

{kind=link}
{kind=link}
小黑麦与黑麦花序结构和籽粒特性比较
[刘汉成1, 2  , 田新会
, 田新会1 , 杜文华1, * ]
, 田新会]
|
|


作者简介:刘汉成(1982-),男,土族,甘肃武威人,讲师,在读博士。E-mail: lhchwl@163.com
植物的花序结构及其形态特征是影响种子生产性能的重要因素。对于禾本科植物而言,其花序大小、小穗数、小花数及小花的外稃、内稃和芒的性状特征直接影响种子形成和产量。本研究通过测定小黑麦品系C2、C35和黑麦品系C13、C33的花序结构特征,得到以下结果:小黑麦花序[(14.30±0.52) cm×(1.24±0.09) cm]明显大于黑麦[(13.20±0.35) cm×(0.82±0.02) cm],小黑麦花序长而粗,黑麦花序短而细;小黑麦花序的小穗数[(21.35±1.47)个]少于黑麦[(30.20±0.79)个];小黑麦每个小穗的小花数较多,为3~4朵,黑麦的小花数趋于稳定,每个小穗有2朵小花。从花序中部小穗的小花结构看,小黑麦下位护颖长[(1.27±0.11) cm]和宽[(0.26±0.03) cm]、上位护颖长[(1.30±0.09) cm]和宽[(0.23±0.04) cm]均显著大于黑麦下位护颖长[(1.04±0.05) cm]和宽[(0.08±0.01) cm]、上位护颖长[(0.94±0.10) cm]和宽[(0.06±0.01) cm];小黑麦中部小穗第1小花的外稃宽[(0.32±0.03) cm]、外稃高[(0.24±0.03) cm]和芒长[(8.35±0.51) cm]、第2小花的外稃宽[(0.35±0.04) cm]、外稃高[(0.25±0.05) cm]和芒长[(8.37±1.19) cm]极显著大于黑麦相应值[(0.26±0.01),(0.15±0.01),(5.50±0.19),(0.25±0.01),(0.17±0.01)和(5.18±0.23) cm];第1和第2小花的外稃长[(1.35±0.06),(1.37±0.06) cm]和内稃长[(0.84±0.04),(1.41±0.06) cm]均小于黑麦的外稃长[(1.49±0.05),(1.47±0.05) cm]和内稃长[(1.45±0.05),(1.47±0.04) cm]。小黑麦的穗粒数[(52.50±1.80)粒]显著低于黑麦[(58.50±2.50)粒] ( P<0.05),但其穗粒重[(2.08±0.04) g]、粒重[(0.04±0.00) g]和籽粒宽[(3.04±0.32) mm]均极显著高于黑麦 ( P<0.01),每个花序的籽粒不仅体积大,而且质量较重,其籽粒的生产性能高于黑麦。小黑麦籽粒呈椭圆形或长卵圆形,浅黄色,表皮皱缩,饱满度差;黑麦籽粒呈窄纺锤形,青灰色,表皮光滑,籽粒饱满。小黑麦和黑麦花序结构与籽粒性状的Pearson相关分析表明,小黑麦花序宽和花序基部小穗数与籽粒长显著正相关( P<0.05),花序长与籽粒宽极显著正相关( P<0.01),花序中部小穗的下、上位护颖宽分别与穗粒重和穗粒数极显著正相关( P<0.01);黑麦花序中部小穗第2小花的内稃长与籽粒长显著正相关( P<0.05),外稃高与穗粒数极显著负相关( P<0.01)。研究小黑麦和黑麦的花序结构和籽粒特征,对正确区分二者和了解其籽粒的生产性能具有重要意义,同时有利于小黑麦的示范推广。
