




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
过量锰处理对苗期柱花草生长及抗氧化酶活性的影响
[刘攀道1  , 罗佳佳
, 罗佳佳1, 2 , 白昌军1 , 陈志坚1, * , 刘国道1, * ]
, 罗佳佳, 刘国道]
|
|


作者简介:刘攀道(1988-),男,云南腾冲人,助理研究员,博士。E-mail: liupandao@foxmail.com
以2份柱花草基因型(TF210和TF226)为材料,分析过量锰处理对苗期柱花草生长、叶绿素浓度(SPAD)、最大光化学效率( Fv/ Fm)、超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性的影响,探究柱花草中潜在的耐锰机制。结果表明,过量锰(400 μmol·L-1)处理显著抑制了柱花草基因型TF210地上部和根系的生长、降低了其叶片SPAD和 Fv/ Fm值,但对TF226生长影响不明显,说明TF226为耐锰基因型,而TF210为锰敏感基因型。虽然过量锰处理增加了两个柱花草基因型组织锰含量,但TF226地上部和根部的锰含量显著低于TF210,暗示在过量锰处理下,TF226积累了较少的组织锰含量。过量锰处理显著增加了两个基因型叶片SOD活性,但仅增加了TF226叶片POD和CAT活性,并且在过量锰处理下,TF226叶片SOD、POD和CAT活性值均高于TF210。以上结果说明维持根系生长、降低组织锰含量和提高抗氧化酶活性是苗期柱花草TF226耐锰的生理机制。研究结果为探索柱花草耐锰机理及耐锰柱花草品种改良提供了理论依据及种质材料。
The aim of this study was to investigate the potential mechanisms underlying stylo ( Stylosanthes guianensis) tolerance to high Mn levels. The growth responses and Mn tolerance in stylo were assessed in two stylo genotypes, TF210 and TF226 grown hydroponically. Plant dry weight, root growth parameters, chlorophyll concentration (soil plant analyzer development, SPAD), maximum photochemical efficiency ( Fv/ Fm) as well as superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT) activities were measured under two Mn levels. Results showed that significant decreases in shoots and roots growth, SPAD and Fv/ Fm values were observed in stylo genotype TF210 compared to TF226 under 400 μmol·L-1 MnSO4 treatment, suggesting that TF226 was a more Mn-tolerant genotype compared to TF210. Although Mn content in shoots and roots of both genotypes was increased by high Mn treatment, Mn content in TF226 was lower than that in TF210, suggesting that less Mn was accumulated in TF226 compared to TF210. Furthermore, SOD, POD and CAT activities in leaves of TF226 were significantly increased under