
{kind=link}
{kind=link}
{kind=link}
绵羊放牧绢蒿荒漠草地植物群落多样性及其环境解释
[陈乙实1  , 孙海荣
, 孙海荣1 , 李娜娜1 , 靳省飞1 , 车昭碧1 , 曹佳敏1 , 鲁为华1, 2, * ]
, 孙海荣]
|
|


作者简介:陈乙实(1994-),女,黑龙江宾县人,在读硕士。E-mail: 765580080@qq.com
对天山北坡放牧条件下的绢蒿草地进行野外调查,以探讨植物群落物种组成、多样性特征及其与环境因子之间的关系。结果表明:1)绵羊放牧过程中形成了明显的放牧强度梯度,草地利用率适中。通过除趋势对应分析 (DCA)将放牧区内的植物群落划分为8个类型,各类型植物群落的物种多样性有显著差异,且群落间存在异质性;2)采用除趋势对应典范分析法(DCCA)中的前项选择,对所获取的9个环境因子进行筛选,最终土壤有机质含量、粪便量、放牧强度、凋落物生物量、海拔、坡向等6个环境因子成为决定物种分布和群落结构组成的主导因素。DCCA排序结果反映出了实际的生态意义,有机质、放牧强度、粪便量在第一排序轴有明显的梯度变化,而坡向和凋落物则在第2轴上呈现出明显梯度变化,物种和群落也沿着上述环境要素的梯度变化而依次分布,很好地解释了放牧条件下绢蒿荒漠草地物种、群落与环境条件之间的关系。
This study investigated plant community species composition and biodiversity in grazed Seriphidium semidesert grassland, to explore the relationship between community structure and environmental factors. The results showed that: 1) Grazed grassland had an obvious grazing gradient by sheep, and the utilization of grassland was reasonable. A detrended correspondence analysis revealed eight plant communities in the grazed area. There were significant differences in species diversity indexes among plant communities, and there was heterogeneity among communities; 2) Six environmental variables (soil organic matter content, amount of feces, grazing intensity, litter biomass, altitude, and slope direction) were chosen by forward selection. The first axis of a detrended canonical correspondence analysis showed gradients of organic matter content, amount of feces, and grazing intensity, while the second axis showed gradients of litter biomass, altitude, and slope direction gradient among communities. The plant species and communities were distributed along gradients of environmental factors. Our results show that quantitative ecological methods provide more detailed information about ecological relationships between vegetation and the environment for grazed Seriphidium semidesert grassland.