Morphological characteristics of inflorescences are important factors in seed production performance. In Gramineae, the size of the inflorescence, number of spikelets and florets, and the characteristics of the lemma, palea and awn can influence the development of seeds. Inflorescence characteristics of different lines of triticale (C2 and C35) and rye corn (C13 and C33) were compared. The inflorescence of triticale [(14.30±0.52) cm×(1.24±0.09) cm] was clearly larger than the rye inflorescence [(13.20±0.35) cm×(0.82±0.02) cm)], spikelet numbers in rye corn (30.20±0.79) were higher than that of triticale (21.35±1.47), flowers number in rye was 2 flowers per spikelet with little variation, while flower numbers in triticale did vary ranging from 3-4 flowers/spikelet. Comparisons of flowers from the middle of the inflorescence indicated that the length [(1.27±0.11) cm] and width [(0.26±0.03) cm] of lower glume and the length [(1.30±0.09) cm] and width [(0.23±0.04) cm] of the upper glume in triticale were significantly larger than the length [(1.04±0.05) cm] and width [(0.08±0.01) cm] of the lower glume and the length [(0.94±0.10) cm] and width [(0.06±0.01) cm] of the upper glume in rye ( P<0.05). The width [(0.32±0.03) cm], height [(0.24±0.03) cm] of lemma and awn length [(8.35±0.51) cm] in the first flower and the width [(0.35±0.04) cm] and height [(0.25±0.05) cm] of lemma and awn length [(8.37±1.19) cm] in the second flower of triticale was significantly higher than those in rye [(0.26±0.01) cm, (0.25±0.01) cm, (0.15±0.01) cm, (0.17±0.01) cm, (5.50±0.19) cm and (5.18±0.23) cm respectively]. Lemma [(1.35±0.06) cm; (1.37±0.06) cm] and