

{kind=link}
{kind=link}
{kind=link}
{kind=link}
虉草适应湿地环境的解剖结构和组织化学特征研究
[王晓娥1  , 张梵
, 张梵1 , 张霞1 , 周存宇1 , 杨朝东1, 2, * ]
, 张梵]
|
|


作者简介:王晓娥(1993-),女,土家族,湖北恩施人,在读硕士。E-mail: 344072499@qq.com
为研究虉草适应湿地环境的结构特征,探讨了该植物不定根、根状茎、直立茎和叶的解剖结构、组织化学及其质外体屏障结构的通透性生理。利用光学显微镜和荧光显微镜对虉草进行了解剖学和组织化学研究。结果表明:1)虉草不定根解剖结构由外而内分别为表皮、外皮层、皮层、内皮层、中央为木质部和厚壁组织;茎结构由外而内分别为角质层、表皮、周缘厚壁机械组织层、皮层、具维管束厚壁机械组织层和髓腔。2)不定根具栓质化和木质化内皮层和外皮层组成的屏障结构;茎具角质层、栓质化和木质化周缘厚壁组织层和厚壁组织层;根状茎具内、外皮层,直立茎仅具外皮层。3)虉草体内气腔包括不定根中通气组织,茎皮层气腔和髓腔,及叶鞘气腔。虉草的机械组织结构、屏障结构和气腔是其适应湿地环境的重要结构特征,该研究为今后选择湿地生态修复的植物物种提供依据。
Reed canarygrass ( Phalaris arundinacea) is a perennial herb adapted to wetland environments. The current work explores the anatomy and histochemistry of the adventitious roots, stems (rhizomes and culms), leaves and the permeability of apoplastic barriers of reed canarygrass. The anatomy and histochemistry of P. arundinacea were studied using an optical microscope and fluorescent microscope. Sections were stained with Sudan red 7B (SR7B) for suberin lamellae, with berberine hemisulfate-aniline blue (BAB) for casparian bands and lignified cell walls, phloroglucinol-HCl (Pg) for lignin and berberine hemisulfate-potassium thiocyanate for apoplastic permeability. Concentrated sulfuric acid digestion (AD) was used to detect the wavy casparian bands in young rhizomes and culms. The adventitious roots had a primary structure comprising epidermis, exodermis, cortex, endodermis and vascular cylinder with sclerenchyma; the stems comprised a cuticle, epidermis, peripheral mechanical ring, exodermis, cortex, endodermis, sclerenchyma ring with vascular bundles and pith cavity. The apoplastic barriers consisted of adventitious roots with a suberized and lignified endodermis and an exodermis, stems possessed a cuticle, suberized and lignified peripheral mechanical ring and a sclerenchyma ring, and rhizomes with endodermis and exodermis, culms only with exodermis. The air space consisted of pith cavities and cortical cavities in the stems, and aerenchyma in roots and leaves. This research revealed that reed canarygrass has the key constitutive, structural and histochemical features in roots and stems that allow it to grow in wetlands, be protected against oxygen leakage from aerenchyma, with-stand seasonal flooding and consequently, widely distributed. The anatomical structures and histochemical features of these perennial, rhizomatous grasses enhance our awareness of the biology of this species in native Asian and North American invaded habitats to select species for the restoration of degraded wetland of the Jianghan Floodplain and Three Gorges Reservoir Region of the Yangtze River.
