

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
γ-氨基丁酸对高温胁迫下黑麦草光合特性及碳水化合物代谢的影响
[王日明1  , 王志强
, 王志强2, * , 向佐湘3, * ]
, 王志强, 向佐湘]
|
|


作者简介:王日明(1971-),男,湖南邵阳人,副教授,博士。E-mail: wangriming@qzhu.edu.cn
以热敏感品种顶峰(Pinnacle)为试验材料,研究了热胁迫下(35 ℃/30 ℃,昼/夜)施用外源γ-氨基丁酸(GABA)对黑麦草生长、光合及叶绿素荧光特性、碳水化合物含量及其代谢关键酶基因表达的影响。结果表明,热胁迫造成黑麦草生长减弱,膜透性增加,光合色素含量下降,光合能力和碳水化合物含量显著降低,糖代谢关键酶(蔗糖合成酶、蔗糖磷酸合成酶和转化酶)基因在短时间内(5 d)表达显著上调,随后下降;外源施用GABA显著缓解了高温对黑麦草生长的抑制作用,提高了高温胁迫下光合色素和碳水化合物含量,净光合速率( Pn)、气孔导率( Gs)、蒸腾速率( Tr)、CO2羧化效率(CE)、最大光合速率( Amax)和光化学效率( Fv/ Fm)、Rubisco最大羧化速率( Vcmax)和RuBP再生速率( Jmax)、光化学猝灭系数( qP)、电子传递速率(ETR)和PSⅡ实际光化学效率( ΦPSⅡ)显著升高,而气孔限制值( Ls)、CO2补偿点(CP)和非光化学猝灭系数(NPQ)显著降低。施用外源GABA降低了高温胁迫下叶片气孔和非气孔限制值,缓解了高温对叶片PSⅡ反应中心的抑制,增强了糖代谢关键酶基因表达的水平,提高了碳水化合物含量,因此有效的提高了黑麦草的耐高温能力。
The aim of this research was to elucidate the effects of exogenous γ-aminobutyric acid (GABA) on high temperature stress metabolism, plant growth, photosynthetic and chlorophyll fluorescence characteristics, carbohydrate content and gene expression levels of key enzymes associated with sugar metabolism. The material studied was a heat sensitive cultivar (Pinnacle) of perennial ryegrass ( Lolium perenne) under high temperatures (35/30 ℃ light/dark) in growth chambers for 15 d. It was found that heat stress inhibited plant growth, increased leaf electrolyte leakage and decreased carbohydrate and chlorophyll content, photosynthesis rate. The gene expression levels of key enzymes associated with sugar metabolism were initially up-regulated and subsequently decreased under heat stress. Exogenously applied GABA significantly alleviated the damage effect of heat stress on plant growth and enhanced the chlorophyll content and carbohydrate content under high temperature. The leaf net photosynthetic rate ( Pn), stomatal conductance ( Gs), transpiration rate ( Tr), carboxylation efficiency (CE), maximum assimilation ( Amax), photochemical efficiency ( Fv/ Fm), maximum velocity of carboxylation ( Vcmax), maximum electron transfer rate ( Jmax), photochemical quenching ( qP), electron transport rate (ETR) and actual photochemical efficiency of PSII ( ΦPSⅡ) significantly increased while leaf stomatal limitation (Ls), compensation point (CP) and none photochemical quenching (NPQ) significantly decreased after application of GABA under heat stress. To summarize, exogenous application of GABA reduced leaf stomatal and non-stomatal limitations to photosynthesis and alleviated the inhibition of PSII reaction centers, leading to an enhancement in light and dark reactions of photosynthesis and in photosynthetic potential. Expression levels of key genes for sugar metabolism were also enhanced. These physiological changes acted cumulatively to confer a higher tolerance to heat stress in perennial ryegrass.