palea [(0.84±0.04) cm; (1.41±0.06) cm] length in the first and second flower in triticale were less than in rye, (1.49±0.05) cm; (1.47±0.05) cm and (1.45±0.05) cm; (1.47±0.04) cm respectively. The number of grains per panicle in triticale [(52.50±1.80) grains] was significantly lower than that of rye ( P<0.05) but the grain weight per ear [(2.08±0.04) g], grain weight [(0.04±0.00) g] and grain width [(3.04±0.32) mm] were significantly higher than rye ( P<0.01). Triticale grains were oval or elongated oval in shape, pale yellow in colour, plump with a wrinkled epidermis while rye grains were narrowed fusiform with smooth skin and a laurel-green colour. Results of Pearson correlation analysis between flower structure and grain traits of rye and triticale revealed there were positive correlations between seed length and length of the inflorescence and the number of spikelet in the base of the inflorescence ( P<0.05), and seed width was significantly positively correlated inflorescence length ( P<0.01). Grain weight per ear and the number of grains per spike were significantly correlated with the width of the lower and upper glume ( P<0.01), respectively. Seed length in rye positively correlated to glume length of the second flower in the middle spikelet ( P<0.05), while grain number per ear and the height of lemma in the second flower of the middle spikelet were negatively correlation ( P<0.01). The structural characteristics of triticale and rye inflorescences are important for correctly distinguishing the two and understanding seed production differences.
