





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
放牧对晋北农牧交错带赖草草地生态系统碳交换的影响
[邢鹏飞1  , 李刚
, 李刚1 , 陈晓鹏1 , 李德颖2 , 王常慧3 , 董宽虎1 , 赵祥1, * ]
, 李刚]
|
|


作者简介:邢鹏飞(1994-),男,山西高平人,在读硕士。E-mail:sxpfxing@163.com
生态系统净碳交换(NEE)是评估碳循环及收支的重要指标,由生态系统总初级生产力(GEP)及生态系统呼吸(ER)共同决定。以晋北农牧交错带赖草草地为研究对象,利用LI-840a静态箱法,通过观测为期2年(2015-2016)围封和放牧管理下NEE、ER和GEP的日动态和季动态,分析其与气温、土壤温度(0~10 cm)、土壤含水量(0~10 cm),以及地上和地下生物量的相关性,旨在说明放牧对农牧交错带赖草草地生态系统碳交换的影响。结果表明:放牧显著降低了赖草草地地上、地下生物量;NEE、ER和GEP的日变化及季节变化特征明显,均呈现单峰型变化,在5、6、9月较低,在7-8月较高;生态系统碳交换的日变化主要受气温影响,而季节变化主要受表层土壤温度影响;同时NEE、ER和GEP与地上、地下生物量也存在极显著相关性;相比围封,放牧明显降低了该草地生物量和生态系统碳交换速率,但未改变日、季变化趋势。说明放牧通过破坏草地植被光合组织,降低了草地生态系统碳交换能力,减弱了草地生态系统碳汇功能。
, LI Gang
Ecosystem net carbon exchange (NEE) is an important index to evaluate the carbon cycle, and is calculated from total primary productivity (GEP) and ecosystem respiration (ER). The aim of this study was to explore the effects of grazing on carbon exchange in the Leymus secalinus grassland ecosystem in the agro-pastoral ecotone of Northern Shanxi. Using the static chamber technique (LI-840a), we measured the daily and seasonal dynamics of NEE, ER, and GEP in the L. secalinus grassland under grazing during 2015 and 2016, and analyzed the correlation between these indexes and atmospheric temperature, soil temperature, soil moisture content, and aboveground and belowground biomass. The NEE, ER, and GEP showed clear daily and seasonal variations, which were the unimodal type. The values of NEE were lower in May, June, and September, but higher in July and August. The daily dynamics of NEE were mainly controlled by air temperature, while the seasonal changes were mainly controlled by soil temperature. Correlations between aboveground biomass as well as belowground biomass and NEE, ER, and GEP were extremely significant. Compared with fencing, grazing significantly decreased the NEE rate of the ecosystem in this grassland, but it did not affect the daily and seasonal trends in NEE. These results showed that, compared with enclosure, grazing can decrease the carbon exchange capacity and weaken the carbon sink function of a grassland ecosystem by reducing the proportion of photosynthetic matter in grassland vegetation.