虉草(Phalaris arundinacea)原产于美国北部, 为根状茎的多年生草本植物[1], 其适应能力强, 固土作用明显, 现广泛分布于世界各地, 同时因其营养丰富并作为一种重要牧草[2, 3]。近几十年来, 对虉草种类和其对环境变化已深入研究[4, 5, 6, 7, 8, 9, 10, 11, 12, 13], 而关于虉草的解剖结构特征与适应湿地环境的关系却未见报道。
湿地植物通常具备以下结构特征适应湿地环境, 第一, 其根茎中有发达的机械组织抵御水力冲刷, 如狗牙根(Cynodon dactylon)、双穗雀稗(Paspalum distichum)、菰(Zizania latifolia)等植物的茎作为营养繁殖体, 其机械组织为具维管束的厚壁机械组织层和周缘厚壁机械组织层[14, 15, 16]。第二是屏障结构保持体内空气不易扩散出去和控制内外环境离子的进出而防止影响生命活动, 屏障结构由初生壁凯氏带, 栓质化和木质化次生壁, 体表角质层组成, 如狗牙根, 双穗雀稗, 藜蒿(Artemisia selengensis)等植物的茎具木质化和栓质化的厚壁机械组织层和周缘厚壁机械组织层及角质层, 或者具内、外皮层, 或者具凯氏带并木质化和栓质化的周皮; 不定根具木质化和栓质化的内、外皮层[14, 15, 16]。第三是通气组织为植物在湿地缺氧环境下输导和储藏空气, 包括茎中髓腔和皮层气腔, 不定根中通气组织[14, 15, 16]。根际微生物活动为根内通气组织提供CO2, 便于洪水威胁解除后的光合作用[17]。同一物种在不同生长环境, 或者不同物种间的形态学、解剖结构和屏障结构的组织化学也存在一定的差异[14, 15, 16, 18, 19, 20]。
虉草5月开花, 生长在夏季, 一般淹没于水中10 m左右, 秋季和冬季发芽, 与江汉洪泛区大部分植物的物候不同[21]。因此, 通过对其不定根、根状茎、直立茎和叶的解剖结构和屏障结构研究, 有利于进一步了解其生物学特征, 为后续三峡库区消落带退化生态系统的修复提供理论和实践基础。
于2017年3月下旬, 自湖北省江汉平原湿地洪泛区采淹没的虉草植株, 用自来水洗净, 取完整不定根; 根状茎长40~60 mm, 具3~6个节; 直立茎长610~750 mm, 具11~13个节, 并用FAA固定液固定待用[22], 叶鞘和叶片采用新鲜样本。用新鲜的不定根、根状茎和直立茎进行质外体渗透试验。实验重复5次。
在立体解剖镜(JNOEC JSZ6, China)下, 用双面刀片分别在距根尖10和30 mm处, 根状茎第1节间和第3~5节间, 直立茎第1节间和第7节间, 以及叶的切片。切片用苏丹红7B(SR7B)染色, 栓质化细胞壁呈红色[23], 用硫氢酸黄连素-苯胺蓝对染(BAB)切片检测细胞壁木质化和凯氏带[24, 25](凯氏带呈现生动黄色, 木质化细胞壁呈现呆滞黄色), 盐酸-间苯三酚-(Pg)检测木质化, 呈樱桃红色[10]。用浓硫酸消化切片细胞壁后留下波浪形凯氏带[26, 27, 28]。苏丹红7B染色切片在莱卡光学显微镜(Leica DME)下观察, 用数码相机(NIKON E5400, Japan)拍照记录图片。硫氢酸黄连素-苯胺蓝对染切片在荧光显微镜(Olympus IX71)蓝色激发光下观察, 用与电脑连接的数码相机(RZ200C-21, China)拍照记录图片, 图片用Photoshop 7.0软件处理, 加标尺, 调整图片亮度和对比度。
不定根、根状茎和直立茎的质外体通透性试验步骤如下, 取不定根20~30 mm长的片段, 用吸水纸吸干, 并用粘蜡封口; 根状茎和直立茎样品切取两端有节的节间, 两端不封口, 实验5次重复。不定根、根状茎和直立茎切段浸没于0.05%硫酸氢黄连素溶液中1 h, 再浸没于0.05 mol· L-1硫氰酸钾中0.5 h, 还包括两组对照, 一是不染色, 二是切段仅浸没于0.05%硫酸氢黄连素溶液中1 h。在立体解剖镜(JNOEC JSZ6, China)下徒手切片以上处理后的样品, 置于荧光显微镜(Olympus IX71)蓝色激发光下检测试验结果[24, 27, 28]。
