分别为52.50%、77.78%和90.83%, 且三者之间差异极显著(P< 0.01)。温度影响种子萌发的主要原因可能是温度影响膜透性、膜结合蛋白的活力以及种子内赤霉素GA3、细胞分裂素和 ABA的合成代谢等[22], 而在羊草中温度影响种子萌发的机理有待深入研究。
变温能够促进羊草种子萌发已被广泛研究证实, 但是变温处理的窗口期及处理最短时长目前还尚不清楚。本研究通过进一步细化1 d内的变温处理效应, 发现只要变温处理时长大于2 h时, 萌发率有显著提高, 且时长越长, 萌发率增加幅度越大, 但当时长达到16 h后, 萌发率不再随时长而提高, 说明2~16 h变温处理导致的萌发率提高有一种累加效应。此外, 不同时间段处理, 导致萌发率提高幅度不同, 前12 h内, 处理时间越往后, 萌发率提高幅度越大, 8~12 h处理, 产生了最大萌发率, 说明变温影响种子萌发的敏感期在种子萌发开始的8~12 h。在羊草种子萌发试验中, 得知吸胀过程大约在种子加水后6 h结束, 而低温是诱导种子萌发的敏感期, 在8~12 h, 这可能就是导致不同时段变温处理萌发率不同的原因。而本实验中变温时长越长, 萌发率越高, 即变温的叠加效应, 可能与低温层积的原理相似[23]。因此, 通过本研究确定羊草种子萌发响应变温的敏感期及关键时间点后, 可采用现代生物学技术方法(新一代测序技术等)阐明恒温抑制及变温诱导羊草种子萌发的分子机理, 为进一步解析羊草种子萌发的分子调控机制提供理论依据。
羊草种子萌发率不仅与羊草种质有关, 而且受萌发温度影响较大。41份不同羊草种质材料的种子间温室内出苗率差异较大, 出苗率从10.00%到61.33%, 出苗率集中分布在20%~30%, 另外其中的35份羊草种子材料在光照培养箱中萌发率差异也较大, 从36.67%到95.00%, 主要集中在60%~100%。恒温条件下种子萌发率均较低, 而变温28 ℃/16 ℃可显著提高种子萌发率, 增幅50%~90%。 且变温处理对种子萌发有时间剂量累加效应, 即变温处理时间越长, 萌发率提高越大。变温处理在第1天的效应最大, 1 d内影响种子萌发的时长在2~16 h, 高于16 h萌发率不随时间增加, 低于2 h萌发率提高不显著; 此外还发现, 萌发24 h之内, 羊草种子对变温处理有一个敏感的窗口期, 即8~12 h, 在这个时段内变温处理对种子萌发率提高最为显著。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|