

{kind=link}
{kind=link}
{kind=link}
{kind=link}
基于SSR标记的小麦族St、H、Y基因组六倍体物种遗传变异及种间亲缘关系研究
[陈仕勇1, 2  , 马啸
, 马啸2 , 张新全2, * , 陈智华1 , 周青平3 ]
, 马啸, 陈智华|
|


作者简介:陈仕勇(1984-),男,四川三台人,讲师,博士。E-mail: chengshi8827@163.com
小麦族含有St、H、Y基因组的物种是禾本科小麦族最大的一个类群,它们不仅包含许多优良的牧草资源,还含有许多优异的抗逆基因资源,是牧草和麦类作物育种重要的基因库,具有重要的经济和生态价值。然而该类群物种的种属界限及系统关系一直存在争议,特别是六倍体物种间的亲缘关系争议较大,相关的研究还较少,其限制了该类群物种种质资源的进一步开发利用。研究小麦族St、H、Y基因组六倍体物种种间亲缘关系可为该类群物种的遗传多样性、种属关系及系统演化关系等提供重要参考。系统收集具有StHY、StStY、StStH、StHH基因组组成的六倍体物种25个,选用具有高通用性的SSR分子标记对其种间遗传变异、分化及亲缘关系进行分析。研究结果表明:1)27对SSR引物在46份供试材料中扩增出条带共229条,其中多态性条带213条,比率为93.01%;2)Dice遗传相似系数表明供试材料间的系数变异为0.479~0.981,平均值为0.670;StHY、StStY、StStH和StHH不同基因组组成物种内的相似系数变化分别为0.544~0.981、0.509~0.899、0.530~0.843和0.550~0.827,平均值分别为0.722、0.700、0.663、0.677;其揭示了供试物种之间存在丰富的遗传变异和不同程度的种间分化;3)聚类分析结果表明,不同基因组组成的物种间亲缘关系相对较远,相同基因组组成物种基本上能聚在一起且表现出不同程度的分化类型,其中StHY物种间分化最为显著,并与其形态特征表现出一定的一致性; 4)进一步明确了国产的披碱草属六倍体物种均为StHY基因组组成。研究结果揭示了供试的不同基因组组成的六倍体物种之间具有丰富的遗传多样性和明显的种间分化,其中StHY物种可明显分化成3种类型;相同或相似基因组组成的物种间表现出了较近的亲缘关系,研究结果支持基于基因组组成的分类体系对该类群物种分类地位的划分。
Species with St, H and Y genomes comprise the largest group in the tribe Triticeae, which includes many important forage grasses and is the source of elite genes for improving forage and cereal crops. While this group has considerable economic and ecological value, the nature of its inter genera and interspecific relationships remains a matter a dispute, especially for hexaploid species. Relatively little research has been undertaken on interspecific relationships among these species, which has limited the ability to utilize their germplasm resources. Therefore, this study aimed to reveal the interspecific affinity of the group’s hexaploid species and so to provide important information on their genetic diversity and biosystematics. A total of 25 hexaploid species with StHY, StStY, StStH and StHH genomic constitutions were systematically collected and highly transferable SSR markers were used to study their genetic variation, differentiation and affinity. A total of 229 bands were amplified based on 27 SSR markers, of which 213 bands were polymorphic (93.01%). Based on SSR data, the Dice genetic similarities coefficient ranged from 0.479 to 0.981, with an average of 0.670. The GS coefficients among StHY species ranged from 0.544-0.981 (average 0.722), while StStY species ranged from 0.509-0.899 (average 0.700), StStH species from 0.530-0.843 (average 0.663) and StHH species from 0.550-0.827 (average 0.677). The results revealed abundant genetic variation and differentiation among the species studied. Moreover, clustering based on the unweighted pair group with arithmetic mean method showed that species with the same or similar genomic constitution could be grouped together. However, some species with the same genomic constitution were highly differentiated. Those with the StHY genome divided into three distinct types, with these types also corresponding with species’ morphology. The results indicate that the genomic constitution of hexaploid species of Elymus in China are of the StHY type. Overall, the results showed that there is rich genetic diversity and differentiation among the hexaploid species and that those with the same or similar genomes have close phylogenetic relationships. These results provide a systematic basis for the classification of the hexaploid species of Elymus and Roegneria based on their genomic constitution.
