

{kind=link}
{kind=link}
{kind=link}
黄土丘陵区不同植物凋落叶片的分解及养分释放特性
[刘晶 , 谢婉余, 张巧明, 徐少君]
, 谢婉余, 张巧明, 徐少君]
, 谢婉余, 张巧明, 徐少君]
|
|


作者简介:刘晶(1977-),女,吉林松原人,博士。E-mail: ccliujing@163.com
叶片凋落物的分解对生态系统的养分循环具有重要意义。刺槐、油松和柠条是豫西黄土丘陵区退耕还林时普遍采用的树种,高羊茅是该区分布广泛的建种草本植物。采用分解网袋法,对该区 4 种植物叶片凋落物一年内的分解特征及营养元素的释放规律进行研究。结果表明:1)柠条叶的分解率分别比刺槐叶、油松和高羊茅高8.86%、19.87%和13.18%,分解规律符合Olson 指数模型,分解速率为柠条叶>刺槐叶>高羊茅叶>油松叶。2)分解过程中 4 种凋落叶片的C/N、P含量持续降低,木质素含量缓慢增加,纤维素含量呈先增加后降低的变化趋势;不同凋落叶片分解过程中全C和全N含量的变化趋势存在差异。4 种凋落叶片的C、P元素属于直接释放模式,N元素属于富集—释放模式。3)凋落叶片的分解速率与总C含量、木质素含量和C/N均呈显著负相关关系( P<0.01),因此该地区凋落叶片的总C、木质素含量和C/N可以作为预测该地区凋落物分解的指标,凋落物叶片功能特征(比叶面积、厚度、硬度等)可能是影响其分解速率的重要因素之一。
The decomposition of leaf litter has important significance for nutrient cycling in ecosystems. Robinia pseudoacacia, Pinus tabulaeformis and Festuca arundinacea are commonly used species for returning farmland to forests in the Loess Hilly Region of western Henan, and Caragana korshinskii is a widely distributed herbaceous plant in this region.We studied the differences in decomposition characteristics of R. pseudoacacia, P. tabulaeformis, F. arundinacea and C. korshinskii leaf litter, using a nylon bag method. Leaf litter of F. arundinacea decomposed, respectively, 8.86%, 19.87% and 13.18% faster than litter of R. pseudoacacia, P. tabulaeformis and C. korshinskii. Leaf litter decomposition processes followed Olson’s exponential model. Decay rates for leaf litter of the 4 species ranked F. arundinacea> R. pseudoacacia> C. korshinskii> P. tabulaeformis. During the decomposition process, the C/N and P content the leaf litter of 4 litters species decreased gradually but the lignin content increased slowly, while the cellulose content initially increased and then decreased. Change with time in C and P concentration of decomposing leaf litter followed a ‘direct release’ model, while N concentration followed an ‘enrichment-release’ model. The concentrations of total C, and lignin and the C/N ratio were significantly, negatively correlated with decomposition rate of the leaf litters ( P<0.01).Therefore, the concentrations of total C, lignin and C/N ratio in leaf litter samples could be used as decomposition indicators to predict litter decay rate in this region. We also noted that physical characteristics of leaf litter (specific leaf area, thickness, hardness, etc.) are additional factors with important effects on leaf litter decomposition rate.