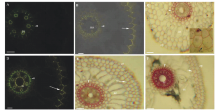
虉草不定根维管柱具6~8个原生木质部(图1A, B, D和F)和1个中央后生木质部(图1B, D和F)及周围具木质化厚壁机械组织(图1D和F)。距根尖10 mm处, 内皮层出现凯氏带(图1A), 距根尖30 mm处, 外皮层2~3层胞壁具“ Y” 凯氏带(图1B), 内皮层和外皮层同时都栓质化, 皮层具通气组织, 表皮未脱落(图1C)。不定根基部切片, 内皮层和外皮层都栓质化和木质化, 内皮层“ U” 形木质化增厚(图1D, E和F), 皮层具通气组织, 表皮仍存在。
 | 图1 虉草不定根横切的显微照片 A:内皮层凯氏带(箭头), 原生木质部, BAB, 标尺=50 μ m; B:内皮层(箭头)和外皮层(箭头)的凯氏带, 后生木质部, BAB, 标尺=50 μ m; C:完全栓质化内皮层(箭头)和轻微栓质化外皮层(箭头), 通气组织, SR7B, 标尺=50 μ m; D:内皮层(箭头)和外皮层(箭头)凯氏带, 插入图-凯氏带之间的切向壁凯氏带, 中柱厚壁机械组织层, BAB, 标尺=50 μ m; E:内皮层(箭头)和外皮层(箭头)栓质化, 通气组织, SR7B, 标尺=50 μ m; F:内皮层具有“ U” 形次生壁(箭头)和外皮层(箭头)轻微木质化, 中柱厚壁机械组织层, Pg, 标尺=50 μ m; 图中字母缩写含义:ae, 通气组织; ep, 表皮; mx, 后生木质部; px, 原生木质部; sc, 中柱厚壁机械组织; BAB, 黄连素-苯胺蓝对染; SR7B, 苏丹红7B染色; Pg, 盐酸-间苯三酚染色。Fig.1 Photomicrographs of P. arundinacea adventitious root transverse sections A: Endodermis casparian bands (arrowhead), protoxylem poles, BAB, scale bars=50 μ m; B: Casparian bands of endodermis (arrowheads) and exodermis (arrows), metaxylem, BAB, scale bars=50 μ m; C: Completely suberized endodermis (arrowhead) and lightly suberized exodermis (arrows), aerenchyma, SR7B, scale bars=50 μ m; D: Endodermis (arrowheads) and exodermis (arrows) casparian bands, inset, tangential casparian walls between casparian bands, stele sclerenchyma, BAB, scale bars=50 μ m; E: Endodermis (arrowheads) and exodermis (arrows) suberin lamellae, aerenchyma, SR7B, scale bars=50 μ m. F: Endodermis with U-shaped secondary walls (arrowheads) and exodermis (arrows) lightly lignified, stele sclerenchyma, Pg, scale bars=50 μ m. Abbreviations in figures: ae, aerenchyma; ep, epidermis; mx, metaxylem; px, protoxylem; sc, stele sclerenchyma; BAB, berberine aniline blue stained; SR7B, sudan red 7B stained; Pg, hydrochloric acid-phloroglucinol stained. |