草坪和草坪草在现代城市发展和人们日常生活中发挥着越来越重要的作用。温度是影响草坪草生长发育的重要环境因子, 在南方亚热带过渡性气候区6-8月份气温长期高达35~38 ℃, 而对于大多数的冷季型草坪草种和品种来说都存在难以越夏的问题。多年生黑麦草(Lolium perenne)是庭园和城市园林绿化以及足球场运动草坪建植中常用的冷季型草坪草种, 因其具有草坪质量优良、分蘖性能好、成坪迅速等优点而常用作草坪建植的先锋草种。黑麦草的最适生长温度为15~25 ℃, 当气温长时间高于35 ℃时草坪的生长发育受阻而造成草坪质量下降甚至死亡[1, 2]。
光合作用是植物对外界环境条件最为敏感的生理反应之一。光合作用的各个环节包括光能的吸收、传递和转换等光反应过程、CO2气体交换及固定和还原等暗反应过程常受到高温胁迫的影响[3]。高温胁迫下植物光合作用的下降, 既有气孔因素的限制, 同时也受高温胁迫下植物细胞内PSⅡ 电子传递和反应中心结构等受损、Rubisco羧化效率和RuBP再生速率下降等非气孔因素的限制[4]。糖是植物生长发育过程中的重要能量提供者, 同时也对植物的抗逆性具有重要调控作用。植物体内非结构性糖既是植物细胞内代谢的重要底物, 也是光合与生长利用之间主要的中间贮存产物[5]。可溶性糖不仅可作为渗透调节物质, 还可以作为抗氧化剂以维持和保护逆境条件下植物细胞膜结构的完整性[6]。高温胁迫下植物叶片光合能力降低, 光合产物减少, 从而糖代谢发生改变, 而蔗糖合成酶(sucrose synthase, SS)、蔗糖磷酸合成酶(sucrose phosphate synthase, SPS)和转化酶(invertase, INV)是糖代谢过程中的关键酶, 对细胞内蔗糖、葡萄糖和果糖的合成与分解起着关键性的作用[7]。
γ -氨基丁酸(γ -aminobutyric acid, GABA)是一种在动植物体内均有分布的四碳非蛋白质氨基酸, 可作为外源小分子物质参与植物的生长发育和逆境胁迫调控[8]。当植物受到水分、温度、盐碱等一些逆境胁迫时, 植物体内GABA的含量大量增加而抗逆性提高[9]。外源GABA 既可作为临时氮库, 还可以清除活性氧和自由基、调节细胞内的pH和渗透压等作用以提高植物的抗逆性[10]。已有研究表明, 外源GABA 能够提高番茄(Lycopersicon esculentum)[10, 11]、网纹甜瓜(Cucumis melo)[12]、小麦(Triticum aestivum)[13]、玉米(Zea may)[14]、白三叶(Trifolium repens)[15]、黄瓜(Cucumis sativus)[16]等植物的抗旱、耐盐、耐寒等抗逆能力。Li 等[17]研究发现外源施用GABA可通过调控氨基酸、有机酸的代谢水平而提高冷季型草坪草匍匐剪股颖(Agrostis stolonifera)的耐旱和耐热能力, 但是关于外源GABA 能否通过调控光合和叶绿素荧光特性及碳水化合物代谢水平来提高黑麦草耐高温能力的研究还未见报道。因此, 本研究在前期试验的基础上, 以耐热性弱的黑麦草品种为试验材料, 通过喷施和浇灌外源GABA, 研究了外源GABA对高温胁迫下黑麦草的生长、光合作用、叶绿素荧光特性、碳水化合物含量及其代谢关键酶基因表达的影响, 为解析外源GABA增强植物的抗逆性和多年生黑麦草的抗逆机理提供理论基础。