凋落物是植物在生长发育过程中新陈代谢的产物, 作为养分的基本载体, 在养分循环中是连接植物与土壤的“ 纽带” , 它的生产和分解是生态系统物质循环的主要途径[1, 2]。土壤是凋落物的载体, 凋落物分解过程中养分的释放对保持土壤肥力、增强土壤结构稳定性、改善土壤生物活性具有重要作用[3, 4]。凋落物分解是陆地生态系统碳库循环的一个重要环节, 全球每年因凋落物分解释放的二氧化碳量约占全球年碳通量70%[5, 6], 但至今, 凋落物对土壤有机碳的影响机制尚不十分清楚。研究表明凋落物分解速率的大小影响着它向土壤释放营养元素的速率, 在环境条件一致的条件下, 凋落物分解及其养分释放主要受凋落物初始化学组成调控, 尤其是凋落物中木质素和氮素含量是影响凋落物分解速率的重要因素[7, 8, 9]。然而, 由于不同凋落物种类的化学组成存在差异, 影响凋落物分解过程的因素又极其复杂, 再加上研究方法的不一致性和养分释放机制的多样性, 导致各地的研究结果差异很大[10, 11, 12], 至今未能获得可普遍应用的凋落物分解的预测指标。
结合退耕还林工程, 国内研究者对安塞、长武、陇中等黄土高原地区不同植被凋落物的性质和分解进行了大量研究报道[13, 14, 15, 16]。但黄土丘陵沟壑区地形破碎, 地理环境复杂, 区域差异很大。豫西黄土丘陵区是河南省水土流失最严重的地区之一, 也是国家最早确定的退耕还林(草)试点地区之一。该区土质疏松, 地力瘠薄, 因此退耕后凋落物的分解对改善土壤理化性质、促进植被生长, 减少该区水土流失尤为重要。刺槐(Robinia pseudoacacia)、油松(Pinus tabulaeformis)、柠条(Caragana korshinskii)是豫西黄土丘陵区退耕还林普遍采用的树种, 高羊茅(Festuca arundinacea)群落则是该区分布较为广泛的建群种草本群落之一, 对以上几种植物凋落物的性质及分解速率进行研究, 可以为该区生态恢复中植被的选择与配置、生态恢复效果的评估提供依据, 也能够为该区生态恢复对碳排放的源汇效应提供基础数据。
试验地位于新安县境内, 为豫西黄土丘陵典型地貌区。刺槐、油松人工林均为2000年退耕后种植, 株行距1.5 m× 1.5 m, 林下伴生少量茅莓(Rubus parvifolius)等小灌丛。柠条为2003年退耕后种植, 株行距1 m× 1 m, 伴生达乌里胡枝子(Lespedeza davurica)等植物。高羊茅草地为2006年弃耕后自然恢复形成, 高羊茅为建群种, 伴生少量狗尾草(Setaria viridis)、长芒草(Stipa bungeana)。
样品的采集采用随机多点取样, 在每个样地内设置10个1 m× 1 m的样方, 每个样方用高30 cm黑色尼龙网围住, 边角用铁丝固定。2014年3月中旬将所有样方内地面上的凋落物清除干净, 以后每隔2个月收集一次刺槐、油松和柠条地样方内的凋落物, 至2015年3月结束; 高羊茅在2014年10月下旬刈割其整个地上部分。刺槐、油松、柠条和高羊茅凋落叶片分别标记为R、P、C、F。每次收集的凋落物带回室内后置于65 ℃烘箱内烘至恒重后称量。将烘干的植物凋落物取部分粉碎过筛(0.5~1.0 mm)后, 测定凋落物的碳、氮、磷、半纤维素、纤维素和木质素含量。