根状茎尖具维管束厚壁组织层及轻微栓质化和木质化, 具气腔的皮层, 具凯氏带的表皮及外侧角质层(图2A, B), 中央为髓腔。根状茎第三节间具内皮层, 厚壁组织层和周缘厚壁层明显木质化(图2C)。根状茎成熟节间, 内皮层具凯氏带并明显栓质化和木质化, 厚壁组织层和周缘厚壁层强烈栓质化和木质化, 皮层具细小栓质化和木质化维管束, 皮层气腔和髓腔扩大(图2D, E和F)。
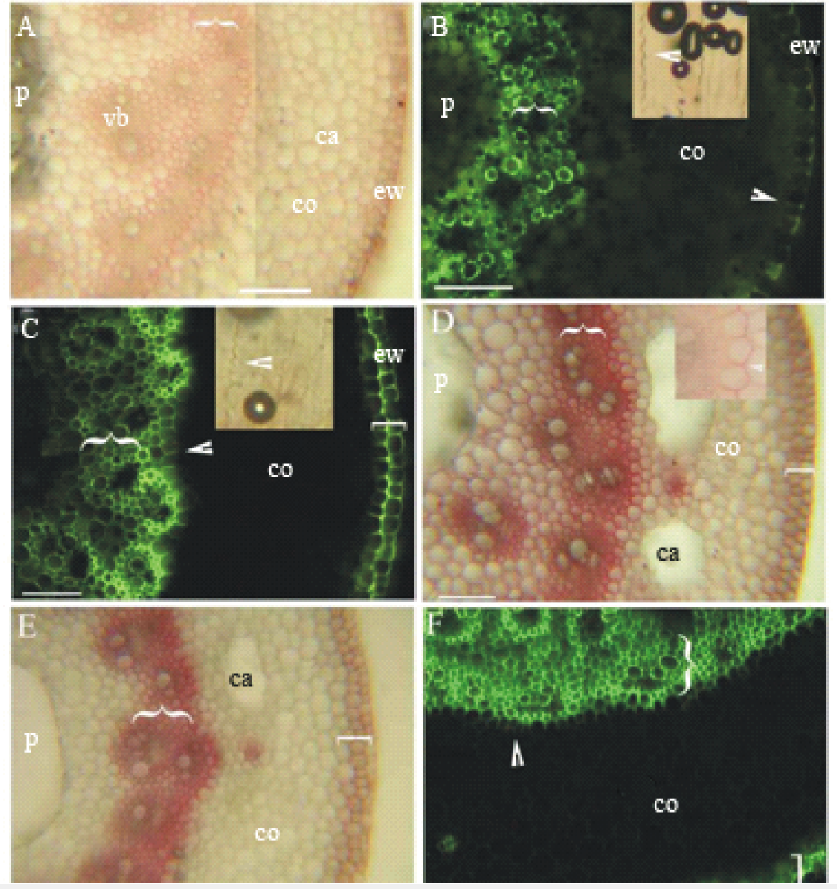 | 图2 虉草根状茎(长40~60 mm, 具3~6节)在第1节间和第3~5节间横切的显微照片 A:第一节间横切, 表皮外切向壁的角质层, 具维管束的厚壁机械组织层({), 髓, 皮层气腔, 维管束, 皮层, SR7B, 标尺=100 μ m; B:第一节间横切, 表皮外切向壁的角质层, 厚壁机械组织层({), 外皮层凯氏带(箭头), 髓, 皮层; 插入图-浓硫酸消化后内皮层凯氏带(箭头), BAB, 标尺=100 μ m; C:第三节间横切, 表皮外切向壁的角质层, 厚壁机械组织层({), 周缘厚壁机械组织层([), 内皮层凯氏带(箭头), 皮层, 插入图-浓硫酸消化内皮层凯氏带, BAB, 标尺=100 μ m; D:第五节间横切, 厚壁机械组织层栓质化({), 周缘厚壁机械组织层栓质化([), 髓腔, 皮层, 皮层气腔, 插入图-内皮层栓质化, SR7B, 标尺=100 μ m; E:第五节间横切, 厚壁机械组织层强烈木质化({), 周缘厚壁机械组织层([), 皮层具细小维管束, 皮层气腔, 髓腔, Pg, 标尺=100 μ m; F:第五节间横切, 厚壁机械组织层({), 周缘厚壁机械组织层([), 内皮层凯氏带(箭头), 皮层, BAB, 标尺=100 μ m; 图中字母缩写含义:ca, 气腔; co, 皮层; ew, 表皮胞壁外角质层; vb, 维管束; p, 髓腔; PMR, 周缘厚壁机械组织层; SCR, 厚壁机械组织层; BAB, 黄连素-苯胺蓝对染; SR7B, 苏丹红7B染色; Pg, 盐酸-间苯三酚染色。Fig.2 Photomicrographs of P. arundinacea rhizomes (40-60 mm long, 3-6 nodes) sectioned at the 1st internode and at 3-5th internode A: 1st internode, cuticle on epidermis outer tangential walls, SCR (brace) with vascular bundles, pith, cortical cavities, vascular bundle, cortex, SR7B, scale bars=100 μ m; B: 1st internode, cuticle on epidermis outer tangential walls, SCR (brace), casparian bands on exodermis (arrowheads), pith, cortex, inset-AD of exodermis casparian bands (arrowhead), BAB, scale bars=100 μ m; C: 3rd internode, cuticle on epidermis outer tangential walls, SCR (brace), PMR (bracket), casparian bands on endodermis (arrowheads), cortex, upper inset-AD of endodermis casparian bands, BAB, scale bars=100 μ m; D: 5th internode, suberin in wide SCR (brace), suberin in PMR (bracket), pith with cavity, cortex, cortical