high Mn treatment, while only SOD activity in leaves of TF210 was increased by high Mn. Activities of the three tested enzymes in TF226 were higher than those in TF210 under high Mn. These results suggest that regulations of root growth, Mn accumulation and antioxidant enzyme activities might be the adaptive mechanisms of stylo tolerance to Mn toxicity. The present study not only provides theoretical basis for dissecting Mn tolerant mechanisms, but also providing germplasm resources for improving adaptation to Mn toxicity in stylo.
锰是植物生长所必需的微量元素之一, 在植物生长发育和生命活动的代谢过程中起重要作用[1]。锰参与光合作用中电子传递系统的氧化还原和光系统II中水的光解过程[1]。锰还可参与ATP合成、脂肪酸和蛋白质的代谢过程[2]。在我国可耕作土壤中, 锰平均含量可达573 mg· kg-1, 并且, 我国土壤锰含量表现出南方酸性土壤锰含量高于北方的石灰性土壤[3, 4]。在酸性土壤中, 由于低氧化还原电势和低pH, 锰以可溶性的形态进入土壤溶液, 导致锰含量显著增加, 过量积累的锰离子会对植物细胞造成毒害, 抑制植物生长[5, 6]。
锰毒害症状主要表现在叶片上, 如出现褐色锰氧化斑、坏死斑和叶片失绿等[1]。锰毒害抑制叶片光合作用, 破坏多种重要代谢酶的活性, 产生活性氧自由基造成氧化胁迫, 影响根系对其他营养元素的吸收、运输和分配, 破坏根系组织结构[1, 7]。植物的耐锰机制主要包括把过量的锰积累在液泡和细胞壁等非活性部位、增强锰毒害下抗氧化能力、增加根系分泌物螯合根际锰离子减少根系对锰的吸收[6, 8]。不同植物品种或植物的不同基因型的耐锰能力不一样。
柱花草(Stylosanthes spp.)是主要的热带豆科牧草, 起源于热带和亚热带地区, 可作为饲料喂养牲畜、作为绿肥覆盖果园以及培肥和改良土壤等, 具有重要的经济价值和生态效益。一般认为, 起源于酸性土壤的热带植物表现出较强的适应酸性土壤生长的能力, 其可能具有独特的耐酸铝和酸锰胁迫的机制[9], 因此, 柱花草是研究酸性土壤锰毒害胁迫机制的良好材料。研究表明, 叶片失绿和皱缩是柱花草锰毒害的主要症状[5]。并且, 增加柱花草根系苹果酸的合成与分泌, 可以减少根系对锰的吸收, 进而缓解锰毒害[10]。目前, 已有针对锰毒胁迫影响柱花草生长、元素平衡等方面的研究, 但是, 柱花草响应锰毒害是一个复杂生理过程, 对不同基因型及不同生长期柱花草响应锰毒害的研究较少, 柱花草是否可以通过维持根系生长及提高抗氧化胁迫的能力缓解锰毒害仍不清楚。因此, 本研究以2份柱花草基因型为材料, 分析不同锰浓度处理对苗期柱花草根系生长和抗氧化酶活性等的影响, 以期为探索柱花草耐锰机理及耐锰柱花草品种改良提供理论依据及种质材料。
本实验所用的2份圭亚那柱花草(Stylosanthes guianensis)基因型分别为TF210和TF226。柱花草种子由中国热带农业科学院热带作物品种资源研究所热带牧草研究室提供。