采用耐热性弱的多年生黑麦草品种‘ 顶峰’ (L. perenne cv. Pinnacle)为材料[18]于2017年9-11月进行试验。
黑麦草品种‘ 顶峰’ 种子经发芽试验确定发芽率, 然后将种子播种于装有栽培基质的塑料花盆中(上口径15 cm× 下口径10 cm× 高15 cm), 培养基质为泥炭土和河砂(1∶ 1, 体积比), 然后置于湖南农业大学教学科研试验温室中培养。幼苗长至高约8 cm时进行修剪, 修剪高度保持约6 cm, 每周每盆浇一次1/2浓度的Hoagland营养液, 每次浇200 mL。
将在温室中培养至冠层和根系基本建成(约45 d)的各盆栽材料转移至光照培养箱中进行预培养, 培养条件设为昼/夜温度为25 ℃/20 ℃, 65%~75%的相对湿度, 12 h光照时间和400 μ mol· m-2· s-1的光照强度。前期预试验结果发现5 mmol· L-1 GABA对黑麦草缓解高温胁迫具有最好的效果。盆栽材料在生长箱预培养3 d后, 用5 mmol· L-1的GABA溶液每盆喷施100 mL并同时浇灌150 mL, 2 d后开始进行高温处理。对照昼夜温度设为25 ℃/20 ℃, 高温处理昼夜温度设为35 ℃/30 ℃, 其余培养条件跟预培养相同。试验完全随机设计, 每个处理设4次重复(4盆)。隔天浇水一次以保持土壤湿润。在高温胁迫处理后的第4、12、24 h和第5、15天取倒数第3~5片功能叶片, 液氮速冻后保存于-80 ℃冰箱用于基因表达分析; 在高温处理后的第5 和15天进行冠层高度、叶片电导率、叶绿素荧光和光合作用测定, 同时取倒数第3~5片功能叶片, 液氮速冻后保存于-80 ℃冰箱用于丙二醛和碳水化合物测定分析。
1)冠层高度:在高温处理前和高温处理的第5和15天用直尺测定各盆栽材料的冠层高度(cm)。
2)细胞膜稳定性:用电导率表示, 根据Hu等[19]的方法进行测定。
3)丙二醛含量(MDA):采用pH 7.8的磷酸缓冲液提取, 然后根据Hu等[19]的方法用分光光度计(UV7500, 上海)进行测定。
4)叶绿素含量:叶片经二甲基亚砜在室温避光条件下浸提后采用分光光度计法进行测定[20]。
5)叶绿素荧光:叶片经暗适应30 min后, 采用德国WALZ公司生产的PAM 2500叶绿素仪进行最大光化学效率(Fv /Fm)、光化学猝灭系数(qP)、非光化学淬灭系数(NPQ)、光合电子传递速率(ETR)、PSⅡ 实际光化学效率(Φ PSⅡ )等叶绿素荧光参数的测定。
6)光合作用:参考Hu 等[21]的方法, 在上午9: 00-11: 00, 采用Li-6400光合仪直接测定叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)和CO2响应曲线(Pn-CO2); 测定光合作用时设定光强为600 μ mol· m-2· s-1, CO2浓度为400 μ mol· mol-1, 温度为25 ℃; 用直线回归法求得Pn-CO2响应曲线的初始斜率, 即为CO2羧化效率(CE), 根据Pn-Ci曲线计算气孔限制度(Ls):Ls=100%× (Aca- Aci)/Aca(ca 为大气CO2浓度), 同时用Photosynthesis Assistant软件计算最大光合速率(Amax)、CO2补偿点(CP)、Rubisco最大羧化速率(Vcmax)和RuBP再生速率(Jmax)。
