


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
高原鼠兔干扰对高寒草甸植物群落多样性和稳定性的影响
[徐海鹏 , 于成, 舒朝成, 金少红, 庞晓攀, 郭正刚
, 于成, 舒朝成, 金少红, 庞晓攀, 郭正刚* ]
, 于成, 舒朝成, 金少红, 庞晓攀, 郭正刚]
|
|


作者简介:徐海鹏(1994-),男,甘肃定西人,在读硕士。E-mail: xuhp17@lzu.edu.cn
物种多样性和功能冗余是评价植物群落稳定性的重要内容。采用野外调查方法研究了高原鼠兔干扰对高寒草甸植物群落物种多样性、功能多样性、功能冗余和植物群落稳定性的影响。结果表明:高原鼠兔干扰增加了物种丰富度指数、功能多样性和功能冗余,降低了物种多样性指数、均匀度指数和植物群落稳定性。随高原鼠兔干扰强度增加,植物群落多样性指数、丰富度指数、功能冗余和植物群落稳定性呈先增大后减小的变化趋势,说明高原鼠兔干扰对植物群落物种多样性和稳定性的影响不仅要考虑干扰与否,而且要考虑干扰强度。
Plant community diversity and functional redundancy are useful vegetation characteristics to consider when evaluating the community stability. A field survey was carried out to determine the disturbance effects of plateau pika on plant community diversity, functional diversity, functional redundancy and community stability of an alpine meadow. It was found that plateau pika disturbance increased plant species richness, functional diversity and functional redundancy, and decreased the plant diversity index, evenness index and community stability. In disturbed areas, plant species diversity, plant species richness, functional redundancy and community stability initially increased and then later decreased with increasing the plateau pika disturbance intensity. These results suggest that research into the effect of plateau pika disturbance on the plant community diversity and stability should consider not only the disturbance, but also the intensity of the disturbance.