cavities, inset, suberin lamellae of endodermis, SR7B, scale bars=100 μ m; E: 5th internode, heavily lignified SCR (brace), PMR (bracket), small cortical vascular bundles, cortical cavities, pith with cavity, Pg, scale bars=100 μ m; F: 5th internode, widely SCR (brace), PMR (bracket), casparian bands on endodermis (arrowheads), cortex, BAB, scale bars=100 μ m. Abbreviations in figures: ca, cavity; co, cortex; ew, cuticle on epidermal cell wall; vb, vascular bundle; p, pith cavity; PMR, peripheral mechanical ring; SCR, sclerenchyma ring; BAB, berberine aniline blue stained; SR7B, sudan red 7B stained; Pg, hydrochloric acid-phloroglucinol stained. |
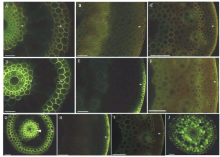
直立茎尖具维管束厚壁组织层, 皮层具叶绿素, 具凯氏带的表皮及外侧角质层(图3A, B), 中央为髓腔。直立茎成熟节间, 厚壁组织层和周缘厚壁层强烈栓质化和木质化, 皮层和厚壁组织层内部薄壁细胞也栓质化和木质化, 皮层气腔和髓腔扩大(图3C, D和E)。叶鞘具气腔, 薄壁细胞具叶绿素, 外表具角质层(图3F), 叶片叶肉具叶绿素, 外表具角质层(图3G)。
 | 图3 虉草直立茎(610~750 mm, 11~13节)在第1节间和第7节间横切的显微照片 A:第一节间横切, 表皮外切向壁上的角质层, 厚壁机械组织层({), 维管束, 皮层, 具叶绿素组织层, 皮层, 髓腔, SR7B, 标尺=100 μ m; B:第一节间横切, 外皮层凯氏带(箭头), 表皮外切向壁的角质层, BAB, 插入图-浓硫酸消化外皮层凯氏带(箭头), 标尺=100 μ m; C:第七节间横切, 厚壁机械组织层强烈木质化({), 周缘厚壁机械组织层([), 厚壁机械组织层内部薄壁细胞木质化(* ), Pg, 标尺=100 μ m; D: 第七节间横切, 厚壁机械组织层木质化({), 周缘厚壁机械组织层([), 厚壁机械组织层内部薄壁细胞木质化(* ), 皮层, BAB, 标尺=100 μ m; E:第七节间横切, 周缘厚壁机械组织层到髓腔的胞壁栓质化(双向箭头), 维管束, 皮层气腔, 皮层, SR7B, 标尺=100 μ m; F:叶鞘表皮外切面壁的角质层(箭头), 皮层或叶肉气腔, 具叶绿素组织层, SR7B, 标尺=100 μ m; G:叶片表皮外切面壁的角质层(箭头), 具叶绿素叶肉组织层, SR7B, 标尺=100 μ m; 图中字母缩写含义:ca, 气腔; co, 皮层; ew, 表皮胞壁外角质层; vb, 维管束; p, 髓腔; chl, 具叶绿素组织层; PMR, 周缘厚壁机械组织层; SCR, 厚壁机械组织层; BAB, 黄连素-苯胺蓝对染; SR7B, 苏丹红7B染色; Pg, 盐酸-间苯三酚染色。