生态系统净碳交换(net ecosystem carbon exchange, NEE), 是由光合作用碳吸收量与呼吸作用碳释放量之间的不平衡所决定的[1], 是生态系统总初级生产力(gross ecosystem primary productivity, GEP)与生态系统呼吸(ecosystem respiration, ER)两个过程的平衡[2], 它体现生态系统的碳汇或碳源能力[3]。由于植物光合作用与生态系统呼吸作用受到植被、土壤和环境因子等多种因素的调控, 而生态系统的气候、养分条件又处于不断变化的状态, 所以不同生态系统的碳源/汇状态也处于不断变化中[4]。同时, 人类活动也会影响生态系统净碳交换[5]。研究表明, 草地的固碳能力对不同草地类型、不同放牧利用方式的响应不同[6]。
草地作为陆地生态系统重要组成部分之一[7], 在陆地碳循环中占有重要地位。放牧作为草地的主要利用方式, 家畜的采食行为和践踏作用影响着草地环境, 改变草地植被结构和功能特征。首先, 家畜采食植被, 减少了地上净初级生产力。Semmartin等[8]的研究表明草地地上净初级生产力与放牧强度之间存在负线性关系, 放牧强度的变化控制了地上净初级生产力的变化。家畜采食植物茎叶, 减少了群落地上生物量, 破坏了植物的光合组织, 因此影响了草地生态系统对碳的吸收。其次, 放牧对植物地下生物量结构的影响很难预测, 已有的研究结果对此意见不一。有研究表明放牧降低了草地地下生物量[9], 而Rueda等[10]在地中海一带的研究表明, 在一些低地中放牧增加了草地表层土的根生物量。此外, 放牧转移植物生物量, 食草动物改变了草地冠层结构及能量平衡, 引起土壤温度和土壤水分平衡的反馈, 因而影响土壤微生物活性, 最终影响净碳吸收[11]。土壤温度的降低也会减慢植被根系的生长、对矿质元素等养分的吸收及向地上部的转运速率、微生物的活动, 从而减小ER[11]。
赖草(Leymus secalinus)为禾本科赖草属植物, 其根茎繁衍迅速, 耐寒并耐旱, 且具有较强的耐盐性[12]。赖草草地作为山西北部农牧交错带低地盐渍化草地的一种重要草地资源, 近年来由于不合理的放牧致使草地群落结构简单化, 生产功能弱化, 从而影响了草地的碳输入, 因此要准确评估我国北方农牧交错带上该草地生态系统碳收支, 有必要对其生态系统碳交换进行研究。本研究对围封和放牧管理8年的赖草草地碳交换速率的日、季动态进行连续观测2年, 分析其与土壤温度、含水量及生物量的关系, 旨在揭示这两种管理方式下赖草草地净碳交换速率的变化特征。
本实验在山西省朔州市右玉县(E 112° 19.660', N 39° 59.808')山西农业大学右玉草地生态系统野外观测研究站进行。海拔1348 m, 年均气温4.7 ℃, 最冷月(1月)均温-14 ℃(-17.8~-9.9 ℃), 最热月(7月)均温20.5 ℃(18.9~22.4 ℃), ≥ 0 ℃年积温2600~3600 ℃。终霜期在5月初, 初霜期在9月中、上旬, 无霜期100~120 d。全年太阳总辐射量为598 kJ· cm-2, 年日照2600~2700 h; 年均降水量435 mm, 属温带大陆性季风气候。样地土壤类型为苏打草甸碱土, pH 9.2。地区植被类型属温性草原类, 试验地草地属低地草甸类盐化低地草甸亚类, 草地型为赖草-鹅绒委陵菜(Potentilla anserina)-杂类草群丛, 主要生长有赖草、鹅绒委陵菜、蒲公英(Taraxacum mongolicum)、艾蒿(Artemisia argyi)、碱地风毛菊(Saussurea runcinata)、碱茅(Puccinellia distans)、碱蓬(Suaeda glauca)、车前(Plantago asiatica)、西伯利亚蓼(Polygonum sibiricum)、芦苇(Phragmites communis)等。