1.2.1 实验处理 参照陈志坚等[5]方法, 柱花草种子去除外种皮后, 80 ℃加热2 min, 冷却后于黑暗条件下催芽2~3 d, 然后, 柱花草幼苗移栽至14 L塑料盆中于温室进行营养液培养。试验于2017年4月进行。培养温度为28 ℃/25 ℃(昼/夜), 湿度75%, 光照强度800 μ mol· m-2· s-1。营养液为1/2 Hoagland溶液, 每7 d更换1次营养液, 每2 d用KOH或H2SO4调节pH值至5.8。参照Chen等[10]方法, 幼苗生长1个月后, 移栽至含有不同锰浓度的1/2 Hoagland营养液中处理7 d。锰浓度分别为5 μ mol· L-1(对照)和400 μ mol· L-1 MnSO4(过量锰处理), 营养液pH为5.0, 每个处理设置4个重复。每2 d用KOH或H2SO4调节pH值至5.0。处理7 d后, 对柱花草活体叶片进行叶绿素和最大光化学效率(Fv/Fm)测定。对一部分植株分别收取地上部和根部样品, 根系用去离子水冲洗干净, 用于根系参数分析后, 地上部和根部样品在75 ℃下烘干至恒重, 称取干重; 另一部分植株叶片样品用液氮速冻, 然后置于-80 ℃超低温冰箱保存, 用于酶活性分析。
1.2.2 测定方法 根系用EPSON 1640XL扫描仪(EPSON, 日本)进行扫描, 总根长和根体积用WinRhizo软件(Regent Instruments, 加拿大)进行分析。采用SPAD502型叶绿素分析仪测定叶绿素含量, 用便携式荧光仪Pocket PEA(Hansatech, 英国)测定叶片最大光化学效率(Fv/Fm)。柱花草叶片超氧化物歧化酶(superoxide dismutase, SOD), 过氧化物酶(peroxidase, POD)和过氧化氢酶(catalase, CAT)活性采用试剂盒进行测定, 试剂盒购买于苏州科铭生物技术有限公司。以上每个指标测定设置4个重复。
采用Microsoft Excel 2003作图, 用SPSS 20.0(SPSS Institute, 美国)软件进行方差分析。
如图1所示, 相对对照(5 μ mol· L-1)处理, 过量锰(400 μ mol· L-1)处理均显著抑制了柱花草基因型TF210的地上部(F=9.2, P< 0.05)和根部(F=9.2, P< 0.05)干重。在过量锰处理下, TF210地上部和根部干重相对对照分别降低了27.1%和17.6%, 差异显著(P< 0.05)(图1)。但是, 过量锰处理对基因型TF226地上部和根部干重影响不明显(图1)。
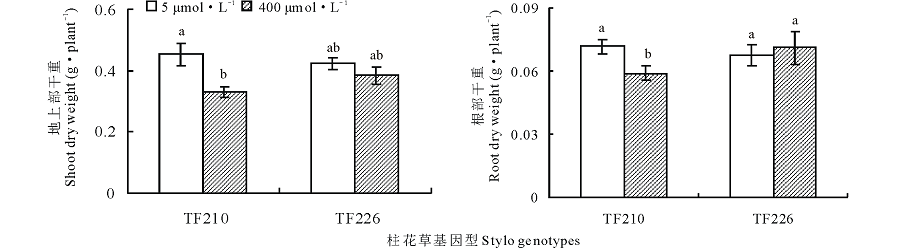 | 图1 不同锰处理对柱花草地上部和根部干重的影响不同小写字母表示同一柱花草种质在不同处理间差异显著(P< 0.05), 下同。Fig.1 Shoots and roots dry weight of stylo at different Mn treatmentsDifferent letters indicated significant difference among different treatments at P< 0.05 level in the same stylo accession, the same below. |
从图2可以看出, 过量锰(400 μ mol· L-1)处理显著抑制了柱花草基因型TF210的主根长(F=5.9, P< 0.05)和总根长(F=10.5, 0.01< P< 0.05), 但对根体积的影响不明显(F=3.7, P> 0.05)。在过量锰处理下, TF210主根长和总根长相对对照分别降低了14.0%和31.5%(图2)。但是, 过量锰处理对基因型TF226主根长、总根长和根体积均无显著影响(图2)。因此, 在过量锰处理下, 基因型TF226能够维持根系生长。
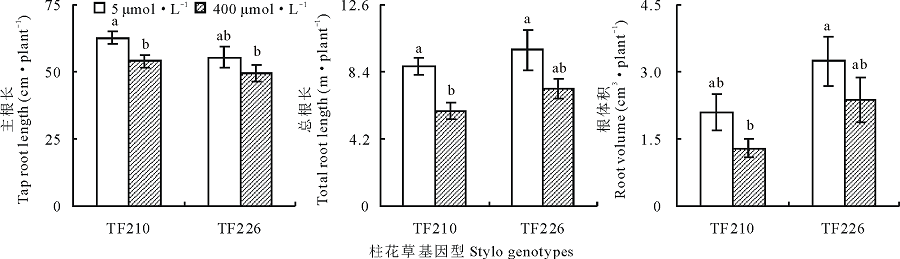 | 图2 不同锰处理对柱花草根系生长的影响Fig.2 Effects of different Mn levels on root growth of two stylo genotypes |
过量锰(400 μ mol· L-1)处理显著降低了柱花草叶绿素浓度(SPDA)(F=36.4, P< 0.001)(图3)。在过量锰处理下, TF210和TF226的SPDA值相对对照分别降低了35.8%和20.0%, 并且, 在过量锰处理下, 基因型TF226的 SPDA值显著高于TF210(P< 0.05)(图3)。
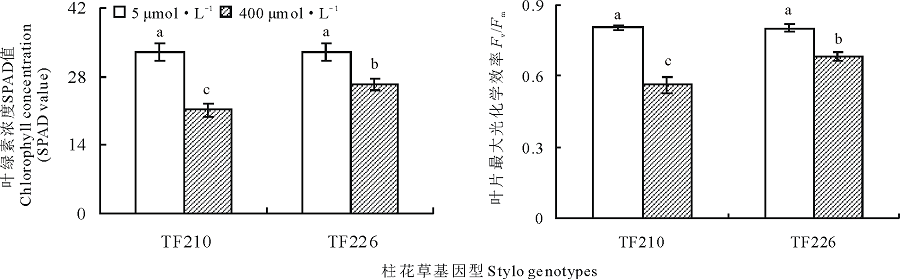 | 图3 不同锰处理对柱花草叶绿素浓度及Fv/Fm的影响Fig.3 Chlorophyll concentration and Fv/Fm in the leaves of two stylo genotypes at different Mn treatments |
