
{kind=link}
{kind=link}
基于遗传多样性评估燕麦品种的农艺性状
[王建丽1, 2  , 马利超
, 马利超3 , 申忠宝2 , 刘杰淋2 , 朱瑞芬2 , 韩微波2 , 钟鹏4 , 邸桂俐2 , 韩贵清1, 2 , 郭长虹1, * ]
, 马利超]
|
|


作者简介:王建丽(1977-),女,内蒙古赤峰人,助理研究员,在读博士。E-mail: wangjianlivip@126.com
为客观评价燕麦种质资源重要农艺性状的遗传多样性,并为黑龙江地区燕麦新品种选育提供关键数据,对51份燕麦种质资源13个农艺性状进行了形态多样性指数分析,并对其中的9个数量性状进行了聚类分析和主成分分析。结果表明,各性状的遗传多样性指数均较大,遗传多样性指数最高的是主穗长(1.517),其次为株高(1.448)和主穗粒重(1.414),性状变异系数最大的是主穗小穗数(34.8%),其次为主穗粒重(33.1%)和单株分蘖数(27.4%);聚类分析将51份燕麦品种的9个数量性状分为4大类群,类群Ⅰ为有益性状不明显,为多目标性状育种的亲本材料,类群Ⅱ为选育矮秆育种目标亲本材料,类群Ⅲ为高杆、增加分蘖数育种目标亲本材料,类群Ⅳ为选育大粒型、多轮层数、多小穗数等育种目标亲本材料。主成分分析结果表明,前3个主成分对变异的累计贡献率为70.09%,第一主成分反映种子产量,第二主成反映单株分蘖数,第三主成分反映株高。
, MA Li-chao
This research aimed to objectively evaluate the genetic diversity of oat germplasm resources for important agronomic traits, and to provide key data for use in oat breeding programs in the Heilongjiang region. A total of 51 oat germplasm accessions from different sources were planted in Harbin at the High Latitude Grass and Science Institute, Heilongjiang Academy of Agricultural Sciences. Genetic diversity indices for the various accessions were calculated, and cluster analysis and a principal component (PC) analysis were carried out to investigate patterns of genetic diversity across accessions for agronomic characters. The index of genetic diversity was high for all traits, with the highest being for the length of the main spike (1.517), followed by plant height (1.448) and seed weight of the main spike (1.414). The largest coefficient of trait variation was for spikelet number of the main spike (34.8%), followed by seed weight of the main spike (33.1%), and tiller number per plant (27.4%). Based on cluster analysis, nine quantitative