试验地于2008年开始围栏封育, 放牧地为全年自由放牧, 放牧强度为每公顷2.00个羊单位, 放牧和围栏样地面积150 m× 150 m。在样地对角线上等距离设10个1 m× 1 m样方。
2015和2016年5-9月每月的4和24日测定生态系统净碳交换速率(NEE)、生态系统呼吸(ER)、大气温度、0~10 cm土壤温度、0~10 cm土壤含水量、植被地上生物量、0~100 cm地下生物量, 其中植被地上、地下生物量于每月中旬进行测量。
1.3.1 NEE和ER的测定 采用静态箱法, 测定前1周在放牧和围封样地各安装10个底座(即测定重复)。采用样线法每隔10~15 m安装一个底座。底座高8 cm, 插入地下5 cm, 地上部露出3 cm。箱体由透明有机玻璃构成, 长宽高均为50 cm, 箱体内配有2个风扇, 使箱内气体充分混匀, 测量时通过导气管与LI-840a CO2/H2O红外分析仪连接, 将同化箱盖在底座上测定箱内CO2和H2O浓度的变化, 测量时间120 s, 记录开始和结束时箱体内的温度。完成NEE测定后通风、复扣同化箱, 盖上遮阳罩测定ER, 最后计算GEP。在测定生态系统碳交换速率的同时测定0~10 cm土壤温度、0~10 cm土壤湿度、大气温度等, 大气和土壤温度用空气温度计和土壤温度计直接测得, 土壤湿度采用TDR 300土壤水分速测仪(Specctrum, USA)测得。每月上旬和下旬各测定一次, 每次监测时间为24 h, 当日7:00开始, 次日7:00结束, 每3 h测定一次。
采用线性拟合的方法计算CO2浓度变化速率(dc/dt), 即斜率。CO2交换速率的计算采用以下公式:
Fc=
其中, Fc为CO2通量(μ mol· m-2· s-1); V为箱体的体积, 即V=同化箱长× 宽× (箱体高度+底座露出地上部的高度); Pav为测量期间箱体内的平均大气压强(kPa); Wav是测量期间箱体内的水气分压(mmol· mol-1); R是大气常数8.314 J· mol-1· K-1; S是同化箱的面积; Tav是测量期间箱体内的平均温度(℃)。
GEP=ER-NEE
当NEE为负值时, 表示吸收, 即为碳汇; NEE为正值时, 表示释放, 即为碳源。
日平均土壤温度是用各个时间段的土壤温度累加平均求得, 日平均土壤含水量是用各个时间段的土壤含水量累加平均求得。
1.3.2 草地地上、地下生物量的测定 草地群落地上和地下生物量采用收获法测定。在生长季每月中旬于样方内齐地面剪下全部植物称鲜重。把所有刈割的样品放入信封, 于烘箱中恒温65 ℃烘干48 h至恒重后称重。采用根钻法(直径70 mm)测定地下生物量, 在地上部分刈割完的样方内按对角线法取根样各3钻, 分7层(0~5 cm、5~10 cm、10~20 cm、20~30 cm、30~50 cm、50~70 cm、70~100 cm)取样, 取得的地下生物量放入网袋带回实验室洗根, 剔除死根, 然后65 ℃烘干至恒重称重。