小麦族(Triticeae)是禾本科植物中十分重要的一个类群, 它不仅包含小麦(Triticum aestivum)、大麦(Hordeum vulgare)等重要粮食作物, 还包含冰草属(Agropyron)、偃麦草属(Elytrigia)和披碱草属(Elymus)等许多优良牧草资源。小麦族物种的分类学与系统学研究是现代麦类作物与牧草育种中利用异种、属种质资源的必要理论基础之一[1, 2]。其中小麦族含有St、H、Y基因组的物种是其中最大的一个类群。该类群主要包括披碱草属和鹅观草属(Roegneria)等多年生属, 它们不仅是重要的牧草资源, 如老芒麦(Elymus sibiricus)和垂穗披碱草(Elymus nutans)等, 还拥有丰富的抗病等抗逆性基因资源, 是牧草和麦类作物遗传改良的重要基因库[3, 4, 5, 6]。该类群由于物种较多, 分类较为复杂, 种属界限等系统关系存在较多争议, 如披碱草属、鹅观草属这两个属的种属关系一直是小麦族种质资源研究利用中的难点, 所以开展小麦族St、H、Y基因组物种的种属间系统关系研究对该类群种质资源的保护利用等具有重要意义。
小麦族St、H基因组分别来源于拟鹅观草属(Pseudoroegneria)和大麦属(Hordeum), 但Y基因组来源未知。该类群物种大多为异源多倍体物种, 主要包括四倍体(2n=4x=28)StH、StY组成物种和六倍体(2n=6x=42)StHY、StStY、StStH、StHH组成物种。目前的研究主要集中在StH和StY四倍体物种上, 相关研究揭示了StH和StY物种的遗传变异和分化、种间亲缘关系[7, 8, 9], 并探讨了其分子系统发育关系、基因组的进化等[10, 11, 12]。然而, 关于StHY、StStY、StStH和StHH组成的六倍体物种的系统发育与进化方面的系统研究还较少, 其种属界限仍存在争议, 种间亲缘关系等有待进一步研究。
该类群六倍体物种主要分属于披碱草属和鹅观草属, 全球约30种, 主要分布于中亚和东亚地区; 其中我国分布约20种, 主要分布于我国青藏高原及内蒙古等地区。经典分类学对该类群的分类处理为每个穗轴节上含有两个或多个小穗的属于披碱草属, 每个穗轴节只含一个小穗的则属于鹅观草属[13]。然而按照细胞学基因组组成划分的处理, 含有St和H基因组的物种如StHH和StStH属于披碱草属, 含有St和Y基因组的物种如StStY属于鹅观草属, 而StHY基因组物种则属于曲穗草属(Campeiostachys)[1, 14]。后者最大的变动是将具有StHY基因组的物种分别从披碱草属和鹅观草属中划分出来归入曲穗草属。但基于基因组组成的划分结果是否能够反映该类群物种的亲缘关系, 其与经典分类学中形态学的对应关系及分类界限等问题都有待进一步研究。目前, 小麦族St、H、Y六倍体物种的研究多集中在StHY物种之间以及某个重要物种如垂穗披碱草等种质资源的遗传多样性评价[15, 16]。如基于SSR等分子标记[17, 18]、胞质基因序列[19]及细胞遗传学[20]等研究结果主要揭示了披碱草属StHY物种之间遗传变异和分化, 而系统地对小麦族St、H、Y六倍体的不同基因组物种种间关系的研究还较少。本研究选用SSR对该类群六倍体不同基因组组成物种的遗传亲缘关系以及形态学特征等进行系统分析, 拟在广泛系统收集的小麦族St、H、Y基因组六倍体物种种质的基础上, 采用SSR分子标记对其遗传变异及种间亲缘关系进行分析, 以期为该类群物种的遗传多样性、种属关系及系统演化等研究提供重要的参考, 特别是对小麦族披碱草属、鹅观草属、曲穗草属等属、种间分类界限及系统关系的研究提供参考。