放牧条件下的草地植物物种多样性一直以来是生态学研究关注的焦点问题, 家畜放牧行为参与到了草地群落演替过程中, 成为影响草地物种多样性的重要因素。前人的研究结果已经充分表明, 轻度及中度放牧会明显增加草地植物的物种多样性[1, 2, 3, 4, 5]。也有研究表明, 在肥沃的以及湿润的环境中放牧对植物多样性具有促进作用, 在干旱和相对肥沃的环境中, 则会降低植物多样性, 而在干旱以及贫瘠的环境中, 则对植物多样性没有多大的影响[6]。气候以及放牧被认为是影响草原生态系统稳定性以及维持其功能的重要因子[7]。
在研究放牧过程对草地生态系统影响的过程中, 主要是利用控制试验设置放牧强度, 然后按照放牧强度的梯度变化去探讨物种多样性特征的变化。但在实际生产过程中, 家畜的牧食行为远远比人为设置的处理或者梯度复杂得多, 家畜采食行为在时间和空间上由于各种外在环境条件的变化而产生无法预测的随机变异, 这就使得控制试验的结果不符合实际情况。因此, 在某一草地生态系统中去探讨放牧过程对草地状态的影响, 应该是以现有通用的放牧策略为试验对象, 通过监控牧群的牧食过程并以量化数据去划分在放牧区域内的放牧强度, 然后再以实际放牧过程产生的梯度去探讨草地植物物种多样性特征的变化, 这符合生产实际并且能够得到令人信服的结论。
20世纪90年代末, 学者们将卫星定位导航系统应用到家畜放牧行为的研究中, 起初主要是为了实时监测畜群的位置[8], 后来是为了建设虚拟围栏来控制畜群采食行为[9]。这种实时监测畜群移动状态的系统为开展相关研究打下了良好基础, 大量畜群在空间上的移动可以被这一系统以轨迹点的形式记录并保存, 通过长时间数据序列的累积可以得到放牧区域内的实际放牧强度的时空分布, 通过这一分布状况可以很直观地了解到实际放牧过程产生的放牧强度在该放牧区域的空间梯度变化。
大量研究表明, 气候条件是影响区域内草地植物多样性的主要环境因子[10, 11, 12, 13], 而因此产生的草地微生境的差异更是在草地植物多样性上直接体现出来, 整体气候条件影响着温度、湿度的不同, 微生境则造成了坡度、坡向、土壤紧实度、海拔、pH值等的不同。
按照中度干扰理论, 在零放牧或者轻度放牧的情况下, 控制植物多样性的原因是植物自身之间的竞争, 因无外界干扰或外界干扰较小, 不同植物之间将会在有限的资源条件下进行生存竞争, 弱者淘汰, 结果会导致植物多样性较低; 在适度放牧的情况下, 植物自身之间的竞争力减弱, 增加了对放牧干扰的适应度, 个体竞争力强的植物和对放牧干扰抵抗力强的植物生存下来, 而且放牧增加了环境的异质性, 从而多方面地增加了植物多样性; 但如果是重度放牧, 因践踏、采食、粪便等多种原因影响下, 各类植物都会减少, 而降低植物多样性。
绢蒿属(Seirphidium)荒漠草地是天山北坡山地垂直地带分布的基础植被之一, 在新疆分布面积为114.25× 104 hm2[14]。该类草地一直以来是畜牧业生产的重要春秋牧场, 承担着两季的放牧压力。近年来, 针对绢蒿荒漠严重退化的状况, 学者们对该类草地也进行了大量研究, 但鲜有通过获取放牧家畜的放牧轨迹去划分放牧强度的梯度变化, 然后根据放牧梯度变化系统取样去研究现实放牧强度空间异质性条件下的物种多样性特征。因此, 本研究以合理放牧前提下的绢蒿荒漠草地和羊群为研究对象, 首先利用GPS定位技术获得群体放牧轨迹点, 然后通过轨迹点的空间分布划分现实放牧过程中的放牧强度分布, 并依据不同放牧强度的空间分布状况进行针对性的样方取样, 主要回答以下3个问题:1)现行的放牧制度在放牧场内会形成什么样的放牧强度梯度分布?2)现有放牧条件下, 可形成什么样植物群落类型, 各类型群落的物种多样性特征如何?3)造成物种在不同群落中分布及群落类型的主要环境因子有哪些, 它们与群落和物种分布的关系如何?通过上述问题的回答, 旨在为该类草地的合理放牧策略的制定提供参考。