花是植物重要的繁殖器官, 开花结果是自然界普遍规律, 开什么花结什么果是由基因决定的, 不同种群之间差异较大。花序形态对于植物类群的鉴定划分具有重要价值。禾本科植物的花序分为圆锥花序、总状花序和穗状花序3种[1]。郭本兆等[2]研究认为, 雀麦族(Bromeae)和短柄草族(Brachypodieae)圆锥花序和总状花序的苞叶和花柄简缩形成小麦族(Triticeae)的穗状花序, 这是一种系统进化过程中的简化形式。花序结构的简化或变异是进化的主要趋势[3]。耿以礼[4]根据花序的不同类型, 将小麦族和虎尾草族(Chlorideae)从禾亚科中分离出来。蔡联炳等[5]根据小麦属(Triticum)简单穗状花序的不同形状将小麦属分为线穗组、柱穗组和扁穗组。小穗是组成花序的基本结构, 小穗的类型、大小和小穗是否有柄等特征在禾本科植物中有较大差异[1], Bor[6]根据小穗的性状差异区分出了须芒草族(Andropogoneae)和黍族(Paniceae)。鹅观草属(Roegneria)植物的分类中将颖片长短和芒的有无作为分组、分系的标准[7]。构成禾本科植物小穗的基本结构为小花, 小花由内外稃、浆片、雄蕊和雌蕊组成, 小花是禾本科植物分类的重要依据, 研究小花的性状特征有助于了解穗粒数、穗粒重和籽粒大小[8]。Baum等[9]研究了小麦、小黑麦和黑麦稃片上龙骨和近芒处裂片的结构, 认为这些细微结构之间的差异是对三者进行区别分类的重要特征。
结构是功能的基础, 不同的花序结构影响和决定着作物的生产性能。苗以农等[10]研究了大豆(Glycine soja)顶端总状花序和顶端穗状花序株型的生理特点, 发现顶端具穗状花序的大豆, 其株型在不同生育时期的光合速率和水分利用效率普遍高于普通大豆类型。潘洁等[11]研究发现, 小穗生长过程中所获得的营养水平及其在花序的位置均会影响籽粒质量, 中部小穗的籽粒数、籽粒重及营养成分含量都具有明显优势。合理调控光、温和施肥量, 有利于小穗在分化过程中产生更多可育花, 提高籽粒产量[12]。熊朝军[13]研究表明, 上、下位护颖的损伤对小麦籽粒产量无明显影响, 而内、外稃损伤对籽粒产量有明显影响。
小黑麦(× Triticosecale)是由小麦属(Triticum)和黑麦属(Secale)植物经属间有性杂交和杂种染色体加倍而人工结合成的新物种, 其饲草产量高、品质好, 可以青饲, 调制青干草和青贮饲料。由于其抗寒性强, 能够在东北三省种植, 也可在北京、河北、甘肃和新疆等地的秋闲田种植, 在青藏高原高寒牧区具有广阔的应用前景[14]。由于小黑麦和黑麦均为穗状花序[15], 即每个花序由多个小穗组成, 而每个小穗又由颖片和若干朵小花组成, 这些花中靠基部的2~4朵可结实(可育花), 而上部花朵不能结实(不育花)[16], 小花由雌蕊、雄蕊和内、外稃组成[8], 在生产实践中很容易被混淆, 许多地方管理部门和农牧民对小黑麦和黑麦区分不清, 认为已经在种植“ 小黑麦” (事实上是黑麦), 不愿意接受真正的小黑麦, 对小黑麦示范推广极为不利[14]。而目前国内外对黑麦和小黑麦的研究主要集中在小黑麦育种[17]、引种试验[18, 19, 20, 21, 22]、生产性能[14, 23, 24]和遗传研究[25, 26, 27, 28, 29]等方面, 尚未有对其花序结构和籽粒特性进行比较的研究报道。因此, 本研究以生长于青藏高原东北缘高寒牧区的小黑麦和黑麦品系为试材, 通过测定和比较小黑麦和黑麦的花序结构和籽粒特性, 以正确区分二者, 并了解其籽粒的生产性能, 对小黑麦的示范推广具有重要意义。