Fig.3 Photomicrographs of P. arundinacea culms (610-750 mm, 11-13 nodes) sectioned at the 1st internode and at 7th internode A: 1st internode, cuticle on epidermis outer tangential walls, SCR (brace), vascular bundle, cortex, chlorenchyma, cortex, pith with cavity, SR7B, scale bars=100 μ m; B: 1st internode, casparian bands on exodermis (arrow), cuticle on epidermis outer tangential walls, BAB, inset-AD of exodermis casparian bands (arrowhead), scale bars=100 μ m; C: 7th internode, heavily lignified SCR (brace), PMR (bracket), lignified parenchyma cells inside the SCR (* ), Pg, scale bars=100 μ m; D: 7th internode, lignified SCR (brace), PMR (bracket), lignified parenchyma cells inside the SCR (* ), cortex, BAB, scale bars=100 μ m; E: 7th internode, suberized cells from PMR extend to pith cavity (two arrows line), vascular bundles, cortical lacuna, cortex, SR7B, scale bars=100 μ m; F: Cuticle on the epidermis outer tangential walls of leaf sheath (arrows), cortical or mesophyll cavity, chlorenchyma, SR7B, scale bars=100 μ m; G: Cuticle on the epidermis outer tangential walls of leaf blade (arrows), chlorenchyma, SR7B, scale bars=100 μ m. Abbreviations in figures: ca, cavity; co, cortex, ew, cuticle on epidermal cell wall; vb, vascular bundle; p, pith cavity; chl, chlorenchyma; PMR, peripheral mechanical ring; SCR, sclerenchyma ring; BAB, berberine aniline blue stained; SR7B, sudan red 7B stained; Pg, hydrochloric acid-phloroglucinol stained. |
不定根维管柱、内皮层和外皮层有较强自发黄色荧光, 而根状茎和直立茎自发荧光较弱(图4A, B和C)。仅黄连素处理切段, 不定根维管柱、内皮层和外皮层, 根状茎表皮强烈吸收黄连素而具强烈黄色荧光, 直立茎仍为自发黄色荧光(图4D, E和F)。黄连素和硫氰酸钾处理切段, 不定根维管柱、内皮层和外皮层, 根状茎表皮强烈吸收黄连素而具强烈黄色荧光, 直立茎仍为自发黄色荧光(图4G, H和I), 不定根外皮层破裂, 内部具硫氰酸黄连素晶体而发强烈黄色荧光(图4J)。该实验表明不定根完整外皮层能吸收阻挡离子进入皮层及内部, 根状茎表皮吸收阻挡离子进入, 直立茎角质层和表皮完全阻挡离子进入。