采用Microsoft Excel 2010进行基础数据的输入与整理, 用Sigma Plot 12.5进行回归分析和绘图, 同时采用SPSS 19.0统计分析软件进行数据独立样本t检验和单因素方差分析, 用Pearson进行双因素相关分析。
生长季内围封和放牧的赖草草地群落地上、地下生物量均呈单峰型动态变化(图1), 除围封下地下生物量最大值出现在9月, 其余最大值均出现在8月。植物在4月底5月初期进入返青期, 但气温仍然较低, 植物生长缓慢。之后温度升高, 降水增加, 植物生长加快, 8月生物量达到最大值, 之后生物量呈下降趋势。围封草地每月地上生物量均显著高于放牧草地(P< 0.05), 放牧作用下, 草地地上生物量最高月份比围封草地减少了62.32%, 分别为389.33和146.69 g· m-2。围封样地地下生物量除5月外均显著大于放牧样地(P< 0.05), 放牧下的地下生物量最高月份比围封草地减少了43.29%, 分别为4052.97和2298.45 g· m-2, 放牧作用下, 赖草草地年均地上和地下生物量分别减少了64.55%和44.54%。
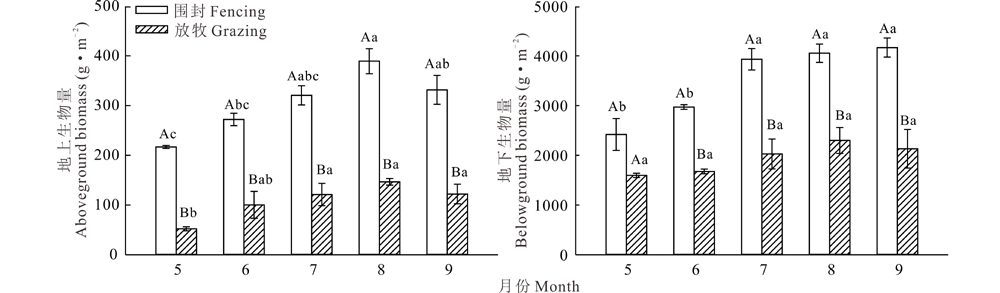 | 图1 封围和放牧下赖草草地生物量的动态 图中不同小写字母表示生长季内不同月份间差异显著(P< 0.05), 不同大写字母表示放牧与围封样地间差异显著(P< 0.05)。Fig.1 Growth dynamics of biomass in L. secalinus communities under fencing and grazing The different small letters mean different months have significant differences; the different capital letters mean the difference between grazing and fencing are significant (P< 0.05). |
2015及2016年围封和放牧样地大气和土壤温度变化趋势一致, 均在7月达到最大值(图2), 而0~10 cm土壤含水量在8月达到最大值。通过比较发现, 与围封相比, 放牧后的土壤温度有上升的趋势, 土壤含水量有下降趋势。两者最高的月份, 放牧比围封土壤温度提高了1.83 ℃, 土壤含水量下降了3.74%, 可能是由于家畜采食导致植被盖度降低, 太阳直射土壤表面导致土壤水分蒸发量加大。整个生长季放牧草地上平均土壤温度提高了1.23 ℃, 平均土壤含水量降低了2.39%。
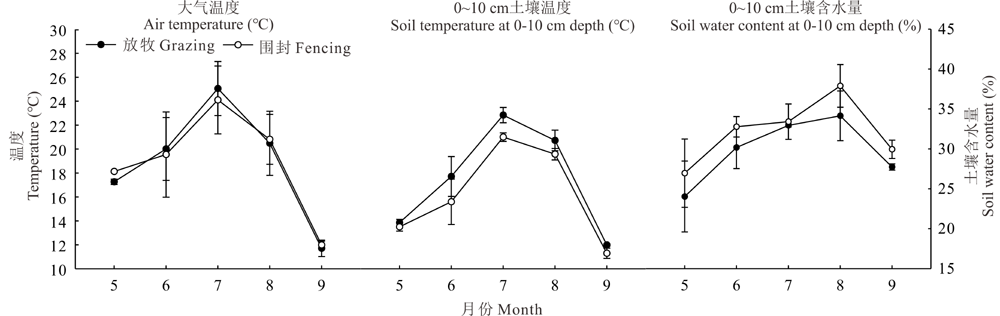 | 图2 不同样地大气温度、土壤温度、土壤含水量的季节动态Fig.2 Seasonal dynamics of air temperature, soil temperature, soil water content in different plots |