供试材料为甘肃农业大学培育的性状稳定、能够代表小黑麦和黑麦花序结构和籽粒特征的六倍体小黑麦品系C35、C2和二倍体黑麦品系C33、C13。
试验地位于兰州大学高寒草甸与湿地生态系统定位研究站。该站位于甘南藏族自治州合作市, N 34° 57'136″, E 102° 53'54″, 海拔2954 m, 年平均气温3.2 ℃, 无霜期113 d, 年降水量671.7 mm。高寒草甸土, 土壤有机质7.56 g· kg-1, 速效氮382.25 mg· kg-1, 速效磷98.84 mg· kg-1, 速效钾45.33 mg· kg-1, pH为7.55[14]。前茬为天然草场, 无灌溉条件。播种日期为2016年5月12日。
随机区组设计, 条播, 行距20 cm, 播种深度3~5 cm, 播量按照750万基本苗· hm-2计算而得, 每个小黑麦和黑麦品系重复3次, 小区面积为8 m2(2 m× 4 m)。试验地周围1 m种植保护行, 播种前和植株拔节期分别施氮肥160 kg N· hm-2。试验期间及时清除杂草, 未进行灌溉。
1.4.1 花序结构 分别于黑麦和小黑麦开花期, 从品系C2、C35、C33、C13的小区内随机取10个植株的主穗, 剪取花序, 测定以下指标。花序长:用游标卡尺测量每个花序从基部到顶端(除去芒)的长度。花序宽:用游标卡尺测量每个花序中部的宽度。小穗数:数取每个花序的小穗数。小花数:分别数取小黑麦和黑麦花序基部、中部(小穗数为奇数者取其中数, 小穗数为偶数者取靠中大数[30])和顶部小穗的小花数。护颖和每朵小花内、外稃和芒的度量特征:用游标卡尺测量中部小穗下位和上位护颖的长度和宽度, 以及每朵小花内稃、外稃和芒的长度。
1.4.2 籽粒特性 于黑麦和小黑麦完熟期, 分别从品系C2、C35、C33、C13的小区内随机取10个植株的主穗, 剪取花序, 测定花序长, 数出小穗数。单独脱粒, 数出每个花序的穗粒数, 用电子天平称取穗粒重, 根据穗粒重和穗粒数计算每个籽粒的粒重。计算公式为:粒重(g· 粒-1)=穗粒重/穗粒数。用游标卡尺测量籽粒的长、宽、高, 籽粒长指籽粒长轴长度, 籽粒宽指垂直于籽粒腹沟的横轴最大值, 籽粒高指籽粒背腹高度。
比较小黑麦和黑麦各个指标的差异时, 以小黑麦品系C35和C2相应指标的平均值作为小黑麦该项指标的特征值, 黑麦品系C33和C13相应指标的平均值作为黑麦该项指标的特征值, 用Excel 2010计算均值和标准差, 用SPSS 19.0软件的配对T测验进行方差分析。小黑麦和黑麦不同品系的方差分析也用SPSS 19.0软件的配对T测验完成, 不同指标的相关性分析用该软件的二元变量Pearson相关性法。
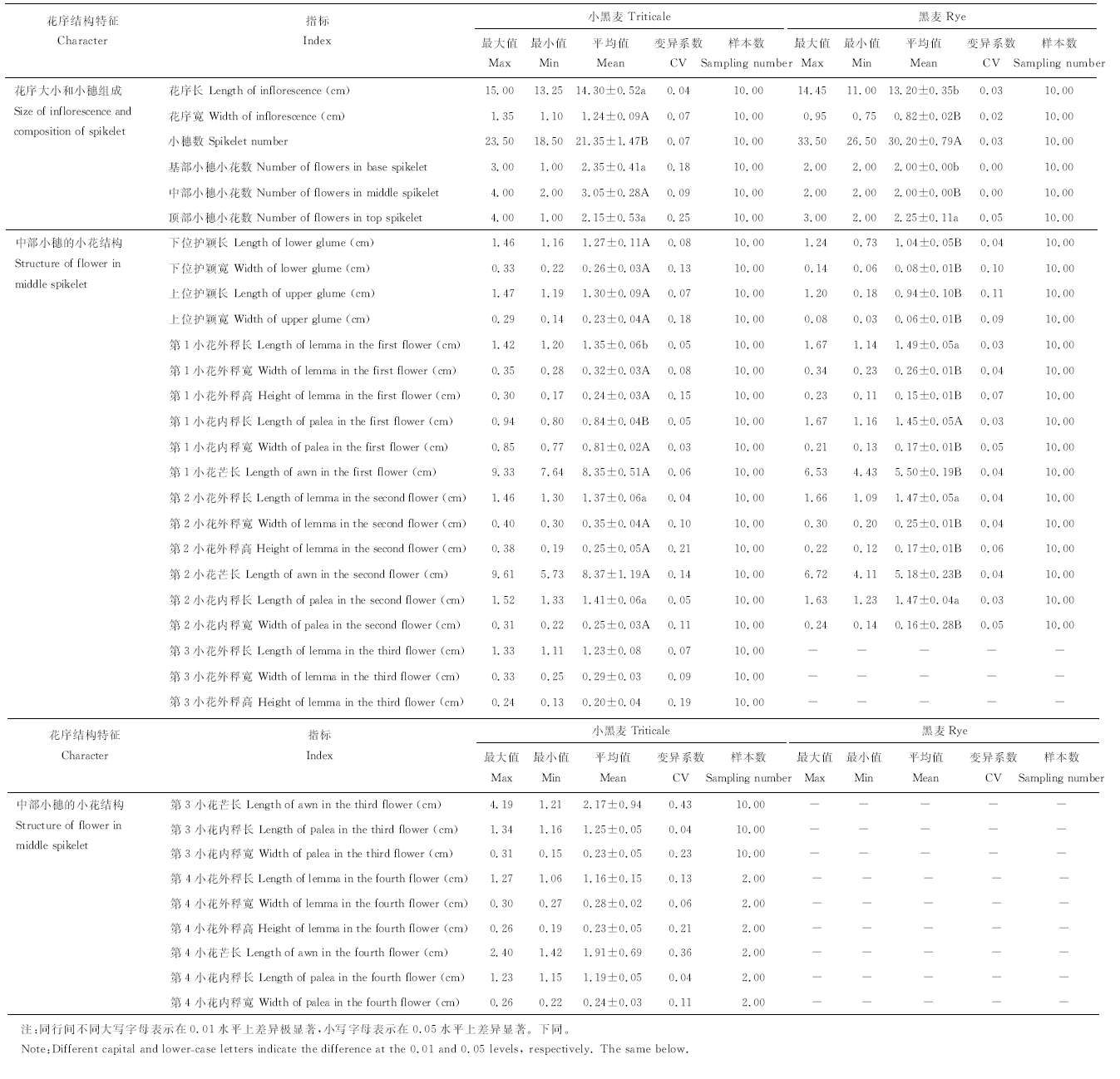
2.1.1 花序大小和小穗组成比较 禾本科植物抽穗后, 花序大小和形状是鉴别物种的主要结构特征, 小黑麦和黑麦同为穗状花序, 其花序大小取决于小穗的小花数目和小花中相关结构的大小, 如单个小穗中的小花数目多且单个小花中内稃、外稃等结构较大, 则该花序结构的长度和宽度会相应变大。通过测定小黑麦和黑麦花序大小和不同位置小穗的小花数等相关指标(表1)可以看出, 小黑麦花序长[(14.30± 0.52) cm]显著大于黑麦[(13.20± 0.35) cm](P< 0.05), 花序宽[(1.24± 0.09) cm]极显著大于黑麦[(0.82± 0.02) cm](P< 0.01), 说明小黑麦花序长而粗, 黑麦花序短而细(图1)。小黑麦花序的小穗数极显著低于黑麦(P< 0.01)。从不同部位小穗的小花数来看, 小黑麦基部小穗的小花数显著高于黑麦(P< 0.05), 中部小穗的小花数极显著高于黑麦(P< 0.01), 顶部小穗的小花数与黑麦相似, 基本都维持在2个左右, 说明小黑麦单个小穗的小花数均大于或等于黑麦。