试验将位于天山北坡的紫泥泉种羊场一大滩绢蒿荒漠放牧地作为野外试验基地(86° 06'-87° 31' E, 43° 27'-44° 05' N)。取样时间为2016和2017年的春秋放牧时间, 该牧场是春秋牧场, 春季放牧时间为4-6月, 秋季为9-11月。该地区是典型的中亚气候型荒漠, 以伊犁绢蒿(Seirphidium transiliense)为优势种, 另外有草原苔草(Carex tristachya)、针茅(Stipa capillata)、羊茅(Festuca ovina)等作为伴生种出现。春季短命和类短命植物可以构成短时优势层片, 主要有顶冰花(Gagea lutea)、角果毛茛(Ceratocephalus orthocera)、郁金香(Tulipa gesneriana)、弯果胡卢巴(Trigonella arcuata)等。
GPS设备选择深圳谷米电子有限公司的GT03C定位跟踪器, 定位误差小于10 m, 整机质量为202 g, 佩带不会对羊的生理行为与采食行为造成影响[15]。利用项圈式GPS定位系统, 每日随机选择5只具有畜群代表性的绵羊佩带GPS项圈进行定位跟踪。获取后的数据利用ArcGIS 10.3进行处理, 得到绵羊每日行走的轨迹点。然后在ArcGIS 10.3中利用其空间分析模块进行轨迹点到放牧强度梯度分布状况的转换。
放牧强度是单位草地面积在一定时间内羊单位数, 以单位面积上放牧羊单位表示。利用Kawamura等[16]的方法, 采用网格法对定位项圈以 3 min为记录时间间隔获得的轨迹点数据进行分析, 生成放牧区域的放牧强度分布图[17]。具体方法为:首先, 对研究区牧场进行网格划分, 根据实际需求, 本研究设定单元格大小为30 m× 30 m; 其次, 对每个单元格内存在的轨迹点进行计数, 进而得到位置点的频数分布。最后, 计算给定时间范围(春秋两季放牧时间段)的放牧强度(grazing intensity, GI)。计算方法如下:
DGI=
式中:DGI为放牧强度, 单位为每平方千米单位羊出现的频次(sheep units, SU), SU· km-2[18]; C为单元格内位置点的总数, 单位为个; N为牧群内羊总数与GPS设备总数的比值, 即每个GPS终端代表羊单位的个数, S为单元格面积(km2)。
利用ArcGIS Desktop的空间分析能力, 借助模型构建器(model builder)以流程化的方式实现放牧强度分析。首先采用“ 创建渔网工具” 对研究区进行网格划分, 再利用“ 空间连接工具” 对给定时间范围轨迹点数据与格网进行处理, 得到给定时间范围内放牧轨迹点频数的空间分布图, 最后使用“ 添加字段工具” 与“ 计算字段工具” 以公式1)计算每个单元格的放牧强度, 进而生成牧场的放牧强度分布。
依据轨迹数据转换成的放牧强度分布图, 确定野外调查取样位置, 取样时以放牧强度分布所显示的放牧路径核心为取样起始位置, 向两侧扩展取样, 涵盖所有放牧强度梯度。共计获得90个样方数据。记录样方内的植物密度、高度、盖度、生物量等基本数据。同时收集地表的凋落物、绵羊排泄粪便, 实地测定土壤湿度、pH值、土壤紧实度等指标, 采集土壤样品, 并进行土壤理化特性分析, 以上述数据作为环境因子指标。
采用Excel 2007软件进行所有数据的初步整理, 采用DPS 7.5进行群落多样性特征、群落相似性比较以及方差分析。利用各群落的物种数量和各物种个体数量进行群落Bray-Curtis相似系数计算。利用Canoco 4.5软件的除趋势对应分析(detrended correspondence analysis, DCA)进行群落类型的初步划分, 利用典范对应分析(canonical correspondence analysis, CCA)分析进行前项选择, 以筛选出主要的环境因子, 然后将样方和环境数据纳入除趋势典范对应分析(detrended canonical correspondence analysis, DCCA)模块进行环境要素和群落数量特征的排序分析, 最终阐明幼苗发生类型和环境要素之间的关系。
Simpson多样性指数(D):
Shannon-Weinerr指数(H'):