2.3.1 赖草草地生长季NEE的日动态 赖草草地NEE具有明显的日变化特征, 表现出白天为负值, 夜晚为正值(图3A)。日出后, NEE由正值转换为负值, 随温度的升高, NEE逐渐增大, 在13:00左右CO2吸收达到最大值, 之后CO2吸收速率逐渐降低, 到傍晚19:00, NEE由负值转化为正值, 昼夜交替的日变化非常明显。生长季放牧草地CO2吸收的最强值出现在10:00-13:00, 为5.22 μ mol CO2· m-2· s-1, 围封草地CO2的吸收最强值也出现在10:00-13:00, 为6.95 μ mol CO2· m-2· s-1, 放牧及围封样地夜间CO2释放的最强值均出现在19:00-22:00, 分别为4.04 和4.58 μ mol CO2· m-2· s-1。NEE日均值均为负值(图3B), 表现为碳吸收, 且放牧地大于围封地, 分别为-0.75和-1.27 μ mol CO2· m-2· s-1, 但未达到显著水平(P> 0.05)。
 | 图3 放牧和围封草地生态系统净碳交换的日动态(A)和日均值(B)Fig.3 Daily dynamics(A) and daily average value(B) of NEE of grazing grassland and fencing grassland |
2.3.2 赖草草地生长季ER的日动态 赖草草地ER的日动态呈单峰型变化趋势(图4A), 白天随气温的升高ER上升, 夜间随气温下降ER逐渐降低, 不同时期由于温度的不同生态系统呼吸速率(ER)的日变化有不同的特点。放牧和围封样地ER的最大值均出现在13:00-16:00, 分别为6.74和7.56 μ mol CO2· m-2· s-1。放牧草地和围封草地的最小值出现在凌晨1:00-4:00, 分别为3.10和4.46 μ mol CO2· m-2· s-1。图4B显示ER的日均值表现为围封地高于放牧地, 分别为5.84和4.67 mmol CO2· m-2· s-1, 高出了25.05%, 未达到显著水平(P> 0.05)。
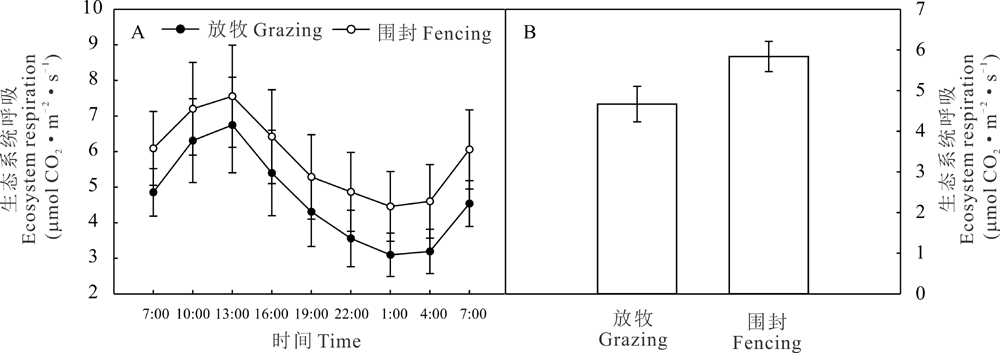 | 图4 放牧和围封草地生态系统呼吸的日动态(A)和日均值(B)Fig.4 Daily dynamics (A) and daily average value (B) of ER of grazing grassland and fencing grassland |
2.3.3 赖草草地生长季GEP的日动态 GEP的日变化趋势也呈单峰型变化(图5A)。上午随温度升高, 光合作用逐渐加强。放牧草地和围封草地GEP最高值都在午后13:00-16:00出现, 分别为11.72 和14.23 μ mol CO2· m-2· s-1。夜晚植物无法进行光合作用, 不吸收碳, NEE与ER相同, 由于GEP=ER-NEE, 故GEP为0。图5B中显示日均值围封地高于放牧地, 分别为5.42和7.10 μ mol CO2· m-2· s-1, 但差异不显著(P> 0.05)。
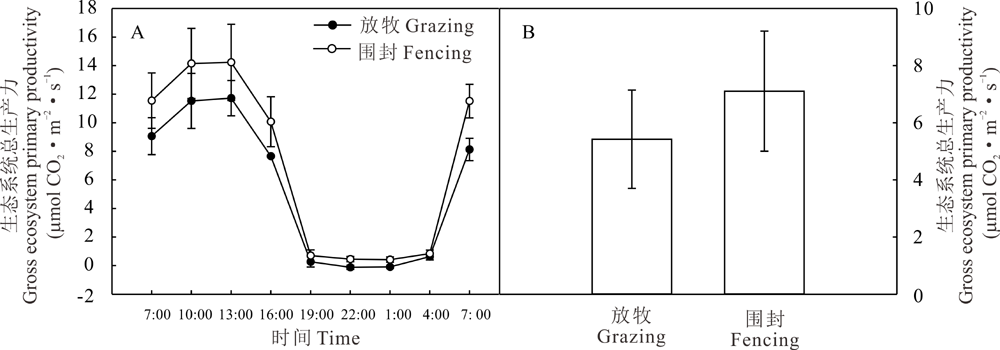 | 图5 放牧和围封草地生态系统总生产力的日动态(A)和日均值(B)Fig.5 Daily dynamics(A) and daily average value(B) of GEP of grazing grassland and fencing grassland |