7)碳水化合物测定:样品烘干后研磨过0.425 mm筛, 用92%的乙醇提取后按王正文[22]的方法用HPLC测定果糖、葡萄糖、蔗糖和淀粉的含量。
8)基因表达分析:总RNA采用Trizol试剂盒参考说明书进行提取, 经Nanodrop 2000检测浓度和纯度以及用1%琼脂糖凝胶电泳检测完整性后, 根据Fermentas cDNA synthesis 试剂盒说明书进行反转录合成cDNA。参考TaKara 公司的SYBR System操作手册, 采用ABI7500实时荧光定量PCR仪进行PCR扩增。每个处理设3个重复。基因特异性引物序列参考表1进行合成[23], 以YT521-B基因为内参。根据得到的Ct值, 利用2-Δ Δ CT法, 分别计算目标基因在不同处理下的相对表达量。
| 表1 试验中所用基因及其引物序列 Table 1 Genes and primers used in the experiment |
采用SAS 软件进行数据统计分析, 然后采用Duncan’ s多重比较检测处理间的差异显著性(P< 0.05), 采用Sigmaplot 12.0作图。
由图1可以看出, 高温胁迫下黑麦草生长显著受到抑制, 冠层高度在高温处理的第5 和15天时比对照低, 分别为27%和38%。但是外源施用GABA显著缓解了高温胁迫对黑麦草生长的抑制, 在高温处理第5 和15天时施用GABA的处理冠层高度比未施GABA的处理分别了提高了16%和20%。
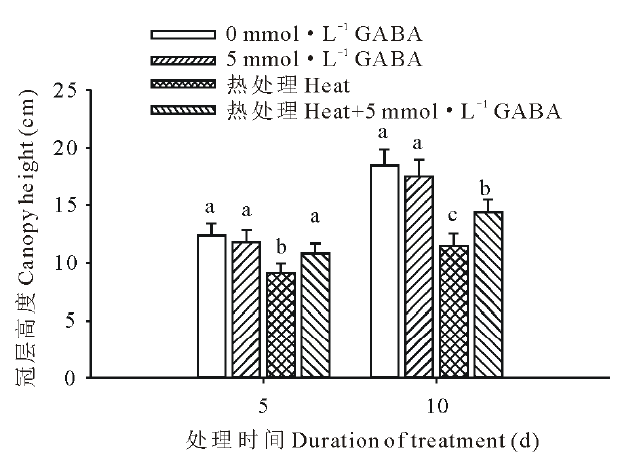 | 图1 外源GABA对高温胁迫下黑麦草冠层高度的影响 图中竖线表示平均值± 标准误, 不同小写字母表示在同一处理时间内不同处理间差异显著(P< 0.05), 下同。Fig.1 Effect of exogenous GABA on the canopy height in perennial ryegrass under heat stress Vertical bars indicate means± standard error (SE). Different lowercase letters indicate significant difference among treatment at a given day of treatment, the same below. |
由图2可知, 在高温胁迫下黑麦草叶片的相对电导率显著升高, 在胁迫的第5和15天时电导率分别提高了31%和78%; 但是高温胁迫下外源施用GABA后, 黑麦草叶片电导率显著降低, 与高温胁迫下未施用GABA的处理相比, 在高温处理的第5和15 天分别下降了30%和28%。
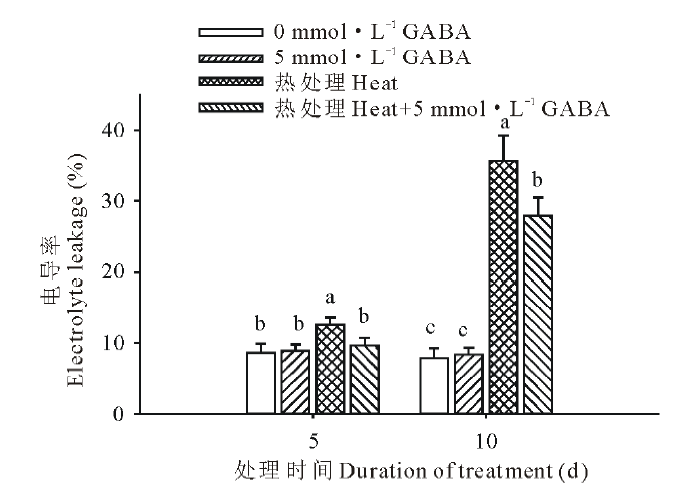 | 图2 外源GABA对高温胁迫下黑麦草叶片电导率的影响Fig.2 Effect of exogenous GABA on the leaf electrolyte leakage in perennial ryegrass under heat stress |
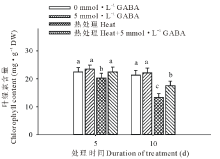
从图3可以看出, 高温胁迫下黑麦草叶片的叶绿素含量显著下降, 与对照相比, 在胁迫处理的第5和15天时的含量分别下降了9%和37%; 外源施用GABA后则显著缓解了因高温而造成的叶绿素含量的下降, 在高温处理后的第5和15天, 施用外源GABA的处理比未施GABA的处理叶绿素含量则分别提高了9%和23%。