本研究广泛系统地收集了小麦族St、H、Y基因组六倍体的不同基因组组成的物种25个, 共计46份种质资源。其中包括StHY基因型物种14个, StHH基因型物种2个, StStH基因型物种3个, StStY基因型物种2个, 未知基因组六倍体物种4个, 不同物种的地理来源等信息详见表1。其中PI、W6编号材料由美国国家种质资源库提供, G编号材料由中国农业部牧草种质资源库提供, J编号材料(E. humidus)由日本冈山大学植物遗传资源中心提供, H编号材料由瑞典农业大学Salomon博士提供, Y编号材料由四川农业大学小麦研究所提供, 其他材料为本课题采集材料。大部分供试材料种植于四川阿坝州红原的西南民族大学青藏基地牧草种质资源圃。
| 表1 材料列表 Table 1 The materials used in the study |
每份材料选取10株左右单株并取幼嫩叶片混合后, 采用植物DNA快速提取试剂盒DP305(北京天根)提取DNA, 采用NanoVue超微量分光光度计检测其浓度及纯度。
本研究SSR分析的引物筛选自小麦族近缘种属中高通用性SSR和EST-SSR引物共27对, 包括20对EST-SSR, 分别为披碱草属10对(Elw)、拟鹅观草属4对(B_)、赖草属(Leymus)5对(Ltc)、小麦1对(Xcwem); 7对SSR, 分别为披碱草属1对(EAGA)、大麦4对(HVM)、小麦2对(WMS), 具体引物信息及PCR流程参考陈仕勇等[21]的方法进行。PCR选扩产物采用6%变性聚丙烯酰胺凝胶电泳进行分离。
对试验结果中各标记目标扩增区域的条带进行统计分析, 将PCR扩增产物每个条带视为1个位点, 按条带有或无分别赋值, 有带记为1, 无带记为0, 构建相关二元数据矩阵。分别统计每对引物总扩增条带数(total number of bands, TNB), 多态性扩增条带数(number of polymorphic bands, NPB), 多态性条带百分比(percentage of polymorphic bands, PPB), 多态性信息含量指数(polymorphic information content, PIC)[22]。采用NTSYS-pc 2.11x 软件分析材料间的Dice遗传相似系数(genetic similarity coefficients, GS)[23], 同时基于Dice遗传相似系数进行非加权成对算术平均法(unweighted pair group method with arithmetic average, UPGMA)聚类分析和主成分分析(principal component analysis, PCA)[24], 聚类树形图节点分支的可靠性采用FreeTree软件进行Bootstrap分析, 设定重复抽样次数为1000次[25]。供试材料的遗传结构采用Structure 2.3进行分析[26]。
本研究中选用高通用性的27对SSR引物在46份供试材料中共扩增出条带(TNB)229条, 其中多态性条带(NPB)213条, 多态性条带比率(PPB)为93.01%。每对引物扩增条带变化数为3~15条, 多态性条带为1~15条, 平均每对引物扩增条带8.48条, 多态性条带数为7.93条; PIC值变化为0.126(HVM40)~0.462(Ltc1305), 平均值为0.305, 其中引物Elw4419s261、EAGA22a和Elw3264s184等在本研究表现出了较高的扩增效率(表2, 图1)。本研究中的高通用性SSR标记在供试物种的遗传关系分析中表现出了较高的多态性和扩增效率。
| 表2 SSR扩增情况 Table 2 Performance of SSR markers in the study |