称取凋落叶片15 g, 装入规格为20 cm× 30 cm、孔径为1 mm的尼龙网袋中, 用尼龙线封口, 于2015年10月20日放入原取样地内(每个样方附近放置1个样袋), 放置时贴近表土, 尽量与周围凋落物处于同一自然状况。每两个月定期取出样袋, 抖落泥土和杂质, 称重后再置于65 ℃的烘箱内烘干24 h, 测定凋落物剩余干物质的重量, 之后放回原地。2016年2月20日、6月20日和10月底, 分别取出各类凋落物3个样袋, 测定残留凋落物的相关养分含量。
凋落物中全C含量用重铬酸钾容量法测定[17]; 全N含量用凯氏法测定[17]; 全磷含量用钼锑抗比色法测定[17]; 半纤维素、纤维素和木质素含量用酸性洗涤纤维法测定[18, 19]。
凋落叶片分解率:D=
式中:W0为凋落叶片初始质量; Wt为分解t时间凋落叶片残留量。
凋落叶片养分释放率: R=
式中:R为养分释放率; W0为凋落叶片初始质量; C0为初始养分浓度; Wt为分解t时间凋落叶片残留量; Ct为分解t时间凋落叶片的养分浓度。
采用Olson指数模型计算凋落叶片分解速率:
式中:Xt为时间t时的干质量; X0为凋落物的初始干质量; k为凋落物的年分解速率。依据各月的剩余率数据, 拟合凋落物的分解速率曲线, 并估算凋落物分解的半衰期t0.5和周转期t0.95。
采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)比较不同凋落物初始化学元素含量的差异, 相关分析均采用SPSS 18.0完成。
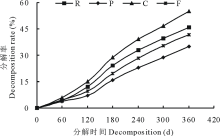
刺槐叶、油松叶、柠条叶和高羊茅凋落叶片分解率的动态变化见图1, 4种凋落叶片在不同分解阶段的分解率存在明显差异。在整个分解过程中, 各凋落叶片的分解率始终为:柠条叶> 刺槐叶> 高羊茅叶> 油松叶。经过360 d的分解, 刺槐叶、油松叶、柠条叶和高羊茅凋落叶片分解率分别为45.98%、34.97%、54.84%和41.66%, 其中柠条叶的分解率分别比刺槐叶、油松叶和高羊茅高8.86%、19.87%和13.18%, 表明柠条叶的分解速率明显快于油松和高羊茅凋落物。
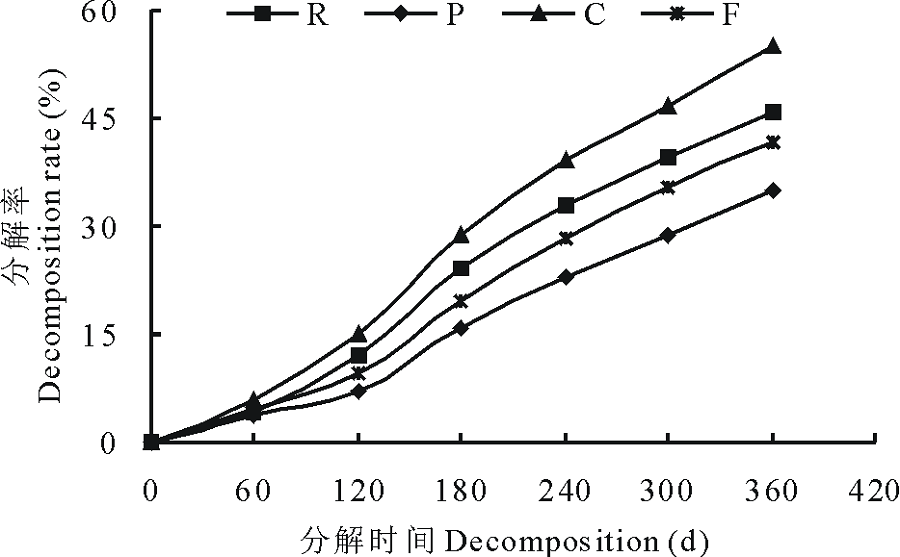 | 图1 不同凋落叶片分解率的变化R:刺槐凋落叶Leaf litters of R. pseudoacacia; P:油松凋落叶Leaf litters of P. tabulaeformis; C:柠条凋落叶Leaf litters of F. arundinacea; F:高羊茅凋落叶Leaf litters of C. korshinskii. 下同The same below.Fig.1 Decomposition rate change of different leaf litters |
依据各月的残留率数据, 采用Olson指数衰减模型拟合各凋落叶片的分解速率曲线, 并估算凋落叶分解的半衰期t0.5和周转期t0.95。由表1可知, 4种凋落叶片的Olson指数模型的相关系数均达到极显著水平, 说明拟合效果良好。平均分解率也即分解常数, 是表征凋落物分解速率的重要指标。刺槐、油松、柠条和高羊茅凋落叶片的平均分解率分别为0.912、0.814、0.964和0.902年; 油松凋落叶的平均分解率最小, 柠条凋落叶的平均分解率最大。