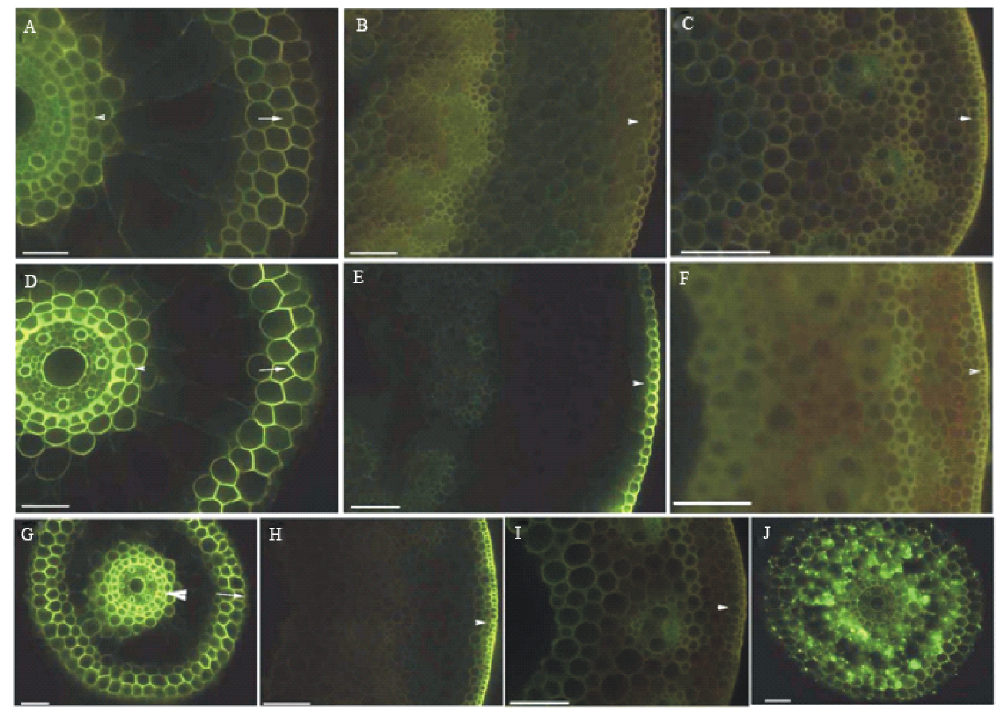 | 图4 黄连素示踪不定根﹑根状茎和直立茎质外体屏障结构的通透性测试 从150 mm长的根选择根尖40~70 mm长, 老根状茎和直立茎切成含两节的切段, 两端不封闭, 除非另有说明。 A:具通气组织的不定根, 内皮层(箭头)和外皮层(箭头)淡黄色, 未染色, 标尺=50 μ m; B:根状茎表皮呈淡黄色(箭头), 未染色, 标尺=100 μ m; C:直立茎表皮呈淡黄色(箭头), 未染色, 标尺=100 μ m; D:具通气组织的不定根, 内皮层(箭头)和外皮层(箭头)发黄色荧光, 具通气组织的不定根, 染色, 标尺=50 μ m; E:根状茎表皮发黄色荧光(箭头), 黄连素染色, 标尺=100 μ m; F:直立茎表皮发出淡黄色荧光(箭头), 黄连素染色, 标尺=100 μ m; G:具通气组织的不定根, 内皮层(箭头)和外皮层细胞壁(箭头)呈亮黄色荧光, 黄连素和硫氰酸钾(KSCN)染色, 标尺=50 μ m; H:根状茎表皮呈亮黄色荧光(箭头), 黄连素和硫氰酸钾(KSCN)染色, 标尺=100 μ m; I:直立茎表皮呈亮黄色荧光(箭头), 黄连素和KSCN染色, 标尺=100 μ m; J:不定根外皮层破裂, 通气组织具硫氰酸黄连素晶体, 黄连素和硫氰酸钾染色, 标尺=50 μ m。Fig.4 Apoplastic barriers permeability tests on roots, rhizomes and culms with berberine tracer Roots segments sectioned at 40 mm to 70 mm from the tip of 150 mm long roots, the aged rhizome and culm samples excised with one internode and two nodes, and unsealed ends, except where noted. A: Roots with aerenchyma, endodermis (arrowhead) and exodermis (arrow) light yellow, unstained, scale bars=50 μ m ; B: Epidermis of rhizomes light yellow (arrowhead), unstained, scale bars=100 μ m; C: The epidermis of culms fluoresced light yellow (arrowhead), unstained, scale bars=100 μ m; D: Roots with aerenchyma, endodermis (arrowhead) and exodermis cell walls (arrow) fluoresced yellow, berberine stained, scale bars=50 μ m; E: The epidermis of rhizomes fluoresced yellow (arrowhead), berberine stained, scale bars=100 μ m; F: The epidermis of culms fluoresced light yellow (arrowhead), berberine stained, scale bars=100 μ m; G: Roots with aerenchyma, endodermis (arrowhead) and exodermis cell walls (arrow) fluoresced bright yellow, berberine and KSCN stained, scale bars=50 μ m; H: The epidermis of rhizomes fluoresced bright yellow (arrowhead), berberine and KSCN stained, scale bars=100 μ m; I: The epidermis of culms fluoresced light yellow (arrowhead), berberine and KSCN stained, scale bars=100 μ m; J: Roots with ruptured exodermis, crystals of berberine thiocyanate enter into aerenchyma, berberine and KSCN stained, scale bar=50 μ m. |