| 表1 小黑麦和黑麦的花序结构特征比较 Table 1 Structural charactertic of triticale rey |
 | 图1 小黑麦和黑麦的花序形态Fig.1 Inflorescence morphology of triticale and rye |
2.1.2 中部小穗的小花结构特征比较 为了更详细地比较小黑麦和黑麦花序结构的差异性, 本研究测定了小黑麦和黑麦不同品系花序中部小穗中小花的详细结构特征(表1)。结果表明, 小黑麦中部小穗平均有3朵小花, 其中2个花序中部小穗有4朵小花, 而黑麦中部小穗的小花数比较稳定, 均为2个。从二者小花的护颖、内稃、外稃和芒的结构特征可以看出, 小黑麦第1小花的外稃长显著小于黑麦(P< 0.05), 第2小花的内、外稃长与黑麦无显著差异(P> 0.05), 小黑麦其他指标特征均极显著大于黑麦(P< 0.01)。
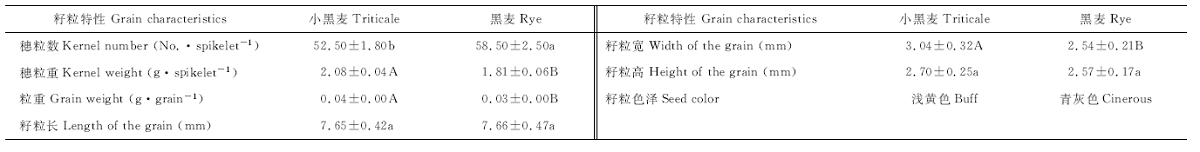
2.1.3 籽粒特性比较 花序结构是影响籽粒特性和生产性能的主要因素, 如小穗数、小花数和小花结构均可影响穗粒数、穗粒重和籽粒大小等特征。为了解花序结构对籽粒特性和生产性能的影响, 本试验在小黑麦和黑麦完熟期取C2、C35、C13、C33品系的主穗, 分别测定了穗粒重、粒重, 以及籽粒长、宽、高, 并观测了色泽(表2)。虽然小黑麦的穗粒数显著低于黑麦(P< 0.05), 但其穗粒重、粒重和籽粒宽均极显著大于黑麦(P< 0.01), 籽粒长和高与黑麦无显著差异。可见, 尽管小黑麦的穗粒数少于黑麦, 但由于其粒重极显著大于黑麦, 穗粒重也极显著大于黑麦。另外, 小黑麦籽粒比黑麦宽。从籽粒形态而言, 小黑麦籽粒呈椭圆形或长卵圆形, 浅黄色, 表皮皱缩, 饱满度差; 黑麦籽粒呈窄纺锤形, 青灰色, 表皮光滑, 籽粒饱满(图2)。
| 表2 小黑麦和黑麦籽粒特性比较 Table 2 Comparision of the grain characteristics between triticale and rey |
 | 图2 小黑麦与黑麦的籽粒形态Fig.2 Grain morphology of triticale and rye |
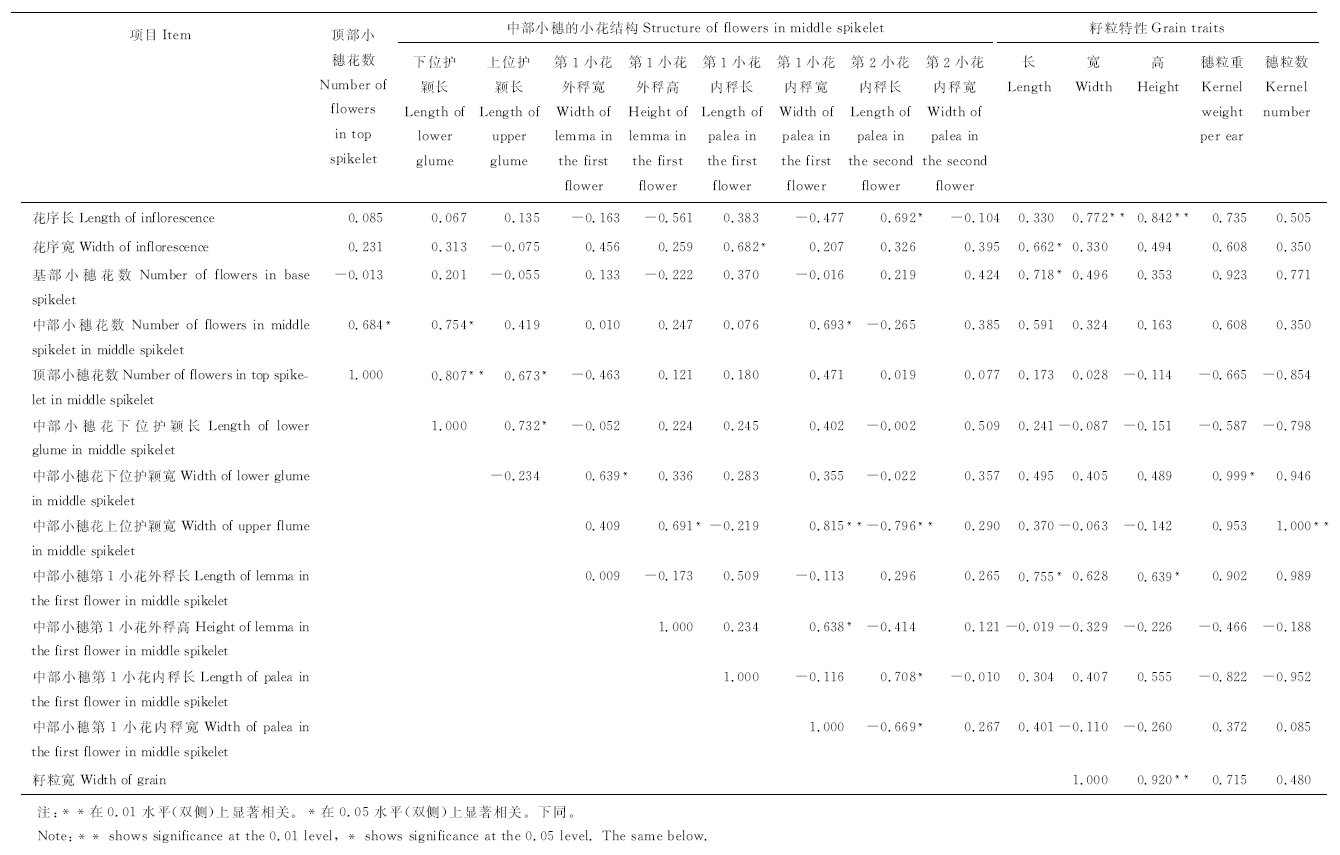
2.2.1 小黑麦花序结构和籽粒性状相关性 利用SPSS 19.0对小黑麦花序结构与籽粒性状特征进行二元变量的Pearson相关性分析(表3)可知, 小黑麦花序宽、基部小穗的小花数和中部小穗第1小花的外稃长与籽粒长显著正相关(P< 0.05); 花序长与籽粒宽极显著正相关(P< 0.01); 花序长和籽粒宽与籽粒高极显著正相关(P< 0.01), 中部小穗第1小花的外稃长与籽粒高显著正相关(P< 0.05); 花序中部小穗的下位护颖宽与穗粒重在0.05水平上显著相关, 相关系数达0.999; 中部小穗的上位护颖宽度与穗粒数极显著正相关(P< 0.01), 相关系数为1。
| 表3 小黑麦花序结构与籽粒性状的 Pearson 相关性 Table 3 Pearson correlation of the inflorescence structure of triticale and grain traits |