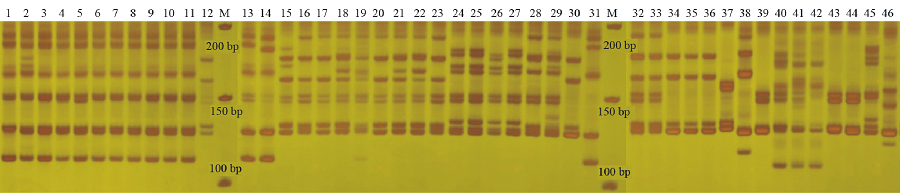 | 图1 SSR引物Elw2202s122的扩增电泳图谱Fig.1 The electrophoretogram amplificaed by SSR marker Elw2202s122 |
基于SSR标记数据, 供试材料间的遗传相似系数(GS)变异范围为0.505~0.981, 平均值为0.675, 其中最大值为E. tangtorum (G05275)和E. cylindricus (HY-4)之间的0.981, 最小值为E. schrenkianus (PI639756)和E. repens (PI565006)之间的0.505。其中StHY基因组物种间的GS值的变异范围为0.544~0.981, 平均值为0.722; StStY基因组材料间的GS值的变异范围为0.509~0.899, 平均值为0.700; StStH基因组材料间GS值的变异范围为0.530~0.843, 平均值为0.663; StHH基因组材料间GS值的变异范围为0.550~0.827, 平均值为0.677。SSR标记结果揭示了供试的六倍体物种之间存在丰富的遗传变异和不同程度的种间分化。
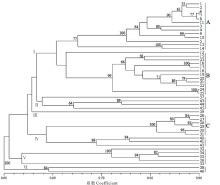
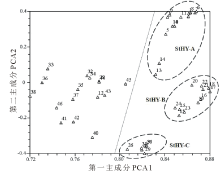
基于SSR数据的UPGMA聚类结果将供试材料分为六大类(图2)。第Ⅰ 类包括StHY基因组的物种, 其进一步被分成StHY-A和StHY-B两种不同类型, 分别为E. dahuricus和E. nutans类型。其中StHY-A类包括E. dahuricus复合群体的物种, 包括E. dahuricus、E. excelsus、E. cylindricus、E. dahuricus var. violeus、E. villifer、E. tangutorum、E. purpuraristatus和E. drobovii等, 表现出直立的穗部形态特征, 且每穗轴节上有2到多个小穗。StHY-B类包括E. nutans、E. submuticus、E. breviaristatus、E. atratus和R. aristiglumis等, 表现出下垂的穗部形态特征, 且除R. aristiglumis外其余材料的每穗轴节上仅有1枚小穗。第Ⅱ 类主要包括具有StHH基因组的3份材料, 如E. patagonicus、E. scabriglumis等和1份StStY基因组物种, 但是后者具有较低的分支支持率。第Ⅲ 类仅包括E. transhyrcanus物种1份材料, 表现一定的特异性。第Ⅳ 类包括了具有StHY和StStH基因组的物种, 如E. repens和StHY基因组的StHY-C类型物种, 包括R. kamoji类型的鹅观草属六倍体物种, 如R. kamoji、E. tsukushiensis和E. humidus等, 其穗状花序下垂且每穗轴节上为单小穗。第Ⅴ 类主要包括StHY和StStY基因组类型物种的5份材料, 如E. schrenkianus、E. tschimganicus等。第Ⅵ 类则包括2份StHH、StStH基因组类型的材料。主成分分析与聚类分析结果基本保持一致, 主成分分析二维散点图(图3)更直观地反映了供试材料间的亲缘关系, 其中前3个主成分分别能解释总变异的68.46%, 5.95%和3.39%。
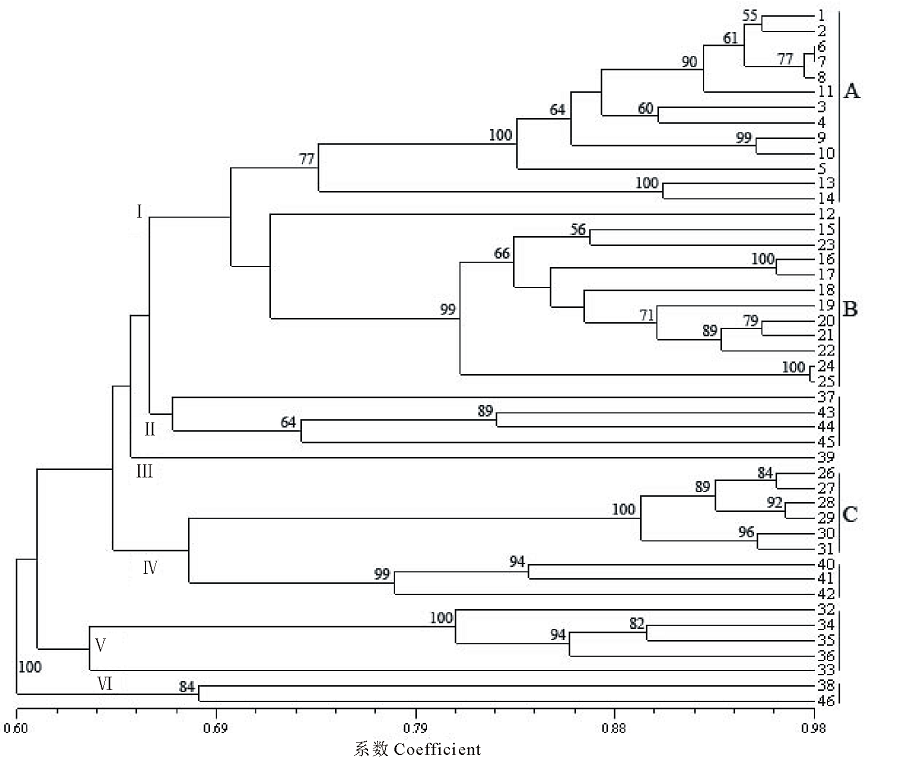 | 图2 基于SSR数据的六倍体物种UPGMA聚类Fig.2 Dendrogram of hexaploid species based on SSR data using UPGMA method |