近些年, 主要研究了江汉平原既耐水湿又耐旱生植物的解剖结构和组织化学特征, 如狗牙根, 双穗雀稗, 牛鞭草(Hemarthria altissima), 南方艾蒿(Artemisia lavandulaefolia), 藜蒿, 天胡荽(Hydrocotyle sibthorpioides), 水芹(Oenanthe javanica), 菰, 假俭草(Eremochloa ophiuroides), 白茅(Imperata cylindrica), 其中水芹不长期淹没, 菰不耐长期旱生, 假俭草和白茅以旱生为主, 耐短时间水湿环境[14, 15, 16, 18, 19, 29, 30, 31]。国外主要报道了水稻(Oryza sativa)、芦苇(Phragmites australis), 水甜茅(Glyceria maxima), 鸢尾(Iris), 亚马逊热带雨林耐淹没植物不定根, 以及香蒲(Typha)不定根和根茎和水生毛茛(Ranunculus trichophyllus)叶柄的解剖结构和组织化学[20, 24, 27, 32, 33, 34, 35, 36]。
虉草不定根解剖结构由外而内分别为:表皮、外皮层、皮层、内皮层、中央为木质部和厚壁机械组织层。其屏障结构由具凯氏带并栓质化和木质化的内皮层和外皮层组成, 与已报道湿地植物狗牙根、双穗雀稗、白茅、水稻、芦苇等相同[14, 15, 16, 18, 19, 20, 24, 27, 33, 35], 虉草不定根与菰、狗牙根、双穗雀稗和白茅等都有内皮层邻近1~2层栓质化和木质化的皮层细胞[14, 15, 30], 与香蒲外皮层中的“ Y” 形凯氏带结构类似[ 24], 根中厚壁机械组织层与菰类似[15], 这说明虉草与其他适应湿地和干旱环境的植物具相似屏障结构。
虉草茎(根状茎和直立茎)的结构由外而内分别为:角质层、表皮、周缘厚壁机械组织层、皮层、具维管束厚壁机械组织层和髓腔。其屏障结构为角质层、栓质化和木质化周缘厚壁组织层和厚壁组织层。根状茎具内、外皮层, 直立茎仅具外皮层, 而南方艾蒿和藜蒿茎具内、外皮层, 天胡荽和香蒲仅具内皮层[16, 29, 32]。与狗牙根, 双穗雀稗等相比较, 都具角质层、栓质化和木质化周缘厚壁组织层和厚壁组织层[14, 15]。不同点在于, 南方艾蒿和藜蒿茎外侧具栓质化和木质化周皮[16]。天胡荽和水芹茎表皮下具厚角组织机械组织[28, 31], 双穗雀稗茎皮层和髓部也具厚角组织[14, 18]。假俭草和白茅仅具木质化周缘厚壁组织层[14, 30]。此外, 水稻胚轴具凯氏带, 水生毛茛叶柄维管束鞘细胞具凯氏带[34, 36]。虉草叶鞘和叶片表面具角质层[3, 4, 5, 7, 8, 9, 12, 15, 21, 23]。
虉草体内气腔包括不定根中通气组织, 茎皮层气腔和髓腔, 及叶鞘气腔。与狗牙根, 假俭草, 牛鞭草, 水芹, 菰, 香蒲等体内气腔类似[14, 15, 16, 18, 19, 20, 24, 31, 32, 33, 35, 37]。而双穗雀稗茎皮层和髓部具蜂窝状通气组织[14, 18], 天胡荽皮层和髓部水湿胁迫后具通气组织[28], 白茅无髓腔[30]。虉草根中气腔为裂溶生通气组织, 茎皮层气腔和髓腔为溶原生性腔, 这些通气组织有助于植物在缺氧环境中把氧气储存和运输到其他器官[38], 是使其能够适应湿地环境重要的结构特征之一。
根据虉草的屏障结构和解剖结构特点表明, 不定根具栓质化和木质化的内皮层和外皮层, 茎具角质层、内皮层、外皮层、栓质化和木质化周缘厚壁组织层和厚壁组织层, 根和茎叶中具通气组织是其适应湿地环境的重要结构特征, 为今后选择合适的物种恢复江汉平原和长江三峡库区消落带生态环境具有重要意义[21]。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|