油松凋落叶的平均分解速率分别为刺槐、柠条和高羊茅凋落叶的0.89、0.84和0.90倍。柠条凋落叶的半衰期t0.5和周转期t0.95分别为0.72和2.24年, 油松凋落叶的t0.5和t0.95分别为1.38和5.86年, 油松凋落叶分解50%所用的时间比刺槐、柠条和高羊茅凋落叶分别多0.54、0.66和0.47年, 分解95%所用的时间则分别多2.44、3.62和1.68年。研究结果表明, 4种凋落叶的平均分解率为柠条凋落叶> 刺槐凋落叶> 高羊茅凋落叶> 油松凋落叶, 半衰期t0.5和周转期t0.95为柠条凋落叶< 刺槐凋落叶< 高羊茅凋落叶< 油松凋落叶, 因此该地区4种凋落叶分解速率大小表现为柠条凋落叶> 刺槐凋落叶> 高羊茅凋落叶> 油松凋落叶。
| 表 1 不同凋落叶片的分解速率 Table 1 The decomposition rate of different leaf litters |
凋落叶片的主要化学性质及分解过程中各种养分含量如图2, 可以看出, 不同凋落叶片之间全C、全N、全P、半纤维素、纤维素和木质素含量存在显著差异(P< 0.05)。油松凋落叶的全C和木质素含量最高, 分别为491.27和198.92 g· kg-1, 半纤维素和纤维素含量最低, 分别为69.43和178.58 g· kg-1; 刺槐凋落叶的全C最低为392.18 g· kg-1, 柠条凋落叶的木质素含量最低为104.25 g· kg-1; 油松凋落叶的全C和木质素含量显著高于柠条和刺槐凋落叶, 半纤维素和纤维素含量显著低于柠条和刺槐凋落叶(P< 0.05)。柠条凋落叶的全N含量最高, 为14.63 g· kg-1, 显著高于刺槐、油松和高羊茅凋落叶的全氮含量(P< 0.05)。刺槐凋落叶的全磷含量最高为3.24 g· kg-1, 油松凋落叶的全磷含量最低为1.78 g· kg-1, 刺槐凋落叶的全P含量显著高于柠条叶和油松凋落叶(P< 0.05), 但与高羊茅凋落叶之间无显著差异(P> 0.05)。
在0~360 d的分解过程中, 刺槐、油松、柠条和高羊茅凋落叶片中P含量呈逐渐降低, 木质素含量呈缓慢增加的趋势, 半纤维素含量在分解0~120 d呈缓慢降低趋势, 120 d后含量急剧降低, 至分解结束, 各凋落叶片半纤维素含量无显著差异(P> 0.05)。在整个分解过程中, 各凋落叶片全C、全N、纤维素含量变化差异较大。整个分解过程中油松凋落叶的全C含量一直为最高, 刺槐凋落叶的全C含量最低, 油松凋落叶在分解过程中全C含量呈现增加-降低-增加的变化趋势, 刺槐和柠条凋落叶呈现增加-降低的变化趋势, 高羊茅凋落叶的全C含量持续降低。油松和高羊茅凋落叶在分解过程中全N含量呈现持续增加的变化趋势, 柠条和刺槐凋落叶的全N含量表现为先增加后降低。柠条、刺槐和高羊茅凋落叶纤维素含量呈现先增加后降低的变化趋势, 油松凋落叶的纤维素含量则一直增加。整个分解过程中, 4种凋落叶片的P和纤维素含量持续降低, P含量刺槐凋落叶最高, 油松凋落叶最低; 木质素含量油松凋落叶最高。4种凋落叶片的C/N虽然有一定差异, 但变化趋势相同, 在整个分解过程中, 几种凋落叶的C/N都呈现出下降的趋势, 油松凋落叶的C/N最大, 柠条凋落叶的C/N最小, 至分解实验结束, 油松凋落叶的C/N为31.34, 显著高于刺槐、柠条和高羊茅凋落叶片的C/N(P< 0.05)。