小黑麦花序结构间的相关性可知, 花序长与中部小穗第2小花的内稃长显著正相关(P< 0.01), 花序宽与中部小穗第1小花的内稃长显著正相关(P< 0.05), 中部小穗的小花数与顶部小穗的小花数、中部小穗的下位护颖宽和第1小花的内稃宽显著正相关(P< 0.05), 顶部小穗的小花数与中部小穗的下位护颖长极显著正相关(P< 0.01), 与上位护颖长显著正相关(P< 0.05), 中部小穗的下位护颖长和宽与上位护颖长和第1小花的外稃宽显著正相关(P< 0.05), 中部小穗的上位护颖宽与第1小花的内稃宽极显著正相关(P< 0.01), 而与第2小花的内稃宽极显著负相关(P< 0.01), 中部小穗第1小花的外稃高与该小花的内稃长、第1小花的内稃长与第2小花的内稃长、第1小花内稃宽与第2小花内稃宽均显著正相关(P< 0.05)。
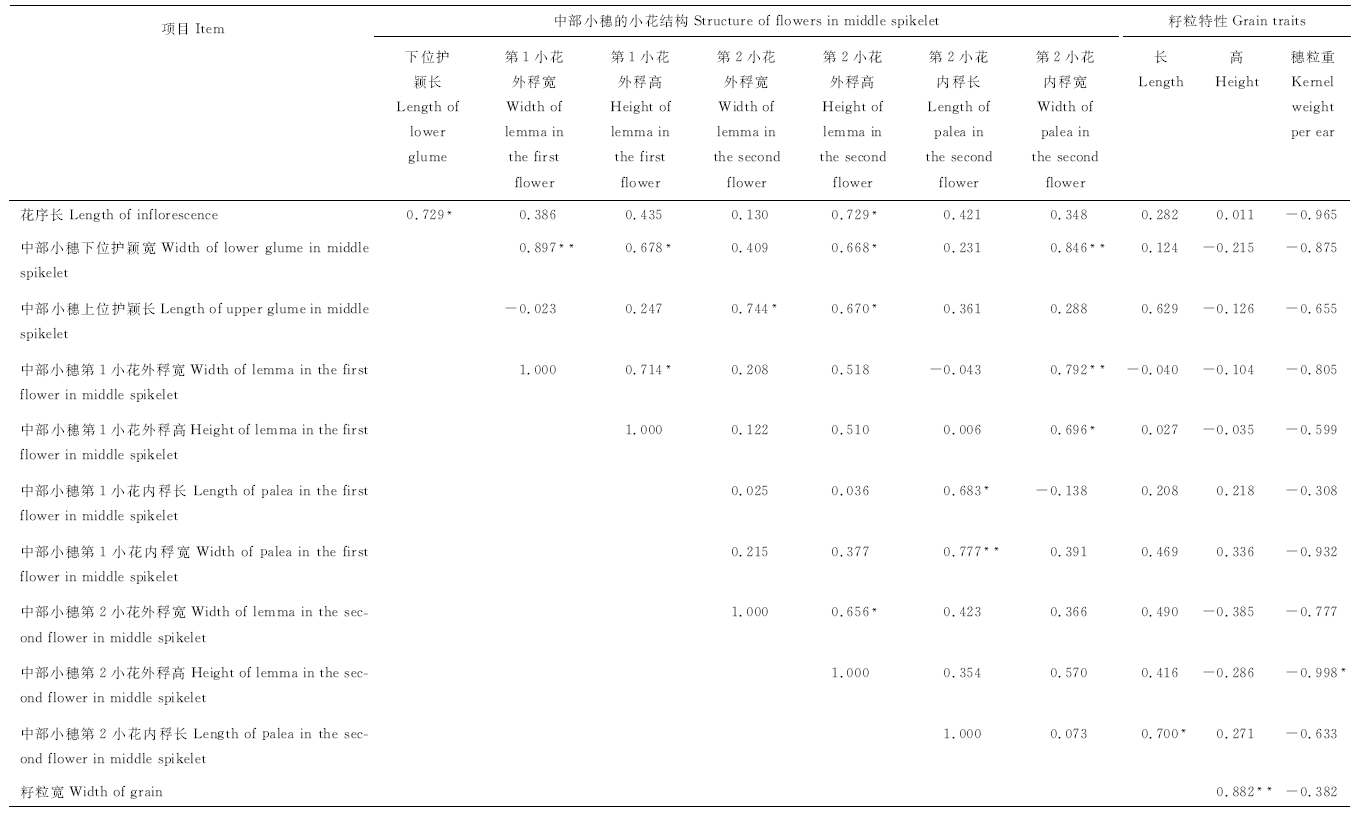
2.2.2 黑麦花序结构和籽粒性状的相关性 黑麦花序结构和籽粒性状相关性结果(表4)表明, 黑麦花序中部小穗第2小花的内稃长与籽粒长显著正相关(P< 0.05), 外稃高与穗粒重显著负相关(P< 0.05), 籽粒高和宽极显著正相关(P< 0.01)。
| 表4 黑麦花序结构与籽粒性状的 Pearson 相关性 Table 3 Pearson correlation of the inflorescence structure of rye and grain traits |
黑麦花序结构间相关性结果为:中部小穗的下位护颖长和第2小花外稃高与花序长显著正相关(P< 0.05); 中部小穗下位护颖宽与第1小花外稃宽和第2小花内稃宽极显著正相关(P< 0.01), 与第1、第2小花外稃高显著正相关(P< 0.05); 中部小穗上位护颖长与第2小花外稃宽和高显著正相关(P< 0.05); 中部小穗第1小花外稃宽与第2 小花内稃宽极显著正相关(P< 0.01), 与第1小花的外稃高显著正相关(P< 0.05); 第1小花外稃高与第2小花内稃宽、第1小花内稃长与第2小花内稃长、第2小花外稃宽与高均显著正相关(P< 0.05); 第1小花内稃宽和第2小花内稃长极显著正相关(P< 0.05)。
黑麦自1979年引入中国以来, 已成为利用价值较高的优质牧草, 适应大部分平原地区种植[31]。由于其抗寒性和抗旱性较强, 在高寒地区也有一定种植面积[32, 33, 34, 35], 并开始大面积推广。 小黑麦是小麦和黑麦经人工杂交后选育的新物种, 既具有黑麦的抗逆性, 又具有小麦高产优质等优点, 同时, 具有较强的杂交优势, 抗逆性、抗病虫害能力更强, 鲜干草产量和营养价值, 以及种子的生产性能等均高于黑麦, 逐渐成为粮饲兼用型新型作物[36]。但由于地方管理部门和农牧民对小黑麦和黑麦区分不清, 不愿意接受真正的小黑麦, 对小黑麦的示范推广造成了极大影响。
在长期进化过程中, 小黑麦种质资源丰富, 黑麦种质资源单一, 因此, 小黑麦农艺和品质性状均表现出丰富的遗传多样性, 而黑麦的农艺性状较为相近[37]。从花序形态和结构特征来看, 小黑麦花序长而粗, 黑麦花序短而细; 由于小黑麦花序上的小穗排列较稀疏, 所以小穗数较少, 相反, 黑麦花序的小穗数较致密, 因此小穗数较多(表1)。从花序不同部位小穗的小花数来看, 小黑麦基部和中部小穗的小花数显著或极显著大于黑麦, 顶部小穗的小花数与黑麦相似, 因此小黑麦单个小穗的平均小花数大于黑麦, 这也是其花序较粗的原因所在。从中部小穗的小花结构看, 第1朵小花中, 除内稃长外, 小黑麦的外稃长、宽、高和内稃宽及芒长均大于黑麦; 小黑麦第2朵小花的外稃宽和高、内稃宽和芒长均大于黑麦, 但其外稃和内稃长均小于黑麦; 小黑麦有第3或第4朵小花, 而黑麦没有。