稳定性是植物群落结构与功能的一个综合特征, 包括抵抗力稳定性和恢复力稳定性等[1, 2]。植物群落稳定性对草地生态系统可持续性具有重要的影响[3], 被视为评价生态系统健康的重要指标之一。虽然评价植物群落稳定性方法众多, 但物种多样性被认为是决定性因素之一[2, 3]。目前关于植物多样性与稳定性关系的假说尚未形成共识, 主要包括正相关、负相关和不相关3种模式[1], 但是Ives等[4]系统地对52个研究结果进行分析, 发现有69%的研究报道了多样性与稳定性呈正相关模式。植物多样性与稳定性呈正相关关系始于Elton[5]提出的生态系统越简单越不稳定, 主要是因为物种多样性越高, 生态系统抵抗外界干扰因子能力越强, 植物群落则越稳定[6]。同时物种冗余比多样性更能解释植物群落稳定性[7, 8], 因为冗余种的缺失不会影响植物群落稳定性, 但影响物种多样性, 主要是不同物种在植物群落中的作用和地位不同所致。然而, 植物群落内物种的地位和作用经常随着外界干扰而发生变化, 某一组分过度扩展或萎缩必然联动影响其他组分的变化, 一定程度上能够改变植物多样性, 引起冗余种变化或改变冗余种位置, 从而对植物群落稳定性产生影响。因此外界干扰, 特别是生物干扰, 对草地植物群落稳定性的影响成为生态学和草学研究领域的重要内容[9, 10]。
青藏高原生态环境脆弱[11], 高寒草甸对生物因子的干扰更加敏感[12], 因此青藏高原高寒草甸是研究生物因子干扰与植物群落稳定性关系的最佳场所。高山嵩草(Kobresia pygmaea)草甸是青藏高原高寒草甸的重要组成, 也是广布的草甸类型之一, 因其草层低矮而成为了高原鼠兔的主要栖息地, 恰巧也是高原鼠兔极易致灾的草甸类型[13, 14]。高原鼠兔(Ochotona curzoniae)是青藏高原地区特有的小型哺乳类动物[15], 也是该地区高寒草甸生态系统的关键种[16], 其通过采食、掘洞、覆土和排泄等活动对高寒草甸产生广泛影响[13, 15, 16]。高原鼠兔与高山嵩草草甸间在长期演化中形成了极其微妙的关系, 因此可以利用高原鼠兔干扰及其不同干扰程度下植物群落稳定性和植物多样性的变化特征, 阐明生物因子干扰与植物群落稳定性间的关系。
高原鼠兔干扰对高寒草甸具有正面和负面的双重影响[15, 17, 18]。已有研究表明, 适宜高原鼠兔干扰强度能够提高植物物种多样性[16], 促进降水入渗, 增加土壤养分含量[16, 19]。然而高原鼠兔干扰过大时, 则会降低土壤养分含量[20], 减少可食牧草比例[15], 降低原优势植物生态位宽度[21], 改变植物种群间的联结性[18]。这证明了高原鼠兔与高山嵩草草甸之间形成既互惠、又拮抗的关系, 而这种关系的转变取决于高原鼠兔的干扰强度[15]。然而高原鼠兔干扰对高寒草甸植物群落稳定性的影响尚需深入研究。因此本研究以高山嵩草草甸为例, 通过分析高原鼠兔干扰及干扰强度下高寒草甸植被物种多样性、功能多样性、功能冗余以及稳定性变化及其多样性与稳定性的关系, 揭示小型草食动物干扰对物种多样性和植物群落稳定性耦合关系的影响, 为全面认知青藏高原地区高原鼠兔的作用和防控提供科学理论依据。
研究区位于青藏高原东缘的甘肃省玛曲县境内(100° 40'-102° 29' E, 33° 06'-34° 30' N), 海拔3300~4806 m。气候严寒, 主要是高原湿润气候, 年均温度1月温度最低(平均 -10 ℃), 7月温度最高(平均11.7 ℃)。年均降水量约为564 mm, 主要集中在5-9月, 年均蒸发量1000~1500 mm, 全年日照时数约2614 h, 年内霜期大于270 d, 无绝对无霜期。