放牧地和围封地NEE、ER、GEP的季节动态趋势一致, 均在8月达到峰值。如图6A所示, 放牧草地和围封草地的NEE季节变化规律一致, 表现为先降低再升高的趋势, 从5月开始, 随着水热条件逐渐转好, 植物生长加快, 光合作用加强, 8月NEE达到最大, 此时放牧和围封草地的NEE值分别为-1.13和-1.70 μ mol CO2· m-2· s-1。赖草草地在两种处理下的NEE均为负值, 表现为碳汇, 可能是测定生长季植物生长良好, 同时施加的放牧处理强度适中, 使得植物群落在生长季都处于碳汇状态。且NEE均值为放牧草地大于围封草地, 在放牧作用下净碳吸收速率下降了40.94%, 但差异不显著(P> 0.05)。
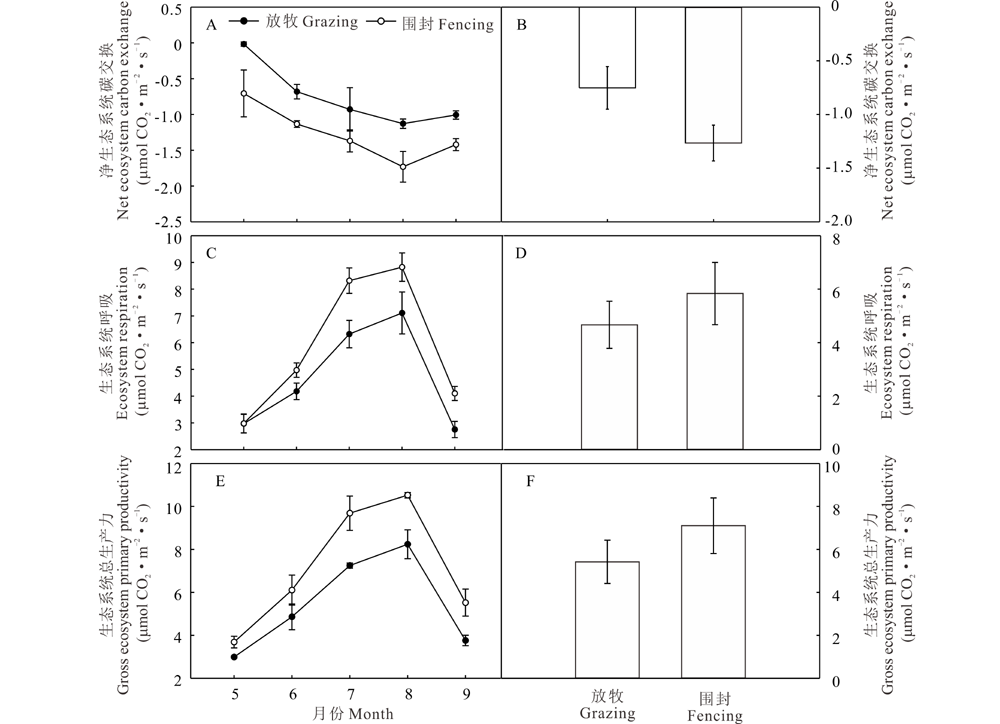 | 图6 放牧和围封草地生态系统NEE、ER、GEP的季节动态 图A、B为NEE的季节动态和均值图, C、D为ER的季节动态和均值图, E、F为GEP的季节动态和均值图。Fig.6 Seasonal dynamics of NEE、ER、GEP of grazing grassland and fencing grassland A and B show the seasonal dynamics and average value of NEE, respectively; C and D show the seasonal dynamics and average value of ER, respectively; E and F show the seasonal dynamics and average value of GEP, respectively. |
从图6C可以看出, 在植物生长季ER具有明显的季节特征, 围栏内外表现出很好的一致性, 即单峰曲线。从5月开始, 赖草草地ER迅速上升, 到8月出现峰值, 此时围封草地的生态系统呼吸速率显著高于放牧草地(P< 0.05), 分别为8.82和7.11 μ mol CO2· m-2· s-1, 9月随着气温降低, 降水减少, 土壤酶活性降低, ER迅速下降。整个生长季ER均值放牧草地低于围封草地(图6D), 放牧作用使ER降低了21.35%, 差异不显著(P> 0.05)。
图6E显示, GEP的季节动态特征也很明显, 且变化趋势与ER一致, 各月GEP值均是放牧草地低于围封草地, 尤其7、8月, 生长旺季差异显著(P< 0.05), 但两种处理下的草地GEP均值差异不显著(P> 0.05), 放牧使得生态系统总生产力降低了23.66%。
围栏内外NEE、ER、GEP的日动态与气温及土壤温度的相关性分析表明(表1), 放牧草地和围封草地的NEE、ER、GEP与大气温度均存在极显著相关性(P< 0.01)。而与0~10 cm土壤温度间的相关性较为复杂, 整个生长季NEE与土壤温度均无显著相关性(P> 0.05), 除7月围栏内外及8月围封草地的ER与土壤温度有极显著相关性外(P< 0.01), 其他月份均无显著相关性(P> 0.05); 7月围封地GEP与土壤温度有显著相关性(P< 0.05), 8月放牧地的GEP 与土壤温度之间相关性达到极显著水平(P< 0.01), 其余月份两种处理方式下的草地GEP与土壤温度之间均无显著相关性(P> 0.05)。同时由于土壤含水量在一天之内变化很小, 所以NEE、ER、GEP的日变化在生长季的每个月份, 在放牧草地与围封草地上与土壤含水量之间均无显著相关性(P> 0.05), 表明在日尺度上气温是对生态系统碳交换过程主要的影响因子。