Jaccard相似性系数(SJ):SJ=
Bray-Curtis指数(SB):
式中: Pi为物种i在群落中所占比重; a是群落A和群落B都有的物种数量, b是群落B有但群落A无的物种数量, c是群落A中有但群落B中无的物种数量; SB为表征群落间的距离系数, n为样本中物种数量, xij, xik为样本j和样本k中第i种物种的个体数量。在实际计算时, 系统会同时给出Bray-Curtis距离系数和相似性指数, 在这里利用相似性指数作为分析参量。
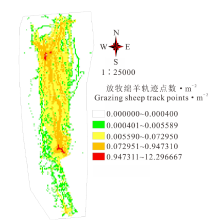
对春季和秋季放牧过程中产生的轨迹点进行整理, 获得了1600余条轨迹数据, 将该轨迹数据以点坐标的形式置入ArcGIS 10.3中, 通过网格分析和频数计算, 最终获得放牧强度的梯度空间分布图(图1)。
 | 图1 牧场放牧强度梯度分布Fig.1 Distribution of grazing intensity gradient |
放牧区总面积为586.5 hm2, 从放牧强度梯度空间分布来看, 其强度分布总体上是由中心位置向两侧依次递减。放牧强度可利用放牧区域内每m2内的放牧绵羊轨迹点数来表征, 可将整个放牧区域按放牧强度由小到大划分为5个放牧梯度, 各放牧强度面积分别占整个放牧区域的62.8%、9.1%、19%、8.8%和0.3%, 草地总体利用率具有明显的梯度变化。
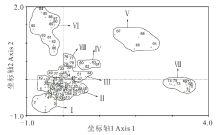
基于上述放牧强度的梯度分布, 按照前文所述取样方法进行样方取样, 将获得的90个样方数据进行整理, 形成样方× 物种的二维数据表, 然后进行基于物种组成的DCA分类, 分类结果见图2。
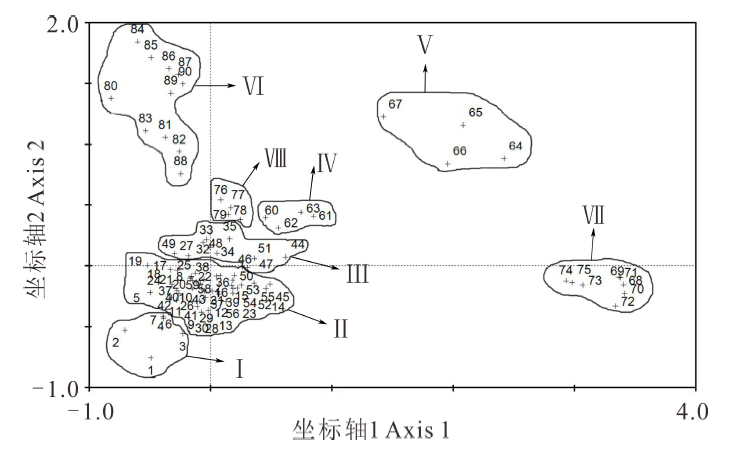 | 图2 90个样方的DCA排序图Fig.2 DCA sorting diagram of 90 sample cubes “ +” : 样方点 Quadrat point. |
从DCA分类图中可以看出, 90个样方被明显地划分为8个类群, 分别为Ⅰ )羊茅+草原苔草+绢蒿群落、Ⅱ )绢蒿+绢蒿幼苗群落、Ⅲ )绢蒿+草原苔草群落、Ⅳ )绢蒿群落、Ⅴ )骆驼蓬(Peganum harmala)+郁金香+顶冰花群落、Ⅵ )骆驼蓬群落、Ⅶ )草原苔草+扁蓄(Polygonum aviculare)群落、Ⅷ )蚤缀(Arenaria serpyllifolia)+角果毛茛+顶冰花群落。
从DCA第1轴排序结果看, 各群落在排序图上呈现有序分布。在实际调查中也发现, 群落Ⅲ 、群落Ⅳ 、群落Ⅷ 主要分布在沟底相对平缓地带, 群落Ⅵ 、群落Ⅴ 主要集中在阳坡区域, 群落Ⅰ 、群落Ⅱ 主要在阴坡区域, 因此第1轴表明的是地形条件的变化; 沿第2轴从群落Ⅵ 至群落Ⅰ 也呈现出明显的梯度变化, 群落Ⅰ 、Ⅱ 、Ⅲ 基本分布在地表粪便数量较少的区域, 而Ⅴ 、Ⅵ 、Ⅶ 则处于地表粪便较多区域, 因此, 第2轴主要反映地表粪便量的梯度变化。
对8个群落的物种多样性指数进行计算, 结果见表1。各多样性指数中, 群落Ⅰ 的物种和物种个体数量最多, 构成群落的物种主要以多年生蒿类半灌木和禾本科植物为主。群落Ⅱ 、Ⅳ 、Ⅶ 物种数为4~6, 以绢蒿、苔草等多年生植物为主要物种; 群落Ⅴ 、Ⅵ 、Ⅷ 包含物种数量最少, 为2~3。群落Ⅰ 、Ⅱ 、Ⅲ 、Ⅳ 、Ⅵ 的香农指数差异不显著, 但显著高于群落Ⅴ 、Ⅶ 、Ⅷ 。群落Ⅰ 、Ⅱ 、Ⅲ 、Ⅳ 辛普森指数要高于其他群落类型, 群落Ⅶ 、Ⅷ 居中, 而Ⅴ 群落最小。群落均匀度在8个群落类型中不存在显著差异, 其范围为0.5~0.8。从多样性特征的分析结果来看, 绢蒿荒漠放牧区的物种数量较少, 但在物种个体总数上不同群落类型存在较大差异。不同群落的香农指数和辛普森指数变化趋势基本相似, 均匀度指数在群落间无显著差异。
| 表1 不同分类群落物种多样性特征比较 Table 1 Comparison of species diversity in different plant communities |