刺槐、油松、柠条和高羊茅4种凋落叶片在分解过程中养分释放率的动态变化见图3。总体来看, 分解过程中4种凋落叶片中C和纤维素的释放率变化模式较接近, 二者都处于净释放状态且分解前期(0~120 d)释放较慢, 后期(120~360 d)稳定增加; P和半纤维素的释放率变化模式接近, 二者也都处于静释放状态且在整个分解过程中释放率持续增加; N和木质素释放率变化趋势相近, 二者在分解前期(0~120 d)表现为富集状态, 随分解时间延长, 凋落物中N和木质素基本上呈单调净释放状态。至分解结束时, 不同凋落叶片各元素释放率存在一定的差异。半纤维素的释放率最高, 分解结束时, 柠条叶凋落物C和纤维素释放率最高, 分别为55.63%和59.79%, 显著高于油松凋落叶C和纤维素释放率(P< 0.05), 分别为40.02%和25.56%, 柠条凋落叶C释放率与刺槐和高羊茅凋落叶片之间无显著差异(P> 0.05), 但纤维素释放率显著高于刺槐和高羊茅凋落叶片(P< 0.05); 柠条凋落叶P的释放率最高为67.41%, 显著高于油松凋落叶P的释放率; 4种凋落叶片的半纤维素释放率都在90%以上, 且各凋落叶片之间无显著差异(P> 0.05); 各凋落叶分解过程中N的释放率差异最大, 整个分解过程中, 柠条凋落叶N呈现净释放状态, 油松凋落叶正好相反, 呈现净富集状态, 刺槐叶和高羊茅凋落叶在分解的前期(0~120 d)表现为富集状态, 后期呈现释放状态。各凋落物木质素的释放率较低, 其中最高者为高羊茅凋落叶, 也仅有32.73%。
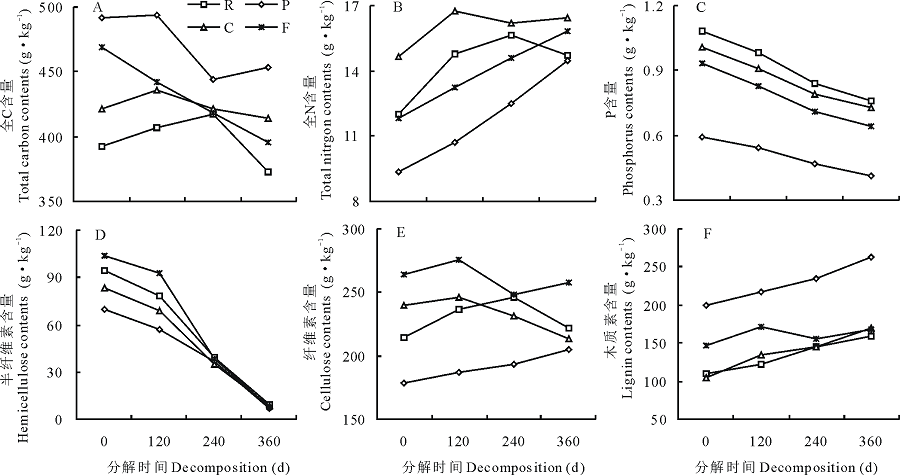 | 图2 凋落叶片分解过程中全C(A)、全N(B)、P(C)、半纤维素(D)、纤维素(E)和木质素(F)含量的动态变化Fig.2 The dynamic change of total C (A), total N (B), P (C), hemicellulose (D), cellulose (E) and lignin (F) contents in leaf litters during decomposition process |
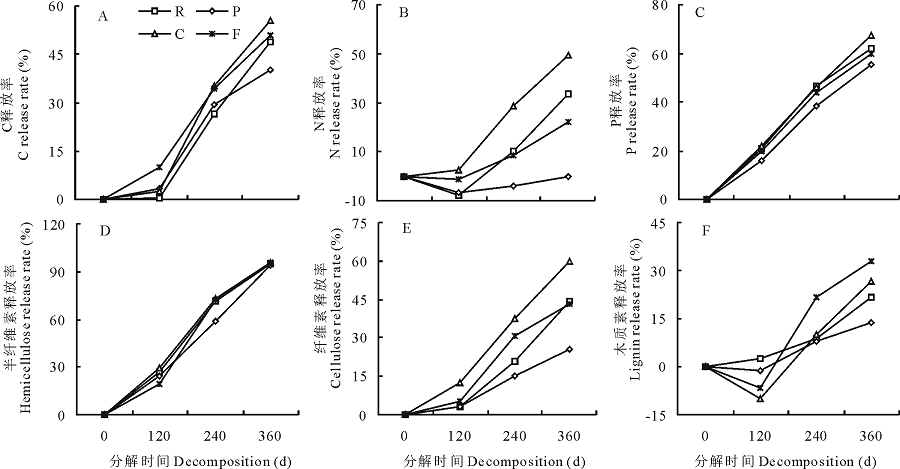 | 图3 凋落物分解过程中全C(A)、全N(B)、P(C)、半纤维素(D)、纤维素(E)和木质素(F)释放率的动态变化Fig.3 The dynamic change of total C(A), total N(B), P(C), hemicellulose(D), cellulose (E) and lignin (F) release rate in leaf litters during decomposition process |
4种凋落叶片分解速率与初始化学组成的相关关系如表2所示。刺槐、油松、柠条和高羊茅凋落叶片的分解速率与总C含量、木质素含量和C/N均显著负相关 (P< 0.01), 除油松凋落叶外, 其余3种凋落叶分解速率与全N、全P含量显著正相关(P< 0.05)。不同凋落叶片分解速率与初始半纤维素含量之间无显著相关性(P> 0.05)。
| 表2 不同凋落叶片分解速率与初始化学性质相关性 Table 2 Correlation between the decomposition rates and the initial chemical properties in different leaf litters |