traits could be used to classify plants into four groups. Plants in germplasm group 1, contained no obviously attractive traits, while group II contained the germplasm that could be used as parents of short-stemmed varieties. The germplasm in group III typically had greater stem length and increased tiller number, compared to others. In contrast, the germplasm in group IV was suitable for breeding cultivars with large grain size, multiple panicle and spikelet numbers .According to principle component analysis, the first three main component (seed yield factor, till factor, plant height factor) made the contribution of 70.09% to variation.
燕麦(Avena sativa)属于禾本科燕麦属一年生草本植物, 具有产量高、营养全面、耐寒、耐盐碱、适应性广等特性, 在冷凉地区生长良好[1]。燕麦不仅是一种优质的粮食和经济作物, 也是各种家禽、家畜喜爱的优质饲料。目前, 燕麦主要分为裸粒型和带稃型两种[2]。在我国, 栽培的燕麦主要以裸燕麦为主。目前燕麦市场存在以下特点:品种繁多, 应用推广缓慢, 产业化落后。主要原因在于燕麦的育种受到种质资源和育种技术的限制, 尤其种质资源遗传关系的不明确, 利用效率低, 改良技术不完善等问题, 导致市场上鲜有具备优良特性的新品种。
利用分子生物学技术鉴定和评价燕麦种质资源对于燕麦遗传育种具有重要意义, 国外学者在这方面取得了较大进展[3, 4, 5, 6, 7, 8]。相比而言国内学者主要关注燕麦种质资源的收集和评价[9, 10, 11, 12] 、生产性能及栽培技术研究[13, 14, 15], 对于种质资源遗传多样性的报道很少[16, 17, 18, 19, 20, 21]。近年来, 我国的燕麦产出远不能满足当下畜牧业养殖的需求, 2018年1-3月, 我国进口燕麦草6.16万t, 预计全年将进口燕麦草超35万t, 因此, 燕麦草产业在我国发展空间巨大。特别是黑龙江省具有独特的地理环境与气候条件, 非常适宜燕麦生产, 深受草企业及有机牧场的青睐。本试验选择 51份燕麦材料在哈尔滨地区种植, 通过观察数量性状和质量性状, 对51份不同生态型燕麦种质资源的形态性状进行遗传多样性、聚类及主成分分析, 揭示燕麦种质资源亲缘关系及变异的特点。评估结果将对合理利用燕麦种质资源、综合评价亲本、燕麦耐寒种质资源新基因的挖掘及种质创新均具有重要意义。
供试材料为 51 个燕麦品种, 包括31份皮燕麦和20份裸燕麦(表1)。
| 表1 51份燕麦品种来源及其皮/裸性 Table 1 Sources and husk/naked of 51 oat cultivars |