从相似性系数的比较可以看出(表2), 存在明显相似性的群落有3对, 包括群落Ⅱ 与群落Ⅲ 、群落Ⅲ 与群落Ⅷ 、群落Ⅵ 与群落Ⅷ , 其他群落间不存在明显的相似性。结合DCA分类的结果, 相似性特征和DCA分类结果具有良好的一致性, 说明在放牧绢蒿荒漠草地上, 由于多个环境条件在相对较小空间尺度内的变化, 造成植物群落结构组成的复杂性和异质性。
| 表2 不同群落相似性程度比较 Table 2 Comparison of similarity degree of different communities |
尽管在取样过程中按照放牧强度的分布进行梯度取样, 但通过群落分类发现, 该放牧地植被群落组成由于地形条件变化较为复杂, 造成了群落结构的多变性, 从90个样方在坐标轴上分布状况来看, 并未严格按照放牧梯度进行有序分布, 因此, 不能简单地将该草地现实状况的形成仅仅归结于放牧强度, 其群落类型的形成可能与其他环境要素有关, 因此, 有必要对其他环境要素与草地植被群落结构进行进一步的分析。
将物种、样方和环境因子数据进行整理, 利用WcanoImp进行数据转换, 将转换后的数据首先进行CCA分析, 对9个环境因子进行前向选择, 筛选出主要的环境因子, 建立一个更简单的模型(即拥有更少的解释代理变量)。利用Monte Carlo检验, 得出对群落特征产生影响的边际影响和条件影响, 环境因子对植物群落组成特征的影响为边际影响, 通过前向选择、预选择和线性替代, 去除掉相关变量, 得到的特征值为条件影响。如表3所示, 根据环境要素的边际影响排名第一的为有机质, 其次为坡向、紧实度、湿度、粪便量、凋落物。但经过前向选择后, 发现有机质含量依然是对物种构成有最重要影响的因素, 但粪便量、放牧强度和凋落物等环境因子的影响值则大幅提升。在Monte Carlo检验中, 选择有机质含量这一变量, 紧实度、土壤湿度以及pH值条件影响立即降到最低(0.06~0.07), 反映出这两个环境要素与有机质存在强相关性。其余5个环境因子在提出有机质这一因子后, 条件影响值也均出现下降, 但仍然通过了Monte Carlo检验(P< 0.01), 最终有机质、粪便量、放牧强度、凋落物、海拔、坡向通过检验构成代理变量, 共汇集了89%环境信息量, 其中有机质提供了最多的信息量(23%)。
| 表3 预选择中各变量的边际及条件影响 Table 3 Marginal and conditional effects obtained from the summary of forward selection |
在经过预选择中, 确定了有机质等6个代理变量后, 再进行DCCA分析, 得出如图3所示的物种-环境和群落-环境的二维图。排序图的第1轴反映了土壤有机质、粪便量和放牧强度的梯度变化, 自左向右, 上述3个环境变量依次增加, 物种分布也从多年生稳定存在可食物种(如从左侧分布的羊茅、针茅)逐渐向不稳定或不可食物种[如右侧分布的骆驼蓬、顶冰花、二裂委陵菜(Potentilla bifurca)、蚤缀]转变。而排序图的第2轴反映了坡向和凋落物的梯度变化, 自上而下, 坡向由阳转阴, 凋落物量也逐渐增加, 相应地, 物种也逐渐从喜阳物种[如分布在上方的狗尾草(Setaria viridis)、扁蓄、串珠老鹳草(Geraniumtransversale)、小车前(Plantago minuta)]逐渐向喜阴物种[如羊茅、宽刺蔷薇(Rosa platyacantha)、绣线菊(Spiraea salicifolia)]转变。