该区域以高寒草甸为主, 优势种为高山嵩草, 主要伴生种为垂穗披碱草(Elymus nutans)、小花草玉梅(Anemone rivularis)、钝裂银莲花(Anemone obtusiloba)、莓叶委陵菜(Potentilla fragarioides)、鹅绒委陵菜(Potentilla anserina)和长毛风毛菊(Saussurea hieracioides)等[20]。全县大约30%的草地鼠害较为严重, 高原鼠兔是最主要的危害鼠种, 其中重度危害区的有效洞口数约为1400个· hm-2, 严重影响了高寒草甸生态系统的稳定和健康[13, 22]。
野外调查于2016年7月在牧草生长旺盛时期进行, 该时期是高原鼠兔干扰最严重的时期[17]。调查在已围栏的冷季牧场(暖季不放牧)进行, 排除其他动物干扰的影响, 选择地势相对平坦且一致的高山嵩草草甸为取样区。首先选择高原鼠兔干扰样地, 以发现高原鼠兔出没和高原鼠兔洞穴为准, 样地面积25 m× 25 m(625 m2), 然后在干扰样地附近, 寻找没有高原鼠兔干扰的样地, 以没有高原鼠兔洞口(或很少, 指1 hm2内洞口密度不超过1个)和没有发现高原鼠兔出没为准, 设置未干扰样地, 样地面积和干扰样地等同, 同时保证干扰样地与非干扰样地的草甸类型和地形地貌保持一致[20, 23]。高原鼠兔具有领域性, 镶嵌分布于草甸[13, 23], 其巢域面积(亦称活动范围)大约为半径20 m的圆[24]。为避免高原鼠兔移动性对非干扰样地的影响, 非干扰样地在保持草甸类型和地形地貌一致的基础上, 尽可能远离干扰样地, 因此干扰和未干扰样地之间的距离在500 m之外。鉴于高原鼠兔镶嵌分布的特征, 干扰样地之间的距离介于13 km, 研究区域内干扰样地和非干扰样地整体呈交错分布状态, 共计设置10个干扰样地和非干扰样地。测定每个干扰样地的有效洞口数量, 以反映高原鼠兔干扰强度, 测定方法为连续3 d堵洞法[15, 20, 25, 26], 10个干扰样地有效洞口数分别为304, 384, 432, 512, 576, 752, 864, 928, 1040, 1216个· hm-2。
每个样地内(干扰和未干扰样地), 两条对角线交汇处设置一个样方, 然后沿任意一条对角线, 从交汇处向外延伸10 m, 再分别设置2个样方, 样方面积为1 m× 1 m, 记录每个样方内的物种数, 测定样方内所有植物的高度、盖度、多度和频度, 随后观察并记录样方中各植物的功能性状。
1.3.1 物种多样性的计算 物种多样性测度采用多样性指数、丰富度指数和均匀度指数, 包括Shannon-Wiener多样性指数(H)[23], 其计算公式:
H=-∑ PilnPi
式中:Pi是种的相对重要值(重要值由盖度、高度和频度计算)。
Patrick丰富度指数(S), S=样方内出现的物种总数
Pielou均匀度指数(J), 其计算公式:
J=H/ln S
1.3.2 功能多样性(Q)的计算 以物种的7个定性性状(科、生活型、叶表面类型、传粉方式、固氮类型、繁殖方式和花期时长)和3个定量性状(株高、物种多度、物种盖度)测定物种的功能特征类型(表1), 采用Rao’ s指数(Q)计算样方的功能多样性, 功能特征值数据的标准化采用Gower’ s距离公式[27]。计算公式如下:
Q=
式中:S表示样方内的物种数; pi和pj分别表示第i和j个物种占总物种个体数的比例; dij表示Gower’ s距离, 即物种i, j在一组性状空间中的相异性, 其数值介于0到1之间, 当两物种具有完全相同的特征时, 其数值为0, 当两物种具有完全不同的特征时, 其数值为1。