本实验中开始120 d(10月20日至2月20日)凋落叶的分解较慢, 分解率较低; 中间120 d(2月20日至6月20日)分解加快, 后4个月分解又变慢, 但分解率逐渐提高, 这种变化规律也是外界环境因素和凋落叶片成分变化共同作用的结果。凋落物分解包括粉碎、淋溶和分解代谢过程, 是一个复杂的物理、化学和生物学过程。通常, 凋落物的分解分为两个阶段:第1阶段为快速分解阶段, 主要分解新鲜有机质、可溶性糖和无机盐类物质, 主要由原生动物、微生物的同化和异化作用以及土壤渗滤作用所致, 受环境因子的控制; 第2阶段为慢速分解阶段, 主要分解第1阶段残留凋落物的高C/N、高木质素/N的难分解有机质组分, 酚类物质和木质素等较难分解的成分, 分解速率主要受凋落物自身的理化性质及微生物组群的影响, 主要由真菌类的担子菌、半知菌、子囊菌以及少数种类的细菌和放线菌进行分解[5, 20, 21, 22]。分解试验开始时(11月), 凋落叶片中易分解物质的含量较高, 但由于该地区气温低, 降水少, 微生物活性降低, 不利于凋落叶片的分解, 因此分解较慢, 分解率较低; 从3月开始, 气温逐渐升高, 降水量逐渐增加, 有利于微生物的大量繁殖, 凋落叶的分解速率明显增加, 分解率提高, 尤其是在分解的第120~180天(2月20日至4月20日)内, 分解率提高很快; 分解后期(6月20日至10月20日)虽然该时段内温度和降水在本区也有利于微生物的生存, 但随着分解时间的延长, 一些易分解的物质已经分解掉, 难分解有机物质、纤维素、木质素等难以被微生物利用的大分子有机物比例增加, 即使环境条件更有利于凋落叶的分解, 但分解变慢, 分解率增加幅度变小。
凋落物在分解过程中养分释放主要有淋溶— 富集— 释放、富集— 释放和直接释放3种模式[23, 24]。C是组成有机质的主要元素, 为分解者提供可利用的能源, 前期分解过程中, 由于作为有机碳主要成分的纤维素、木质素还未分解, 有机碳损失量小于干物质损失量, 因此有机碳浓度表现为上升过程; 随着半纤维素、纤维素等有机物大量分解, 有机碳浓度开始降低。虽然不同凋落叶片分解过程中有机碳浓度呈现先升高后降低的变化趋势, 但整个分解过程中所有凋落叶片的有机C释放率均随分解时间的延长而增加, 因此可以认为凋落叶片C元素的释放模式为直接释放模式。凋落物C/N值是发生N固持和矿化的关键值。Parton等[25]研究表明当C/N小于40时才会发生净N的矿化, 而Patra等[26]认为当C/N值> 30时发生N的固持而当C/N值< 30时发生N的矿化。本研究中几种凋落物的初始C/N介于28.80~52.49之间, 但刺槐叶在分解的前120 d, 油松凋落物在分解的前240 d, N元素仍处于富集模式。理论上来讲C/N应该控制凋落物分解过程中的净N释放, 因为微生物分解者只有当其N需求被满足之后才可能释放N。较高C/N的凋落物在分解时自身N含量难以满足微生物(分解者)的N需求, 微生物则需要通过吸收外源N构成微生物量或胞外酶[27, 28, 29]。不同地区各类凋落物分解时主导的微生物不同, 对某种元素的需求阈值也就可能存在差异[18, 21]。就本研究而言, N元素总体仍属于富集— 释放模式, 并且油松凋落叶片的富集度最高, 其次是刺槐和高羊茅, 柠条最低。