 | 图3 90个样方的DCCA排序[物种分布(A)和群落结构形成(B)的环境解释] Al:海拔Altitude; Om:有机质Organic matter; Gi:放牧强度Grazing intensity; Fe:粪便量Dung; Px:坡向Exposure; Li:凋落物Litter. 狗尾草S. viridis; 扁蓄P. aviculare; 球根老鹳草G. transversale; 平车前Plantago depressa; 角果藜Ceratocarpus arenarius; 画眉草Eragrostis pilosa; 草原苔草C. liparocarpos; 沙蓬Salsolacollina; 蒲公英属Taraxacum; 蚤缀A. serpyllifolia; 二裂委陵菜P. bifurca; 骆驼蓬P. harmala; 败酱属Patrinia; 棘豆属Oxytropis; 荒漠庭荠Alyssum; 羊茅F. ovina; 灰绿藜Chenopodium glaucum; 绢云母苗Seriphidium seedling; 伊犁绢蒿S. transiliense; 针茅Stipa capillata; 红杆蒿Kochia prostrata; 紫毛蕊花Verbascum phoeniceum; 伊犁郁金香Tulipa iliensis; 棱叶韭菜Allium caeruleum; 顶冰花G. lutea; 宽刺蔷薇R. platyacantha; 金丝桃叶绣线菊Spiraea hypericifolia; 冰草Agropyron cristatum.Fig.3 DCCA sorting diagram of 90 sample cubes [environmental interpretation of species distribution (A) and community structure formation (B)] |
群落分布在二维排序图上也有其规律性, 在引入6个环境变量后, 各样方相对位置只发生了略微的变化, 其中在DCA排序图中(图2)的Ⅱ 、Ⅲ 、Ⅳ 群落合并, 该合并类型主要受到凋落物、坡向和海拔高度的影响。群落Ⅴ 主要受到粪便和放牧强度的影响, 群落Ⅵ 是在海拔高度的作用下形成的, 群落Ⅶ 主要受到粪便量的影响, 群落Ⅷ 的结构组成主要由放牧强度所决定。通过DCCA分析可以很好地进行各群落类型形成的环境解释, 并可将群落类型形成的与主导环境因子分别提取出来。
在解释环境变量对物种和群落分布的影响时, 进一步对排序特征值和物种与环境因子的相关系数进行了计算, 结果见表4。经过前向选择后, 6个环境因子的信息基本集中在了第1轴和第2轴上, 2个环境轴提供了75%的环境信息量, 且均很好地通过了Monte Carlo检验(P< 0.01), 说明所筛选出的代理变量非常有效, 重要的环境因子在采用代理变量分析时未被遗漏, 同时也代表着大部分的环境信息。皆说明使用代理变量的合理性, 同时能够直观地反映研究区域的植物群落分布特征与主要环境因子之间的关系。
| 表4 经过前向选择后DCCA分析排序特征值及物种与环境因子相关性分析 Table 4 Eigenvalues and species-environment correlation coefficients for the four axes of DCCA |
绢蒿荒漠草地在北疆天然草地放牧畜牧业中有着重要地位, 承担着两季的放牧压力, 因此, 探讨其在放牧条件下的草地群落多样性特征对于该类草地的合理利用具有重要的现实意义。以实际放牧强度为基本依据, 进行取样、后续分析以及放牧制度制定或改进是最符合生产实际的有效途径[19, 20, 21]。
在自然放牧草地上, 放牧强度并非是决定草地植物群落多样性的唯一决定性要素[22], 尤其在山区草地, 由于地形条件的变化造成水热状况再分配是草地产生分异的本底原因[23]。本研究中之所以将放牧强度这一要素作为基础要素进行系统取样, 就是因为考虑这一要素是最典型的人为驱动过程, 在系统取样时容易在空间上进行样点设置。本研究中基于放牧强度空间分布得到的90个样方数据, 在DCA分析中被分成8个群落类型, 而8个群落类型所包含样方数量差异很大, 从侧面也反映了群落结构的形成受众多因素影响, 放牧过程仅仅是众多影响因素其中的一个方面[24, 25]。从进一步的群落多样性指数及群落相似性比较中也反映出群落结构形成的复杂性。