过量锰处理显著影响柱花草最大光化学效率(Fv/Fm)(F=66.8, P< 0.001), 并且, 锰处理对Fv/Fm的影响具有基因型差异(F=7.3, P< 0.05), 锰处理和基因型两因素对Fv/Fm的交互影响显著(F=7.8, P< 0.05)(图3)。在过量锰处理下, TF210和TF226的Fv/Fm值相对对照分别降低了30.1%和14.8%, 并且, 在过量锰处理下, 基因型TF226的Fv/Fm值显著高于TF210, TF226的 Fv/Fm值是TF210的1.2倍(图3)。因此, TF226是耐锰基因型, 而TF210是锰敏感基因型。
过量锰处理显著增加了柱花草地上部组织锰含量(F=225.9, P< 0.001)(图4)。在过量锰处理下, TF210和TF226地上部锰含量相对对照分别增加了15.7和15.3倍(图4)。类似的, 过量锰处理显著增加了柱花草根部组织锰含量(F=1963.7, P< 0.001), 并且, 锰处理对根部组织锰含量的影响具有基因型差异(F=76, P< 0.001), 锰处理和基因型两因素对根部组织锰含量的交互作用显著(F=40.3, P< 0.001)(图4)。在过量锰处理下, 虽然TF210和TF226根部锰含量相对对照分别增加了8.7和10.9倍, 但是, 基因型TF210的根部锰含量显著高于TF226, TF210根部锰含量是TF226的 1.4倍(图4)。此外, 在过量锰处理下, TF210和TF226根部锰含量分别是地上部锰含量的1.7和1.5倍(图4)。
 | 图4 不同锰处理对柱花草地上部和根部锰含量的影响Fig.4 Mn content in shoots and roots of stylo at different Mn treatments |
从图5可以看出, 不同锰浓度处理影响柱花草叶片抗氧化酶(SOD, POD和CAT)活性, 并且, 锰处理对抗氧化酶活性的影响具有基因型差异。过量锰(400 μ mol· L-1)处理显著增加了柱花草SOD活性(F=129.7, P< 0.001)。相对对照(5 μ mol· L-1)处理, TF210和TF226在过量锰处理下的SOD活性分别增加了63.3%和80.5%(图5), 并且, 在过量锰处理下, TF226的SOD活性显著高于TF210(F=11.1, P< 0.05)。
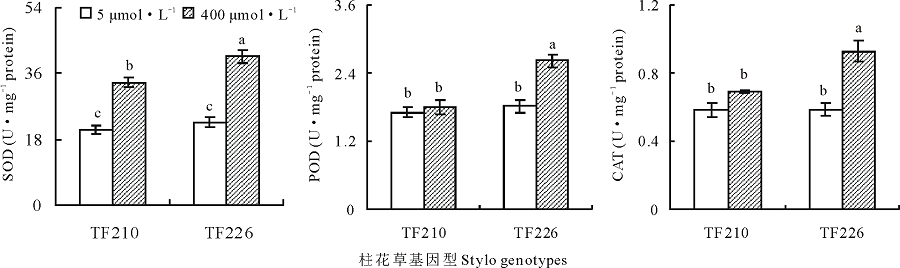 | 图5 不同锰处理对柱花草叶片SOD, POD和CAT活性的影响Fig.5 SOD, POD and CAT activities in the leaves of two stylo genotypes at different Mn treatments |
锰处理和基因型两因素对POD(F=10.6, P< 0.05)和CAT(F=7.7, P< 0.05)活性的交互影响显著(图5)。过量锰处理显著增加了耐锰基因型TF226的POD和CAT活性(P< 0.05), 但对TF210的POD和CAT活性影响不明显。在过量锰处理下, TF226的POD和CAT活性相对对照分别增加了44.0%和58.6%(图5)。并且, 在过量锰处理下, TF226的POD和CAT活性分别是TF210的1.5和1.3倍(图5)。
锰毒害是酸性土壤中限制作物生长和产量的重要因素之一。起源于酸性土壤的植物能够形成独特的耐酸锰酸铝胁迫的机制, 从而表现出超强的适应酸性土壤的能力[9]。通过比较分析12种不同豆科植物的耐锰能力, 包括大豆(Glycine max)、菜豆(Phaseolus vulgaris)、苜蓿(Medicago sativa)和柱花草等, 发现在这些豆科植物中, 柱花草具有较强的耐锰能力[11]。在豇豆(Vigna unguiculata)、大麦(Hordeum vulgare)和大豆中, 50 μ mol· L-1锰浓度处理会导致叶片形成褐色锰斑点的锰毒害症状, 并抑制了植物的生长[12, 13, 14]。研究发现, 柱花草在低于200 μ mol· L-1的锰浓度处理下并没有出现锰毒害的症状, 只有在400 μ mol· L-1的锰浓度处理下才显著降低了柱花草锰敏感基因型TPCR2001-1叶片的叶绿素浓度和植株干重, 说明柱花草锰毒害表现出基因型差异[10]。因此, 本研究以2份柱花草基因型(TF210和TF226)为材料, 分析过量锰(400 μ mol· L-1)处理对苗期柱花草生长的影响及其可能的耐锰机制。