试验于2018年在哈尔滨市道外区民主乡黑龙江省农业科学院草业研究所试验地内进行。该地区地理位置为北纬45.2° , 东经122.6° , 海拔169 m。年平均气温为5.1 ℃, 极端最高气温 37.5 ℃, 最低气温 -35.5 ℃, ≥ 10 ℃ 年活动积温2887 ℃, 年平均降水量469.1 mm, 年平均无霜期139 d左右, 土壤为黑土, pH值为7.0~7.5, 全氮含量1.23 g· kg-1, 全磷含量0.47 g· kg-1, 速效氮含量83.51 mg· kg-1, 速效磷含量18.32 mg· kg-1, 前茬作物为籽粒苋(Amaranthus paniculatus)。供试材料于4月5日播种, 采用随机区组设计, 3 次重复, 每小区11行, 行长5 m, 行距30 cm, 小区面积3 m× 5 m=15 m2, 小区间距60 cm, 燕麦出苗至成熟期进行人工除草 3次。
在种子成熟期随机抽取10株进行形态性状的观察, 数据采集及记载严格按照燕麦种质资源描述规范和数据标准进行[14]。田间观察和测量指标包括株高、单株分蘖数、茎粗、主穗穗长、主穗小穗数、主穗粒数、主穗粒重、轮层数、千粒重、株型、粒色、粒形、皮/裸性共13个性状。
采用Excel和SPSS 22.0软件进行数据统计分析。质量性状主要计算各性状的多样性指数和频率, 数量性状主要计算平均值、变异系数、标准差、极差、最大值、最小值和遗传多样性指数, 遗传多样性指数采用Shannon-Weaver指数(H'), 计算公式:多样性指数
对51份燕麦推广品种4个质量性状进行遗传多样性分析(表2), 结果表明, 籽粒颜色遗传多样性指数最高, 为0.960, 以浅黄色居多, 分布频率为0.588, 说明该性状遗传差异较大, 遗传基础比较广泛; 粒形以纺锤形为主, 频率为0.569; 株型以披散为主, 频率为0.745; 皮裸性以皮燕麦为主, 频率为0.607; 由表2可知, 在4个质量性状中, 遗传多样性指数大小为粒色> 粒形> 皮裸性> 株型。
| 表2 燕麦种质资源4个质量性状的遗传多样性 Table 2 Analysis on genetic diversity of 4 quality characteristics in oat germplasm |
对供试51份燕麦推广应用品种9个数量性状的遗传多样性进行统计分析(表3), 结果表明, 株高变化范围 66.5~131.3 cm, 单株分蘖数变化范围 1.0~3.7, 茎粗变化范围 1.5~3.6 mm, 主穗长变化范围8.8~25.0 cm, 主穗小穗数变化范围 7.7~35.3, 主穗粒数变化范围60.3~141.0, 主穗粒重变化范围1.4~5.8 g, 轮层数变化范围4.0~7.3层, 千粒重变化范围 18.7~43.6 g。主穗长的遗传多样性指数最高, 为 1.517; 单株分蘖数的遗传多样性指数最低, 为0.980。9个数量性状的多样性指数平均值 1.285, 说明各性状具有丰富的多样性, 遗传基础广、差异大。主穗小穗数和主穗粒重的变异系数较大, 其值分别为34.8%、33.1%, 表明选择的燕麦品种间在这两个性状上存在较大的差异, 变异类型丰富, 选择的基础较广, 品种改良的潜力大。轮层数的变异系数最小, 为11.8%, 说明此性状在所选择的燕麦品种间差异较小, 比较稳定。对种子产量起关键作用的主穗粒重、主穗小穗数、主穗长以及千粒重的变异系数差异较大, 说明参试品种间变异类型丰富, 亲本选择范围广。
| 表3 燕麦种质资源9个数量性状的遗传多样性 Table 3 Analysis on genetic diversity of 9 quantity characteristics in oat germplasm |
利用SPSS 22.0软件对51份燕麦推广品种的9个数量性状用最长距离法进行聚类分析(图1)。
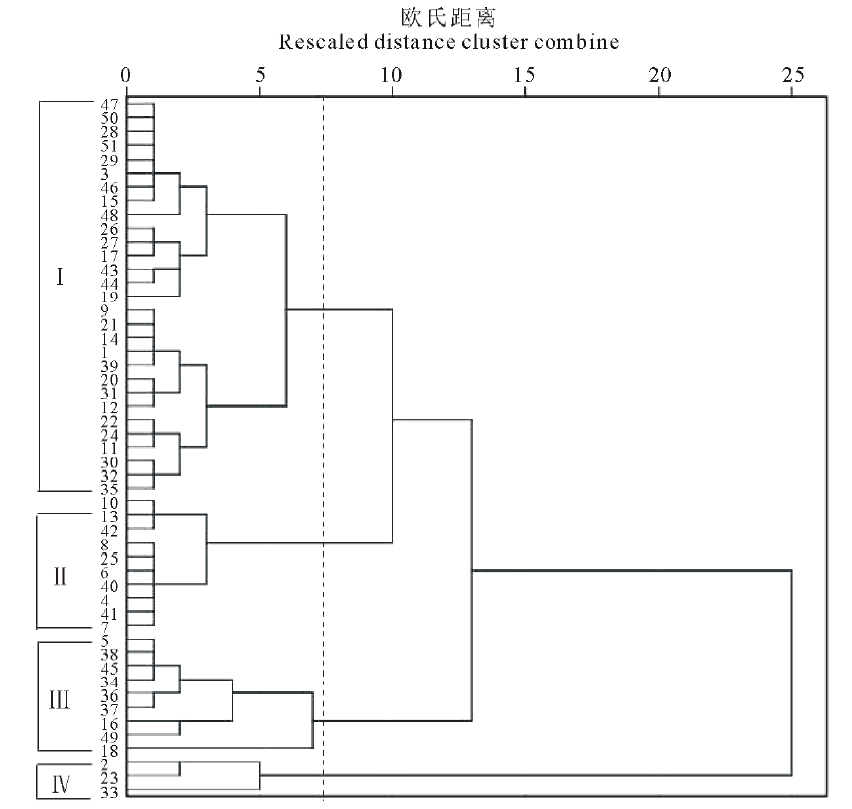 | 图1 51份燕麦种质资源形态性状聚类图Fig.1 Dendrogram generated by cluster analysis for 51 oat germplasm |
在聚类分析过程中, 当欧氏距离系数为6.8时, 51份材料共分为4个类群。各类群的形态特征见表4。