影响植物多样性的环境条件很多, 相关研究表明[10, 11, 12, 13, 26], 在大区域及全球范围内, 气候条件是影响植被类型及生活型的主要因子; 在景观或群落尺度上, 地形(海拔、坡度、坡向)、土壤(凋落物、有机质含量、营养元素含量、pH值等)等环境因子是决定植被分布格局的主导因子[27]。但在实际研究过程中, 并不是环境因子越多越好, 产生的冗余变量可能会对结果造成干扰, 因此, 使用前向选择法和Monte Carlo显著性检验, 找出存在共线性关系的环境因子能准确地提取对群落影响显著的环境因子, 排除存在共线性的环境因子, 减少了环境要素和计算工作量, 大量研究已经证明了该方法的合理性[28]。本试验即从9个环境变量中提取了6个环境因子作为代理变量。代理变量共提取出89%的环境信息, 其中土壤有机质所蕴含的环境信息量最多, 达到23%, 随后依次是放牧强度、粪便量、凋落物、海拔和坡向。
本研究中土壤有机质含量、坡向、凋落物等因素的综合影响效果较放牧强度这一单一变量的影响效果更明显。坡向和坡度也是影响植物群落组成和分布的主要环境因子, 一般来说, 阳坡的温度、光照较阴坡充足, 但是湿度较低, 因坡度和坡向的不同产生很多小的生境条件特征, 也会造成明显的群落组成差异。凋落物的影响是呈正负双向的[29, 30, 31], 正向影响为凋落物增加了地面的粗糙程度, 便于地面对种子的获取, 但负向影响则表现为因凋落物增加了地面厚度, 会阻碍种子萌发出苗。粪便的数量和分布对植物群落多样性有着显著的影响, 放牧家畜粪便在空间内的分布可直接影响草地植物群落组成甚至生产力的改变[32]。海拔是环境影响因子的另一主导因子[33], 尤其是大部分研究人员将海拔梯度变化看做是小尺度范围内纬度变化的缩影, 对于植被分布、物种多样性的海拔梯度研究提供重要依据[27]。
物种分布模式和群落结构是反映植物群落形成过程的两个重要尺度。首先, 物种分布模式可以从源头上阐释群落内物种组成的机理及原因[34], 对环境条件需求的同质性是促成物种组合形成的关键, 而物种分布的DCCA分析是解释这一现象的最好工具[35]。从本研究的物种分布排序图来看, 不同物种对环境要素的选择偏好在二维排序图中分布界限明显, 从另一侧面讲, 植物可以作为不同生境的“ 指示者” [36], 如本研究中的骆驼蓬就指示了其分布区家畜高强度采食(GI)和高粪便积累量(Fe)这一环境特点。狗尾草、扁蓄、串珠老鹳草、小车前等主要分布在阳坡的较干旱地区, 而羊茅、宽刺蔷薇、绣线菊等物种主要分布在阴坡较湿润地区。其他物种也相应地指示出各自所在地的基本环境条件。其次, 群落形成是物种和物种、物种和环境相互选择的结果, DCCA分析可以将这种互相选择后的群落差异在二维排序图中显示出来, 并明确每一群落是由哪种或几种环境要素主导影响而形成, 这为群落形成的环境要素解释提供了很好的工具。本研究中的6个环境要素所主导的群落类型在二维图上就有很清晰的界限, 并能和物种分布产生很好的对应。
本研究从这一角度出发, 因绵羊在放牧过程中形成的放牧梯度, 利用GPS定位技术将羊群的采食路径进行记录并与放牧区卫星影象叠加, 利用ArcGIS中的空间分析和网格计算确定出区域内的放牧强度, 然后再进行基于放牧强度的系统取样和后续相关数据分析, 使试验方法合理, 分析过程有针对性, 最终结果符合生产实际[37, 38]。从放牧强度分布图可以直观反映出时间和空间上的草地利用状况, 本研究的放牧强度划分结果也明显反映出自然放牧状态下草地利用在空间上的不均匀性, 正是这种非均一的利用方式才会使整个放牧区域的植物群落产生分化而形成多种类型, 这种影响甚至会使草地在景观尺度上显示出明显的异质性[39, 40]。
1)绵羊在放牧过程中形成了明显的放牧梯度, 实际发生的不同放牧梯度可以作为空间内系统取样的依据, 使取样更符合生产实际; 2)通过DCA分析, 将研究区内草地落划分成8个群落类型, 每个群落类型之间具有明显的界线, 各自的多样性指数亦存在差别, 并且各个群落之间无明显的相似性; 3)采用前向选择和Monte Carlo检验筛选出了6个影响物种分布和群落形成的主要环境因子, 各个环境因子对物种分布和群落形成具有显著的影响, DCCA排序结果反映了各个环境因子的实际生态意义, 对放牧绢蒿荒漠草地物种、群落特征做了很好的环境解释。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|