在水稻(Oryza sativa)、小麦(Triticum aestivum)和大麦等植物中, 锰毒害所产生的氧化胁迫破坏了叶绿体结构、影响光合作用的电子传递过程, 进而抑制植物的生长, 而耐锰的植物或基因型则表现出能够维持较强的光合作用和植株生长能力[14, 15, 16]。在本研究中, 过量锰处理显著抑制了柱花草基因型TF210的地上部和根部干重以及根系生长, 降低了TF210叶片SPAD和Fv/Fm值, 但对基因型TF226生长影响不明显(图1~3), 说明TF226为耐锰基因型, 而TF210为锰敏感基因型。值得注意的是, 过量锰处理显著抑制了柱花草基因型TF210的主根长和总根长, 但对TF226根系生长影响不明显(图2)。类似的, 过量锰处理会显著抑制大豆、水稻和小麦根系的生长[15, 16, 17]。研究发现, 在过量锰处理下, 小麦根系可以通过调节其他矿质元素(如镁、钙、铁和磷等)的吸收和分布, 从而影响小麦的耐锰能力[16]。因此, 在过量锰处理下, 通过维持根系的正常生长, 可能有助于TF226根系对其他养分的吸收和运输, 从而缓解TF226锰毒害。
在本研究中, 尽管过量锰处理显著增加了柱花草组织锰含量, 但耐锰基因型TF226地上部和根部的锰含量显著低于TF210(图4), 这与Chen等[10]研究结果相似, 说明降低组织锰的积累可能是TF226耐锰的机制之一。另一方面, 在过量锰处理下, 虽然TF226地上部和根部锰含量显著低于TF210, 且未表现出锰抑制生长的明显症状, 但是过量锰处理仍然显著增加了TF226地上部和根部的锰含量(图4)。研究发现, 在一些植物中, 细胞中累积的锰, 可能被有机酸等螯合并储存在液泡中[18, 19, 20]。例如, 在豇豆中, 植株内部80%的锰离子可以被柠檬酸所螯合[21]。研究发现, 过量锰处理显著增加了柱花草耐锰基因型Fine-stem叶片和根部的内源苹果酸浓度, 增加合成的苹果酸可能进一步形成锰-苹果酸复合物, 从而缓解了锰毒害[10]。因此, 柱花草基因型TF226组织中的锰离子可能被某些物质螯合(如有机酸), 并隔离在液泡中, 从而未表现出锰毒害的症状。
在柱花草中, 组织中锰离子的过量积累, 会产生活性氧自由基(reactive oxygen species, ROS)造成氧化胁迫, 破坏细胞内多种重要代谢酶的活性, 从而抑制叶片光合作用, 进而影响了柱花草植株和根系的生长。为了适应锰毒害, 植物可以通过增强抗氧化能力(抗氧化酶活性和抗氧化物质含量)提高抵抗锰毒害的能力[13, 15, 22]。研究发现, 在过量锰诱导的氧化胁迫下, 水稻幼苗可以通过增强SOD, CAT和APX(抗坏血酸过氧化物酶)等酶活性, 清除组织内积累的活性氧自由基, 从而缓解锰毒害[15]。并且, 在锰毒害下, 增强POD活性及其蛋白表达, 被认为是豇豆响应锰毒害的重要机制[22]。在本研究中, 过量锰处理显著增加了耐锰基因型TF226叶片SOD, POD和CAT活性(图5), 这些抗氧化酶活性的增加, 可能在降低由锰毒害造成的氧化胁迫中起重要作用, 从而增强了TF226耐锰能力。
综上所述, 在柱花草中, 锰毒害下过量积累锰离子, 会对细胞产生氧化胁迫, 破坏叶绿素合成及光合作用, 进而抑制柱花草植株生长。相对柱花草基因型TF210, TF226具有较强的耐锰能力, 是耐锰基因型。在锰毒害下, 基因型TF226可能通过减少锰的积累并提高抗氧化酶活性, 降低由锰毒害造成的氧化胁迫伤害, 从而保持较高的叶绿素浓度和最大光化学效率, 进而维持了地上部和根系的生长。本研究结果为探索柱花草耐锰机理及耐锰柱花草品种改良提供了理论依据及种质材料。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|