 | 图3 外源GABA对高温胁迫下黑麦草叶绿素含量的影响Fig.3 Effect of exogenous GABA on the leaf chlorophyll content in perennial ryegrass under heat stress |
从表2可以看出, 正常温度下施用外源GABA短期内(5 d)提高了黑麦草的ETR, 但对Fv/Fm、Φ PSⅡ 、qP和NPQ无显著影响, 但较长时间(15 d)后对ETR、Fv/Fm、Φ PSⅡ 、qP和NPQ均无显著影响。短时间高温(5 d)对黑麦草叶片的Fv/Fm无显著影响, 较长时间高温(15 d)下黑麦草Fv/Fm显著下降; 高温胁迫下黑麦草叶片的ETR、qP和Φ PSⅡ 显著降低, 而NPQ显著升高; 但高温胁迫下施用外源GABA不论是在短期(5 d)还是较长时间(15 d)内均显著提高了黑麦草叶片的Fv/Fm、ETR、qP和Φ PSⅡ , 而降低了NPQ; 与高温下未施用GABA相比, 在处理5 d时, ETR、qP和Φ PSⅡ 分别提高了26%, 9%和13%, 而NPQ则降低了25%; 但到高温处理15 d, Fv/Fm、ETR、qP和Φ PSⅡ 则分别提高了17%, 50%, 21%和18%, 而NPQ则降低了32%。
| 表2 外源GABA对高温胁迫下黑麦草叶片叶绿素荧光参数的影响 Table 2 Effect of exogenous GABA on leaf chlorophyll fluorescence parameters in perennial ryegrass under heat stress |
由表3可知, 施用外源GABA对正常温度下黑麦草的Pn、Gs、Ci和Tr均无显著影响。高温胁迫下除了较长时间(15 d)处理下Ci外, 黑麦草叶片的Pn、Gs和Tr显著降低, 在处理5 d时, 分别降至对照的64%, 52%和67%, 到处理15 d时, 则分别仅为对照的35%, 37%和55%; 高温下黑麦草叶片的Ci在短期内(5 d)显著降低, 而较长时间(15 d)处理下则与对照没有显著性差异。高温胁迫下施用外源GABA显著提高了黑麦草叶片的Pn、Gs和Tr, 与未施GABA的处理相比, 在5 d时分别提高了31%, 45%和18%, 在处理15 d时则分别提高了73%, 45%和34%; 与高温胁迫下未施GABA的处理相比, 施用外源GABA在5 d时显著提高了黑麦草叶片的Ci, 而在处理15 d时则显著降低了叶片Ci。
| 表3 外源GABA对高温胁迫下黑麦草叶片光合作用的影响 Table 3 Effect of exogenous GABA on leaf photosynthesis in perennial ryegrass under heat stress |
由表4可知, 在短期(5 d)处理条件下, 高温胁迫和施用外源GABA对黑麦草叶片的CE、Vcmax和Jmax均无显著影响; 处理较长时间(15 d)时, 高温胁迫显著降低了黑麦草叶片的CE、Vcmax和Jmax, 但是施用外源GABA则显著缓解了高温条件下CE、Vcmax和Jmax的下降, 比未施用GABA的处理分别提高了70%, 31%和23%。高温胁迫显著提高了黑麦草叶片的Ls和CP, 降低了Amax, 但是施用外源GABA则显著缓解了因高温胁迫而造成Amax的下降以及Ls和CP的升高。