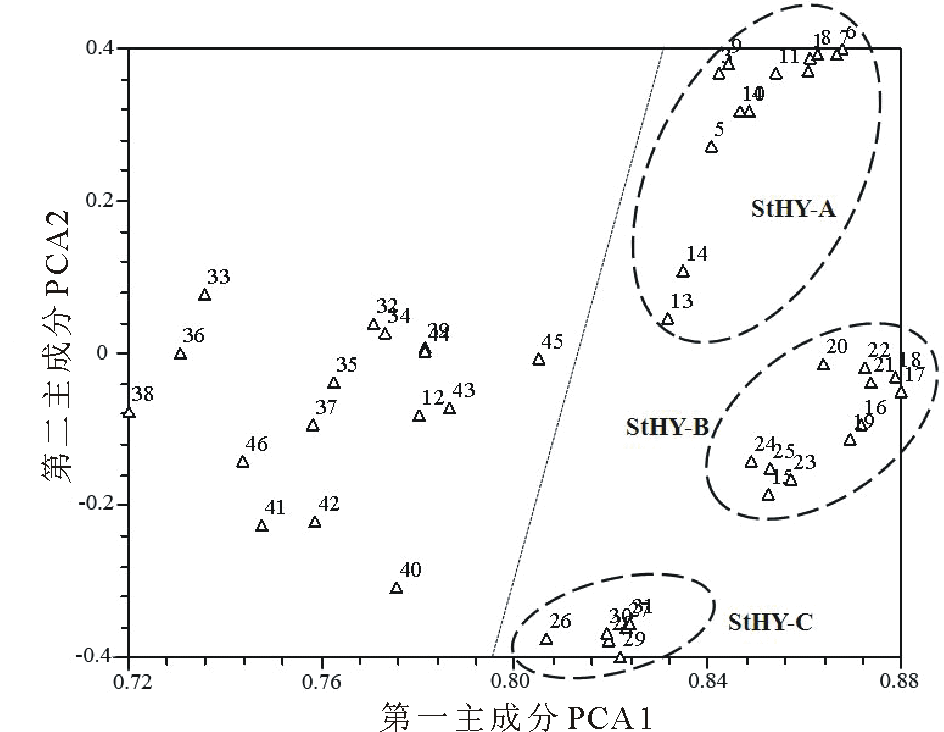 | 图3 基于SSR数据的六倍体物种的主成分分析图Fig.3 The PCA of hexaploid species based on SSR data |
根据遗传结构分析结果表明, 供试材料被划分为4个类群(K=4, 图4)。其中第Ⅰ 类(黄色)包括了所有的StHY-A类的物种。第Ⅱ 类(紫色)包括了所有的StHY-B类物种。第Ⅲ 类(红色)则包括了StHY-C类物种。第Ⅳ 类(蓝色)包括的类型较多, 包括StStY、StStH、StHH以及具有StHY基因组组成的E. schrenkianus和E. himalayanus。遗传结构分析与聚类及主成分分析结构也基本保持一致, 遗传结构分析进一步揭示了供试物种间的遗传亲缘关系及种间分化。
本研究SSR分析结果均表明E. breviaristatus、E. submuticus、E. atratus 3个物种与E. nutans聚为一类且具有较高的分支支持率, 亲缘关系非常近, 这与其形态特征相吻合, 均为下垂穗状花序且每个穗轴节含有多个小穗。从基因组组成上分析, E. breviaristatus、E. submuticus、E. atratus与E. nutans应该具有相同的StHY的基因组组成, 并形成StHY-B分化类型。这与Dong等[27]基于nrITS、matK、trnH-psbA等序列分析的结果保持一致。另一直穗物种E. dahuricus var. violeus在聚类、主成分及遗传结构分析中均聚在E. dahuricus复合群体中, 与E. dahuricus、E. excelsus、E. cylindricus等物种表现出较近的亲缘关系, 其基因组组成也应为StHY, 属于StHY-A直穗分化类型, 这与E. dahuricus复合群体材料基于荧光原位杂交FISH(fluorescence in situ hybridization)和基因组原位杂交GISH(genomic in situ hybridization)技术的分析结果保持一致[28, 29], 这也揭示了国产披碱草属六倍体物种均为StHY基因组组成的物种。
 | 图4 供试六倍体物种种质资源的遗传结构Fig.4 The genetic structure analysis of the hexaploid species |
本研究对小麦族St、H、Y不同基因型组成的六倍体物种进行分析, 遗传相似系数及聚类分析等结果表明其具有相同或相似的基因组组成, 物种之间亲缘关系较近, 不同属但具有相同基因组组成物种能够聚在一起。例如具有StHY基因型组成的物种R. aristiglumis原属于鹅观草属且每穗轴为单小穗, 但与披碱草属StHY物种E. nutans和E. submuticus表现出较近的亲缘关系。种间关系的研究结果也给传统的以形态学为标准的披碱草属、鹅观草属等的种属间界限提出了异议[13], 且作者在野外资源采集过程中也会发现一些属于鹅观草属的物种每穗轴节上含有2个小穗的基因型出现。本研究结果也揭示了基于基因组组成的分类体系可能更能反映其遗传亲缘关系和系统分类地位, 但是仍有少部分具有相同基因组组成的物种却表现出较远的亲缘关系, 聚类不明显, 如本研究中来自巴基斯坦的E. himalayanus、来自伊朗的E. transhyrcanus等材料。这可能主要由于St、H基因组的不同供体物种参与了披碱草属、鹅观草属异源多倍化的进程, 具有多个起源, 且其基因组分化较明显[12, 27]。此外, 不同的地理来源和生境也可能造成这种大的种间遗传分化以及形成不同的地理分化类型。在披碱草属物种的种内就存在明显的地理分化, 如分子标记及细胞学证据均表明垂穗披碱草(E. nutans)存在广泛的遗传变异和地理分化, 来自青藏高原与新疆的材料间的分化非常明显, 青藏高原地区也存在不同的分化类型[15, 29]。