从而说明, 小黑麦中部小穗小花结构的大多数指标均大于黑麦, 从一定程度上也说明了小黑麦花序长而粗的原因。Gupta[38]建议将小黑麦定义为一个属, 但Stace[39]认为之所以不能将小黑麦从小麦属和黑麦属中分离出来单列成属, 是因为尚未找到将小黑麦与小麦和黑麦区别开来的显著特征。Baum 等[9]在分析了小麦、黑麦和小黑麦的颖片和稃片的结构特征后, 强烈认为应该把小黑麦单列为一个属。本试验小黑麦和黑麦的花序结构特征较有力地支持了Baum 等[9]的观点:小黑麦和黑麦花序结构差异明显, 应当单列为小黑麦属。
陈培元等[40]研究表明, 小麦籽粒产量与花序长显著正相关。李冬梅等[41]、陈丽霞等[42]和郭建文等[43]研究表明, 小黑麦和黑麦的种子产量与穗粒重极显著正相关, 与小穗数和穗粒数显著正相关, 与穗长无显著相关性。本试验结果表明, 虽然小黑麦的穗粒数极显著或显著低于黑麦, 但其穗粒重、粒重和籽粒宽均极显著高于黑麦, 说明小黑麦每个花序的籽粒不仅体积大, 而且较重, 因此小黑麦籽粒的生产性能高于黑麦。另外, 由于小黑麦的分蘖性能较黑麦强, 单位面积的枝条数和有效分蘖较多[14], 这更有利于其籽粒生产性能的提高。
从小黑麦和黑麦花序中部小穗的小花结构可知, 小黑麦花序中部小穗的护颖长和宽、第1和第2小花的外稃高和宽、内稃宽和芒长均显著大于黑麦(表1)。而且, 小黑麦籽粒宽和高均与护颖宽和外稃长显著正相关(表3), 从而说明, 较宽的颖片和较长的稃片有利于孕育体积较大的籽粒(表2), 这与袁光孝等[44]的研究结果一致。此外, 李寒冰等[45]认为, 禾本科植物的芒和护颖有助于叶绿体发育, 芒和护颖的基粒数和叶片相近。芒的光合量占花序的20.2%~77.1%, 光合强度远大于护颖和穗轴, 也超过旗叶[38]。而且芒在作物衰老后期, 仍能保持一定的光合活性[46]。小黑麦作为小麦和黑麦的杂交种, 其芒长大于黑麦[47]。本研究通过比较小黑麦和黑麦花序结构特征发现, 小黑麦中部小穗的护颖和第1、2小花的芒长均显著大于黑麦, 故其光合性能高于黑麦, 植株和籽粒均能获得更多能量, 有助于生产更多种子。
本研究以2个小黑麦品系和2个黑麦品系作为试材, 以研究小黑麦和黑麦的花序结构特征。结果表明, 除花序长和宽、小穗数、花序基部的小穗数、上位护颖宽、第1和第2小花的内稃长等指标有显著差异外, 2个小黑麦品系的其他相关指标均无显著差异; 2个黑麦品系中, 除花序长、小穗数、中部小穗的下位护颖长和宽、第1和第2小花的外稃高有显著差异外, 其他相关指标也无显著差异。从而说明, 本研究选取的试验材料具有代表性, 能够用以研究小黑麦和黑麦的花序结构特征。
小黑麦和黑麦花序结构差异较大, 外观形态特征明显, 应单列为小黑麦属。具体而言, 小黑麦花序长而粗, 黑麦花序短而细。小黑麦花序上的小穗排列较稀疏, 小穗数较少; 黑麦花序的小穗数排列致密, 小穗数较多。小黑麦基部和中部小穗的小花数为3~4朵, 顶部为2朵; 黑麦所有小穗只有2朵花。
小黑麦的穗粒数显著低于黑麦, 但其穗粒重、粒重和籽粒宽均极显著高于黑麦, 每个花序的籽粒不仅体积大, 而且质量较重, 其籽粒的生产性能高于黑麦。小黑麦和黑麦的籽粒特性和花序结构有一定相关性。
小黑麦籽粒呈椭圆形或长卵圆形, 浅黄色, 表皮皱缩, 饱满度差; 黑麦籽粒呈窄纺锤形, 青灰色, 表皮光滑, 籽粒饱满。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|