| 表4 外源GABA对高温胁迫下黑麦草叶片光合特性的影响 Table 4 Effect of exogenous GABA on leaf photosynthetic traits in perennial ryegrass under heat stress |
从表5可以看出, 在短期(5 d)处理条件下, 高温胁迫和施用外源GABA对黑麦草叶片的果糖含量无显著影响; 处理较长时间(15 d)时, 高温胁迫显著降低了黑麦草叶片的果糖含量, 但是施用外源GABA则显著缓解了高温条件下果糖含量的下降。高温胁迫处理显著降低了黑麦草叶片的葡萄糖、蔗糖和淀粉含量, 在高温处理5 d时, 分别降低至对照的76%, 79%和73%, 在处理15 d时则分别降低至对照的66%, 64%和53%; 高温处理下施用外源GABA显著提高了黑麦草叶片葡萄糖、蔗糖和淀粉的含量, 与未施GABA处理相比, 在处理5 d时, 分别提高了17%, 18%和19%, 在处理15 d时, 则分别提高了24%, 22%和40%。
| 表5 外源GABA对高温胁迫下黑麦草叶片碳水化合物含量的影响 Table 5 Effect of exogenous GABA on leaf carbohydrates content in perennial ryegrass under heat stress (mg· g-1 DW) |
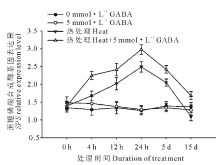
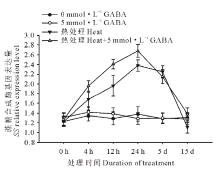
由图4和图5可知, 高温诱导了黑麦草叶片蔗糖磷酸合成酶基因(SPS)和蔗糖合成酶基因(SS)的上调表达, 并且随着处理时间的延长表达水平呈显著上升趋势, 在高温处理24 h时达到顶峰, 随后呈缓慢下降趋势, 在处理15 d时表达量降至对照水平; 在正常生长条件下, 外源施用GABA对SPS和SS基因的表达水平没有显著影响; 但在高温胁迫条件下, 与未施用GABA的处理相比, 外源施用GABA则显著诱导和提高了黑麦草叶片SPS和SS基因的表达水平, 在15 d的处理时间内均显著高于正常对照和高温下未施用GABA的处理。
 | 图4 外源GABA对高温胁迫下黑麦草叶片蔗糖磷酸合成酶基因表达的影响Fig.4 Effect of exogenous GABA on the expression level of SPS genes in leaves of perennial ryegrass under heat stress |
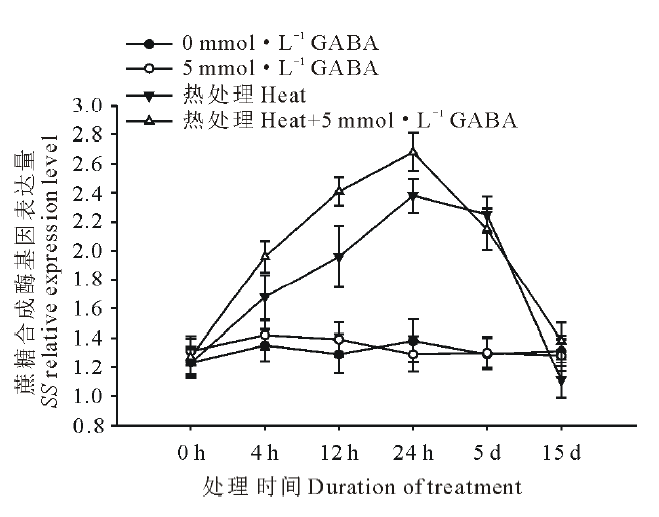 | 图5 外源GABA对高温胁迫下黑麦草叶片蔗糖合成酶基因表达的影响Fig.5 Effect of exogenous GABA on the expression level of SS genes in leaves of perennial ryegrass under heat stress |
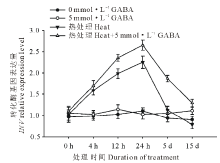