| 表1 赖草草地NEE、ER、GEP与大气温度和0~10 cm土壤温度及土壤含水量相关性 Table 1 The correlation coefficients between NEE, ER, GEP and atmospheric temperature, 0-10 cm soil temperature and soil water content in L. secalinus grassland |
回归分析表明(表2), NEE的季节变化与土壤温度和大气温度间不存在显著相关性(P> 0.05), 与土壤含水量仅在放牧地存在显著线性负相关(P< 0.05), 因为NEE均为负值, 而在围封地则无显著相关性(P> 0.05)。ER与温度、土壤含水量的相关性均为指数关系(P< 0.05), 即随着温度和土壤水分含量的升高ER逐渐增大。GEP与3个环境因子之间均存在显著相关关系(P< 0.05), 且以线性负相关为主。而NEE、ER、GEP与生物量之间以线性正相关为主(P< 0.05), NEE与地上和地下生物量之间有二次项关系, 可能是由于优势植物生长茂盛, 阻断了太阳光向地面及下层植物的照射, 同时随着根系增多土壤呼吸增多, 导致CO2净吸收先上升后下降。由表2可以看出, 各自变量与土壤温度的拟合效果要优于土壤含水量, 表明在季节尺度上土壤温度成为影响生态系统碳交换的主要因子。
| 表2 赖草草地NEE、ER和GEP的季节变化与温度、土壤含水量及植物生物量的关系 Table 2 Relationship between seasonal change of NEE, ER, GEP and temperature, soil moisture, plant biomass in L. secalinus grassland |
温度和水分是影响生态系统碳交换的重要环境因子[13, 14, 15, 16]。温度通过影响光合作用酶活性、空气湿度控制气孔导度而影响植物光合作用[17, 18], 而土壤水分则与土壤微生物活性、植物生理活性有关。武倩等[19]的增温实验表明, 增温使得植物冠层温度增加, 光合作用增强, 同时也增强了土壤微生物活性, 增强了土壤呼吸进而使得ER增加; 胡毅等[20]的研究则表明土壤含水量与NEE和GEP呈线性负相关关系, 与ER呈现指数相关关系。本试验中赖草草地NEE、ER和GEP均具有明显的日变化特征, 白天随日出温度升高, NEE、ER和GEP逐渐增大, 由碳释放转为碳吸收, 中午13:00左右达到一天中的最大值, 之后随温度降低开始下降, 由碳吸收转为碳释放。但由于土壤温度在一天中变化较小, 故对NEE、ER和GEP日变化的影响不显著。此外, 其季节变化趋势也较为明显, 呈现“ 钟型” 趋势, 先增大后降低, 最大值均出现在8月。并通过分析NEE、ER和GEP季节变化与温度、土壤含水量的相关性, 发现NEE与环境因子之间相关性不显著, 可能是由于试验过程中7月的气温及土壤温度最高, 而土壤含水量较低, 过高的温度以及一定程度上的水分亏缺使得空气饱和水汽压差增大, 从而导致蒸腾增加, 植被叶片气孔关闭[21], 从而降低生态系统净碳吸收, 而此时较高的温度也可能会提升相关酶活性, 促进土壤呼吸作用[13], 也会影响净碳交换及其对环境因子的响应。ER和GEP与环境因子之间存在极显著相关性, 说明环境因子不直接影响生态系统净碳交换, 而是通过影响生态系统呼吸和光合来间接影响净碳交换, 这与伏玉玲等[22]研究羊草(Leymus chinensis)草原的结果基本一致。ER和GEP与温度的相关性大于与水分的相关性, 说明生态系统碳交换对温度的敏感性大于水分, 这与靳宇曦等[23]关于短花针茅(Stipa breviflora)荒漠草原的研究结果基本一致, 即土壤温度对生态系统净碳交换的贡献大于土壤湿度。