| 表4 燕麦种质资源各类群的生物学性状 Table 4 Characteristics of biological character traits in oat germplasm |
类群Ⅰ 包括29份材料, 编号分别是1、3、9、11、12、14、15、17、19、20、21、22、24、26、27、28、29、30、31、32、35、39、43、44、46、47、48、50、51。这一类株高、单株分蘖数、茎粗、主穗小穗数、主穗粒重、轮层数和千粒重均处于中等水平, 粒色以浅黄色为主, 粒形以纺锤形为主, 皮燕麦居多, 主穗长在4大类群中最短, 其值为13.61 cm, 茎粗、主穗长、主穗小穗数变异系数在4大类群中最大, 分别为19.75%、19.16%、10.68%。
类群Ⅱ 包括10份材料, 编号分别是4、6、7、 8、10、13、25、40、41、42。 该类群株高、茎粗、主穗小穗数、主穗粒重、主穗粒数和轮层数在4大类群中最低, 其值分别为81.72 cm、2.34 mm、14.90个、1.76 g、65.40个、5.35, 粒色多为浅黄色和黄色, 粒形以披针形为主, 皮燕麦为主, 株高、主穗粒数和千粒重变异系数在4大类群中最小, 分别为11.13%、5.9%和15.28%, 可作为选育矮秆皮燕麦目标的优良亲本材料。
类群Ⅲ 包括9份材料, 编号分别为5、16、18、34、36、37、38、45、49。 这一类群品种以裸燕麦为主, 粒色为浅黄色和黄色, 粒形披针形为主。此类材料中, 单株平均分蘖数最多为2.93个, 变异系数最低为18.20%, 千粒重最低为26.42 g, 主穗长、主穗粒重变异系数在4大类群中为最小, 其值分别为15.26%、13.35%。
类群Ⅳ 包括3份材料, 编号分别是2、23和33。 这3个品种粒色均为浅黄色皮燕麦。这类材料中, 除单株分蘖数在4个类群中最低为2.00个外, 其余8个数量性状均处于最高, 平均株高101.90 cm、茎粗3.13 mm、主穗长21.57 cm、主穗小穗数24.67个、主穗粒数133.20个、主穗粒重4.30 g、轮层数6.43层、千粒重35.53 g, 主穗小穗数变异系数最大为52.64%, 可作为选育大粒型、多轮层数、多小穗数等育种目标的皮燕麦亲本材。
对燕麦品种9个数量性状进行主成分分析, 结果表明(表5), 在9个主成分构成中, 主信息主要集中在前3个主成分, 其累计贡献率达到了70.09%。主成分一贡献率最大, 其值为36.50%, 其次为主成分二、三, 贡献率分别为19.99%、13.60%。
| 表5 燕麦主要农艺性状主成分分析 Table 5 Principal component analysis of main quality properties in oat |
第一主成分特征值为3.285, 贡献率为36.50%。特征向量中符号均为正数, 载荷较大的性状有主穗粒重、主穗小穗数、主穗粒数、轮层数及茎粗(图2), 其向量值分别为0.800、0.719、0.667、0.661和0.659, 此类性状均与种子产量有关, 可作为产量构成因子, 单株分蘖数的载荷较小。说明在高产育种工作中, 主穗粒重、主穗小穗数、主穗粒数、轮层数及茎粗可进行独立选择, 株高和千粒重要适中, 单株分蘖数越少越好。
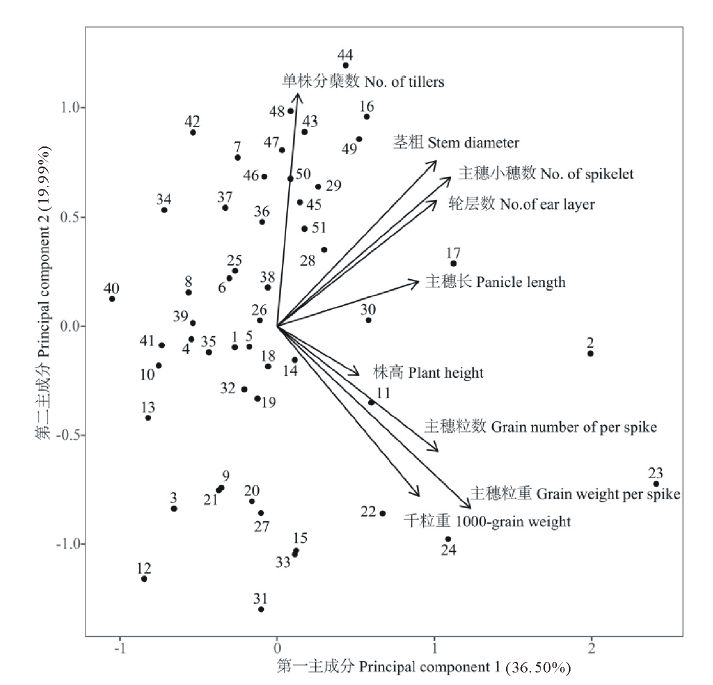 | 图2 51份燕麦种质资源9个数量性状的二维坐标图Fig.2 Two-dimensional chart of 9 quantitative characters in 51 oat germplasm |
第二主成分特征值为1.799, 贡献率为19.99%。特征向量符号中有正有负, 主要反映单株的分蘖数(图2), 其向量值为0.693, 由载荷数值可以看出, 分蘖数影响产量构成因子主穗小穗数、轮层数, 同时对千粒重、主穗粒重和主穗粒数等因子起到了制约作用。因此在育种的过程中, 要适度把握。