由图6可以看出, 高温胁迫诱导了黑麦草叶片转化酶基因(INV)的上调表达, 且随着处理时间的延长表达量上升, 在处理24 h到达顶峰, 随后下降, 在处理5 d以后则降至对照水平。在正常生长条件下, 外源施用GABA对INV基因的表达水平没有显著影响; 在高温胁迫条件下, 与未施用GABA的处理相比, 外源施用GABA则更显著地提高了黑麦草叶片INV基因的表达水平, 除了在处理4 h以外, 在15 d的处理时间内均显著高于正常对照和高温下未施用GABA的处理。
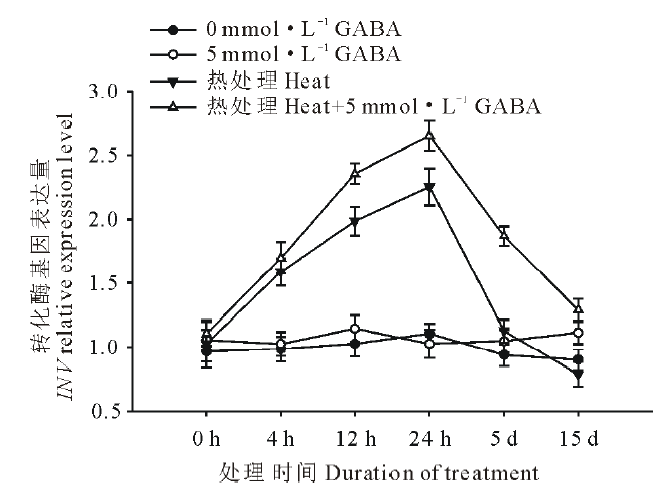 | 图6 外源GABA对高温胁迫下黑麦草叶片转化酶基因表达的影响Fig.6 Effect of exogenous GABA on the expression level of INV gene in leaves of perennial ryegrass under heat stress |
高温常造成冷季型草坪草生长发育受阻、草坪质量下降、细胞膜受伤害、叶片枯黄衰老等变化, 而植物的生长速率和电导率常作为衡量植物受高温胁迫损害程度和抗逆性大小的重要指标[24]。GABA可以作为外源物质参与到植物的生长发育以及逆境调控等一系列的生理过程和抗逆能力[25]。本试验结果显示, 高温胁迫下黑麦草的生长显著受到抑制, 叶片电导率升高, 但是施用外源GABA后黑麦草的冠层高度增加、细胞膜透性下降, 这表明外源GABA对黑麦草在高温胁迫下的生长具有促进作用, 施用外源GABA可以显著提高黑麦草的耐高温能力, 这与在冷季型草坪草匍匐剪股颖中的研究结果相一致[17]。
光合作用是植物对外界环境因子最敏感的生理过程之一, 在一定程度上可反映植株对逆境胁迫的抗性[3]。高温胁迫下植物叶片光合作用的降低既受气孔因素的限制, 同时也受非气孔因素的影响[4]。根据Farquhar等[26]观点, 气孔因素引起的Pn降低表现为Gs和Ci都降低, 而非气孔因素引起Pn降低则伴随着Gs的下降而Ci升高。本试验结果显示, 在高温胁迫5 d时, 黑麦草叶片的Pn随着Gs和Ci下降而降低, 且CE、Vcmax和Jmax无显著性变化, 但在高温胁迫15 d时Pn和Gs显著下降, 而Ci则没有下降甚至升高, 并且CE、Vcmax和Jmax均显著降低, 表明在短时间高温或轻度高温胁迫时, Pn的下降主要受气孔导度的下降等气孔因素的影响, 而在较长时间的高温胁迫或重度高温胁迫下光合作用的下降主要是受CE、Vcmax和Jmax等非气孔因素的限制, 这与前人的研究结果基本一致[27, 28]。已有研究证明外源喷施GABA能调节盐碱等逆境条件下植物叶片的光合性能[29]。本研究结果显示, 高温胁迫下施用外源GABA后黑麦草叶片的Pn显著提高, 缓解了高温对Gs和Tr的抑制, 减少了Ci的积累, 此外还显著提高了Amax、CE、Vcmax和Jmax, 表明外源GABA可通过缓解高温对叶片气孔和非气孔因素的限制, 从而改善了黑麦草叶片的光合特性, 提高了植株的耐热性, 这与黄娟等[30]在黄瓜上的研究结果一致。