P的矿化释放与林分所处的环境条件及其初始含量有关。内蒙古典型草原区克氏针茅(Stipa krylovill)和羊草(Leymus chinensis)中P含量较低, P表现为先释放后富集; 糙叶黄芪(Astragalus scaberrimus)和星毛委陵菜(Potentilla acaulis)中P含量较高, P则一直表现为释放[30]。本研究中4种凋落叶片P元素的释放率在分解的0~360 d呈上升趋势, 由于本试验开始时间为11月, 研究区降水很少, 因此也可以认为P元素属于直接释放模式, 分解过程中未出现P富集及淋溶。
凋落物的基质质量是制约其分解的内在因素, 在特定的区域内, 凋落物的分解与其初始化学性质关系密切[21, 31]。不同植物的同一器官初始元素含量不同, 凋落物初始化学元素含量的差异可能会造成凋落物后续分解过程和养分释放模式的差异。研究表明亚热带地区不同植被恢复阶段植被凋落物的分解速率随初始N、P浓度增加而增加, 随初始木质素浓度、C/N和木质素/N的增大而减小, 凋落物的初始P、N和木质素含量及木质素/N比值可以作为预测亚热带地区凋落物分解速率的良好指标[8, 32, 33]。本研究发现, 刺槐、油松、柠条和高羊茅凋落叶片的分解速率与初始总C含量、木质素含量和C/N均呈显著负相关关系(P< 0.01), 除油松凋落物外, 刺槐、柠条和高羊茅凋落叶片分解速率与初始N、P含量呈正相关关系(P< 0.01)。这可能是由于松科植物叶表皮强烈木质化, 比叶面积小, 叶片硬度高, 土壤动物和微生物较难快速分解和腐熟[34, 35]; 其他几种凋落叶片比叶面积较大, 即单位质量凋落物具有更大的环境接触面积, 一方面加大了雨水的淋溶作用, 另一方面更有利于微生物定居和土壤动物的碎裂作用, 从而大大提高了分解速率。因此, 不同凋落叶片功能特征(如比叶面积、叶片厚度、硬度等)可能是影响其分解速率的重要因素之一; 凋落叶片的全C、木质素含量和C/N与其分解速率均呈显著负相关关系, 是控制其分解速率的最主要因素, 因此可以作为预测该地区凋落叶片分解速率的指标; 而初始N、P含量与凋落叶片分解速率的相关性在不同凋落物之间存在差异, N、P含量是否可以作为预测该地区凋落叶片分解速率的指标尚需进一步研究。
通过对黄土丘陵区刺槐、油松、柠条和高羊茅凋落叶一年内分解及养分释放情况研究发现, 4种凋落叶分解均符合Olson 指数模型, 分解速率为柠条凋落叶> 刺槐凋落叶> 高羊茅凋落叶> 油松凋落叶。C是凋落叶片分解质量损失的主体, C、P元素属于直接释放模式, N元素属于富集— 释放模式。分解过程中4种凋落叶片的C/N、P含量持续降低, 木质素含量缓慢增加, 纤维素含量呈现先增加后降低的变化趋势, 但不同凋落叶片分解过程中全C和全N含量的变化趋势存在差异。相关性分析表明, 凋落叶片的分解速率与其初始全C含量、木质素含量和C/N均呈显著负相关关系(P< 0.01), 凋落叶片初始全C、木质素含量和C/N越高, 分解速率越低, 因此在研究区内凋落叶片的全C、木质素含量和C/N是控制凋落叶片分解速率及其养分释放的主要因素, 可以作为预测研究区凋落叶片分解的主要指标。研究结果为我们深入了解黄土丘陵区不同退耕地物质循环提供一定的理论依据。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|