供试材料中具有StHY基因组的物种表现出了3类明显的分化类型, 即E. dahuricus(StHY-A)、E. nutans(StHY-B)、R. kamoji(StHY-C)类型, 同时其分化类型同形态学特征相吻合, 这也与已经报道的关于披碱草属StHY物种分化情况相一致。李永祥等[17]采用分子标记揭示了国产披碱草属六倍体物种中直穗类型与垂穗类型之间的分化。Dou等[20]采用原位杂交技术分析了1个四倍体E. sibiricus及5个六倍体物种E. nutans、E. barystachyus、E. xiningensis、E. excelsus、E. dahuricus的分子核型变异, 结果将其划分为E. sibiricus类、E.nutans类及E. dahuricus类。Sasanuma等[18]基于扩增片段长度多态性标记(amplified fragment length polymorphism, AFLP)结果揭示了来自日本的E. tsukushiensis、E. humidus和东亚的E. dahuricus 3个StHY基因组物种的分化, 形态上相似的E. tsukushiensis和E. humidus之间的分化程度较低并具有较近的亲缘关系, 它们与E. dahuricus之间的分化程度较高, 亲缘关系也较远。本研究在系统分析StHY基因组物种遗传亲缘关系的基础上, 提出了关于StHY基因组物种的3种分化类型, 但具有StHY基因组组成的E. schrenkianus物种也表现出了与这3种分化类型的差异, 且更偏向与StStY基因组物种较近的亲缘关系。这也可能与本研究采用的标记数目及地理来源等有关, 所以关于其分类地位及系统关系有待进一步分析。
在披碱草属六倍体物种中, 直穗类型物种E. dahuricus、E. tangutorum、E. excelsus、E. cylindricus、E. purpuriaristatus等物种具有非常相似的形态学特征和较近的亲缘关系, 即披碱草复合类群(E. dahurics complex), 其复合类群物种间的种间关系和界限也一直是研究的焦点。划为曲穗草属后, Baum等[14]将E. dahuricus复合群体中的其他几个种处理为C. dahuricus的几个变种, 即C. dahuricus var. excelsus、C. dahuricus var. tangutorum、C. dahuricus var. cylindrica等。Agafonov 等[30]利用蛋白质电泳、DNA分子标记及种间杂交染色体配对分析也揭示了E. dahuricus复合群体的分化, 同时表明了它们是具有广泛地理分布和变异的多态性物种, 并建议将其降级为种以下的分类群。在本研究结果中E. dahuricus复合群体几个物种明显聚在一起, 表现出了较近的亲缘关系, 甚至个别物种的种间界限不是很明显, 所以也支持将其分类等级降为变种处理。
本研究采用SSR分子标记分析了小麦族St、H、Y不同基因组组成的六倍体物种, 研究结果揭示了供试的不同基因组组成的六倍体物种之间具有丰富的遗传多样性和明显的种间分化, 其中StHY物种明显分化成3种类型; 相同或相似基因组组成的物种间表现出较近的亲缘关系, 本研究结果支持基于基因组组成的分类体系对该类群物种分类地位的划分。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|