放牧家畜主要通过采食和践踏直接影响植物群落组成、地上生物量(或地上净初级生产力)、凋落物而影响NEE[24], 此外, 还会引起土壤温度和土壤水分平衡的反馈, 最终影响净碳吸收[25]。本研究中围封和放牧下NEE、ER和GEP在日尺度和季节尺度上的变化趋势没有差异, 但是与放牧相比, 围封显著增加了碳交换速率。在季节尺度上, 放牧和围封样地NEE值分别为-0.75和-1.27 μ mol CO2· m-2· s-1, 表明在季节尺度上围封样地的碳吸收速率大于放牧样地, 原因是围封后植被得以恢复[26]。地上绿色植被是植物固定光能的唯一途径, 而放牧家畜通过采食, 降低了植被地上生物量, 破坏了植株的光合器官, 使群落光合作用下降, 因此降低了生长季群落的碳汇功能。同时, 放牧地ER小于围封地。因为地上生物量减少, 导致每年的凋落物及立枯物较少, 从而使得土壤C、N含量减少[27], 且地上植被是根系获得能量的途径, 地上植被的减少必然影响到植被地下根系, 进而影响根系呼吸, 而根呼吸是土壤呼吸的重要组成部分[28]。因此, 围封通过增加群落地上、地下生物量进而增强了群落总呼吸[29]。由于GEP等于碳净交换量与生态系统呼吸释放碳量之和, 故放牧同时也降低了群落GEP。2015年赖草群落围封样地和放牧样地GEP总值分别为3826.49和3002.81 g CO2· m-2, ER释放CO2分别为3082.2和2675.2 g CO2· m-2, 净固碳量分别为744.3和327.6 g CO2· m-2, 2016年围封样地和放牧样地GEP总值分别为4699.15和3501.78 g CO2· m-2, 生态系统呼吸总释放量分别为3923.64和2925.85 g CO2· m-2, 净固碳量分别为775.51和575.93 g CO2· m-2。说明围封对于提高草地生产力, 增强生长季群落的碳吸收能力是有效果的。同时, 土壤含水量也是限制群落生产力和土壤生物活性的主导因素[30], 含水量过低或过高都会影响土壤微生物活性[13, 27]及土壤的物理性质和植被的生长[25], 但是本研究结果表明, NEE只在放牧条件下与土壤含水量的相关性达显著水平(P< 0.05), 围封地中生物量大于放牧地, 对土壤的保水作用较大, 在放牧地的水分胁迫程度高于围封地, 土壤水分利用效率也较高。因此, 提高草地植被可利用的土壤含水量也可以促进草甸草原生态系统碳交换。
温度和土壤含水量是影响赖草草地生态系统碳交换的两个重要因子, 生态系统碳交换与温度和土壤含水量存在显著的相关关系, 土壤温度的作用大于土壤含水量。赖草草地生态系统碳交换规律都表现为明显的日变化和季节变化, 均呈单峰曲线; 放牧并没有改变生态系统碳交换的变化规律, 而降低了草地生态系统净碳交换能力, 减弱了草地生态系统碳汇功能, 但围栏封育通过保护植被, 其碳吸收显著高于围栏外放牧草地。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|