第三主成分特征值为1.224, 贡献率为13.60%。特征向量符号中有正有负, 载荷最高的数量性状是株高, 其向量值为0.840, 说明第三主成分主要反映株高, 随着株高的增加会影响主穗小穗数和千粒重, 从而影响种子产量, 所以第三主成分分值越高, 越适合饲用燕麦育种。
我国燕麦主要分布在内蒙古、河北、山西、甘肃、青海及吉林等地区。经过多年的研究发现, 黑龙江地区也非常适宜燕麦生产[22], 但目前燕麦遗传基础狭窄, 不利于其稳定发展。本试验选用51份燕麦材料, 31份来自国内(华北地区11份:内蒙古2份、河北7份、山西2份; 西北地区17份:甘肃13份、青海4份; 东北地区3份:吉林3份), 20份来自国外(美国、欧洲、澳大利亚等燕麦产区的主打品种)。这些材料大部分来自高纬严寒地区, 与黑龙江地区地理环境相近, 这种有目的性的引种, 对黑龙江地区燕麦新品种选育具有重要意义。
燕麦种质资源农艺性状的遗传多样性分析已被广泛应用于燕麦的遗传改良中[23]。本试验对51份燕麦推广应用品种的13个性状的遗传多样性进行了分析, 结果表明, 4个质量性状的遗传多样性指数以粒色最高, 而9个数量性状的变异系数均大于10%, 表明这些性状在种质个体间差异较大, 变异系数以主穗小穗数最大, 说明不同品种间的遗传变异程度不一致, 可为燕麦品种改良提供丰富、优良的亲本材料, 同时可拓宽燕麦新品种选育空间和方向。9个数量性状的遗传多样性指数以主穗长最高, 说明主穗长这个性状遗传基础广、变异丰富。
借助聚类分析的方法, 对 51 份燕麦种质资源的 9个数量性状进行了聚类, 使其性状相近的聚为一类, 结果表明, 来自同一区域的材料并没有聚为一类, 可能与不同区域相互引入有关, 说明材料之间的遗传差异不一定与地理来源有关, 这与齐冰洁等[19]、王鸣刚等[24] 的研究结果相似。不同于上述报道, 本研究把51份燕麦推广品种种质分为4类, 类群Ⅰ 中包括了大部分国外引进品种, 茎粗、主穗长、主穗小穗数变异系数在4大类群中最大, 有益性状不明显; 类群Ⅱ 株高、茎粗、主穗小穗数、主穗粒重、主穗粒数和轮层数在4大类群中最低, 皮燕麦为主, 可作为选育矮秆皮燕麦目标的优良亲本材料; 类群Ⅲ 为选育高秆、增加分蘖数育种目标亲本材料; 类群Ⅳ 除单株分蘖数在4个类群中最低外, 其余8个数量性状均处于最高, 可作为选育大粒型、多轮层数、多小穗数等育种目标的皮燕麦亲本材。这样, 本研究明确了 51 份燕麦种质资源的不同生态型, 在随后的品种选育过程中, 可根据育种目标选择相对互补的亲本配置组合, 使燕麦育种在亲本的选择上更加科学。
主成分分析法已在小麦(Triticum aestivum)[25] 、大豆(Glycine max)[26]和燕麦[12]等作物上应用, 不仅可以掌握作物综合形态性状, 而且可以简化选择程序。本试验将选用的51份材料中9个数量性状进行主成分分析, 前3个主成分对变异的累计贡献率达到70.09%, 第一主成分反映种子产量, 载荷较高的数量性状有主穗粒重、主穗小穗数、主穗粒数、轮层数, 说明在高产育种工作中, 主穗粒重、主穗小穗数、主穗粒数及轮层数可进行独立选择; 第二主成分载荷最高的数量性状是单株分蘖数, 其对千粒重、主穗粒重和主穗粒数等产量构成因子起到了制约作用, 因此在育种的过程中, 要适度把握; 第三主成分载荷最高的数量性状是株高, 株高是燕麦新品种选育推广的重要指标, 其对主穗小穗数、千粒重和轮层数等产量构成因子起到了制约作用, 所以第三主成分分值越高, 越适合饲用燕麦育种。
综上所述, 在黑龙江地区燕麦育种过程中, 亲本材料的选配可以根据主成分排序, 具体全面评价每个亲本材料综合指标的优势, 根据制定的育种目标, 结合聚类分析, 合理地选配组合, 可以加快育种进程, 更早选育出适合黑龙江地区种植的高产、优质燕麦新品种。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|