叶绿素荧光参数对环境逆境胁迫反应敏感, 可反映环境因子对植物光合生理状况的影响[31]。因此, 对叶片叶绿素荧光参数的测量成为评价逆境胁迫对植物光合器官损伤的重要指标[32]。本试验结果显示, 高温胁迫下黑麦草叶片的Fv/Fm、qP、ETR和Φ PSⅡ 显著下降, 而NPQ显著升高, 表明在高温胁迫下黑麦草叶片的光系统Ⅱ 活性中心受到了伤害, 光能转换效率和电子传递能力下降, 光能利用率降低, 光能过剩加剧了对叶片光合的抑制和光合器官的破坏, 从而造成了叶绿素荧光参数的下降, 这与在冷季型草坪草高羊茅(Testuca arundinacea)中的研究相一致[27]。维持植物光合器官结构的稳定, 提高叶片光合能力对抵抗逆境胁迫至关重要。前人研究表明[11], 施用GABA可提高低氧胁迫下甜瓜叶片和盐胁迫下番茄叶片的光合能力。在本研究中, 外源施用GABA显著提高了高温胁迫下黑麦草叶片的Fv/Fm、qP、ETR和Φ PSⅡ , 降低了NPQ, 表明外源GABA可提高光系统Ⅱ 反应中心的光能转换效率、潜在活性和开放比例, 同时还增强了过剩光能的耗散, 有利于减轻高温胁迫对光合的抑制, 增强光合系统反应中心的稳定性, 从而减轻高温胁迫对黑麦草叶片光合器官造成的损伤, 提高了高温胁迫下黑麦草的光合能力和耐热性。
不同的植物种类对温度的敏感性和耐受性不同, 一定范围内的高温可以促进植物体内的生理生化反应, 从而促进光合和碳水化合物代谢, 而过高的温度便可降低光合作用, 增强了呼吸作用而消耗了更多的碳水化合物等能量物质使其含量显著降低[33]。逆境条件下可溶性糖(包括蔗糖、果糖、葡萄糖等)的增加能够增加细胞汁液浓度, 有利于细胞结构的稳定, 从而能够维持逆境下细胞内正常的生理代谢而提高抗逆性[34]。前期的研究结果表明[18], 高温胁迫下可溶性糖和淀粉含量显著下降, 并且其含量的高低与黑麦草的耐热能力密切相关。在马铃薯(Solanum tuberosum)中发现, 高温胁迫下其叶片蔗糖含量升高, 淀粉含量减少, 而葡萄糖含量无显著变化, 这可能与蔗糖合成酶(SS)和转化酶(INV)酶活性的提高有关[7, 35], 而在甘蔗(Saccharum officinarum)中却发现, 高温胁迫下其叶片的糖代谢关键酶(SS、SPS、INV)活性都降低, 蔗糖和淀粉含量减少[35]。本研究结果显示, 短期的高温胁迫下(5 d)黑麦草叶片的SPS、SS和INV酶基因的表达量显著升高, 而在较长时间(15 d)的高温胁迫下表达水平显著下降, 同时叶片的葡萄糖、果糖、蔗糖和淀粉等碳水化合物含量也显著下降, 表明高温对黑麦草的碳水化合物代谢造成了影响, 致使光合产物减少, 代谢关键酶基因表达量降低, 促进了呼吸作用使淀粉分解而含量下降。但是, 高温胁迫下施用外源GABA, 显著提高了黑麦草叶片SPS、SS和INV酶基因的表达水平和碳水化合物(蔗糖、果糖、葡萄糖、淀粉)含量, 表明外源GABA可通过缓解高温胁迫下光合作用的下降和提高碳水化合物代谢关键酶基因的表达水平和光合产物, 从而提高了碳水化合物代谢水平和增强了黑麦草的耐高温能力。
通过对高温条件下施用外源GABA对多年生黑麦草的生长、细胞膜透性、光合色素含量、光合及叶绿素荧光参数、碳水化合物含量及其代谢关键酶基因表达的分析表明, 高温胁迫下施用外源GABA, Fv/Fm、ETR、qP和Φ PSⅡ 提高了13%~50%, 而NPQ则降低了25%~32%, CE、Vcmax和Jmax分别提高了70%, 31%和23%, 降低了叶片气孔和非气孔限制值, 增强了糖代谢关键酶基因表达, 碳水合化合物含量提高了17%~40%, 缓解了高温对叶片PSⅡ 反应中心的抑制, 增强了光合能力, 提高了碳水化合物含量, 因而有效地提高了黑麦草的耐高温能力。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|