| 表1 植物功能特性类型 Table 1 Types of plant functional traits |
1.3.3 功能冗余(functional redundancy, FR)的计算 采用Bello等[29]的公式计算功能冗余:
FR=D-Q
式中:D为Simpson指数, D=1-
1.3.4 群落稳定性计算 植物群落稳定性采用改进后的Godron稳定性计算方法[30]。首先把各样地中不同种植物的频度由大到小的顺序排列并计算相对频度, 按相对频度由大到小的顺序逐步累加起来; 然后将样地内植物种类的总和取倒数并按植物种类的顺序逐步累积; 再将植物种类倒数百分率同累积相对频度一一对应, 画出散点图建立二次曲线模拟模型y=ax2+bx+c。最后再与直线y=100-x相交, 位于第一象限的交点即为所求群落的稳定点。一般地, 交点坐标距离(20, 80)越小, 群落就越稳定。
采用独立样本T检验分析高原鼠兔干扰对测定指标影响的差异性是否显著, 若差异不显著, 则不做进一步分析, 若差异显著(P< 0.05), 则对高原鼠兔不同干扰强度下各指标的变化进行线性拟合分析。利用Pearson相关法分析高原鼠兔干扰下植物群落多样性与稳定性的关系, 采用R软件进行数据分析。
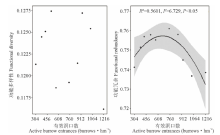
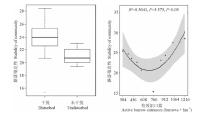
高原鼠兔干扰对植物物种多样性指数、均匀度指数和丰富度指数的影响不一致(图1), 表现为高原鼠兔干扰显著增加了物种丰富度指数(F=3.989, df=18, P=0.030), 但显著降低了物种多样性指数(F=0.908, df=18, P=0.018)和均匀度指数(F=10.029, df=18, P=0.048)。随着高原鼠兔干扰强度增加, 物种多样性指数和丰富度指数变化趋势相一致, 均呈先增加后降低的变化趋势, 但均匀度指数没有明显的变化规律(图2)。
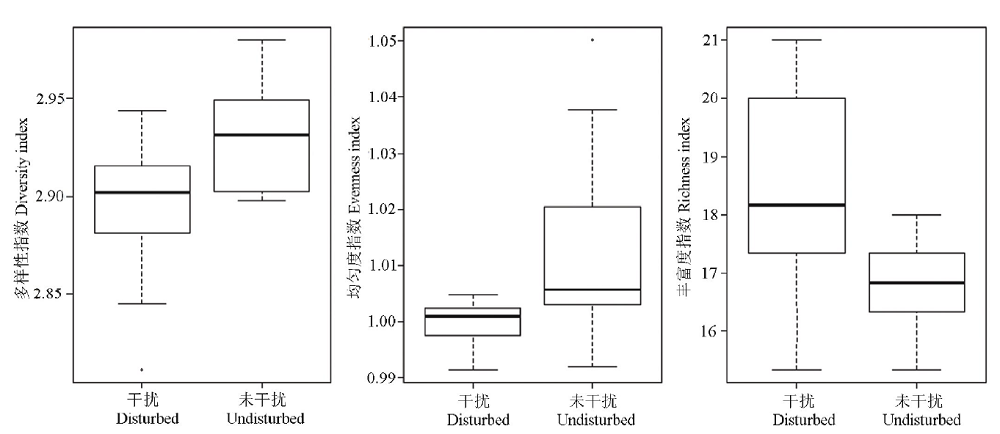 | 图1 高原鼠兔干扰对高寒草甸物种多样性的影响Fig.1 Effect of plateau pika disturbance on species diversity in alpine meadow |
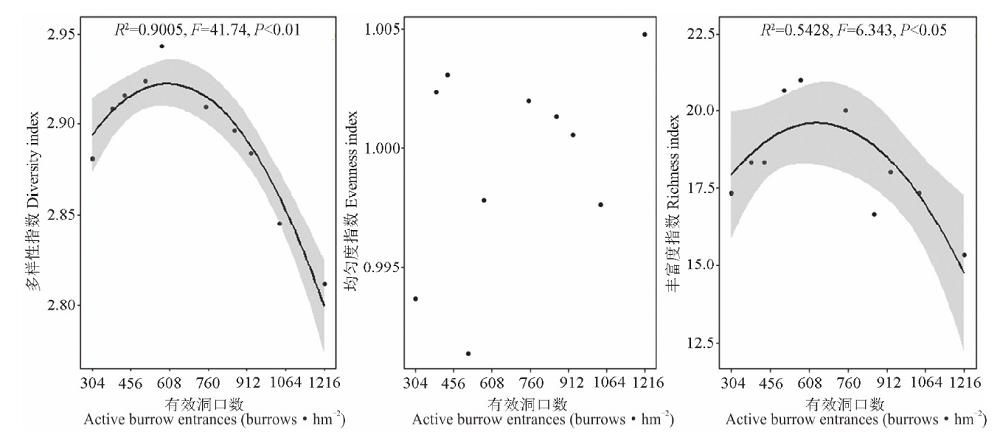 | 图2 高原鼠兔干扰区不同干扰强度下物种多样性的变化Fig.2 The species diversity under different disturbance intensities of plateau pika |
高原鼠兔干扰对植物群落功能多样性(F=0.000, df=18, P=0.004)和功能冗余(F=3.768, df=18, P=0.025)具有显著的影响, 表现为高原鼠兔干扰显著提高了植物群落的功能多样性和功能冗余(图3)。随着高原鼠兔干扰强度的增加, 植物群落功能多样性和功能冗余变化过程并不一致, 其中功能冗余表现出单峰变化趋势, 且在中度干扰下植物群落功能冗余最大(图4), 而植物群落功能多样性无明显的规律。
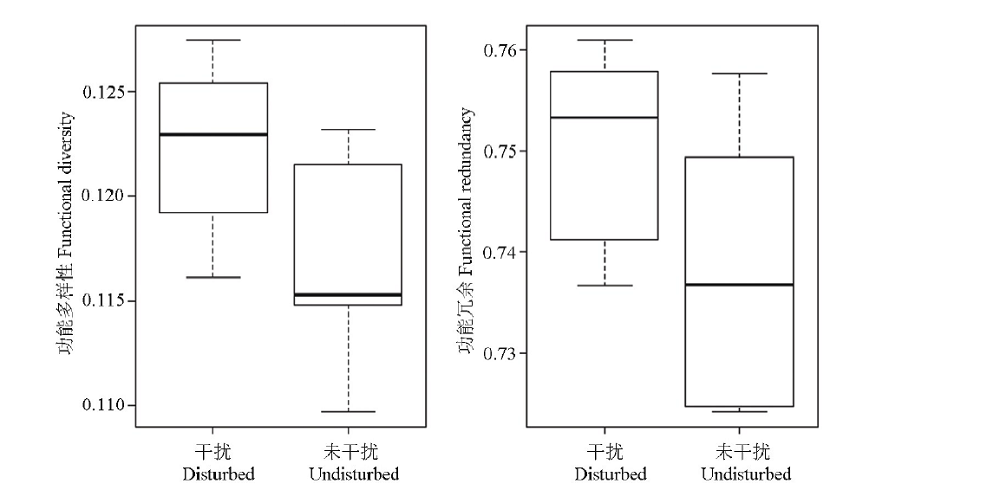 | 图3 高原鼠兔干扰对高寒草甸植物群落功能多样性和功能冗余的影响Fig.3 Effect of plateau pika disturbance on functional diversity and functional redundancy in alpine meadow |
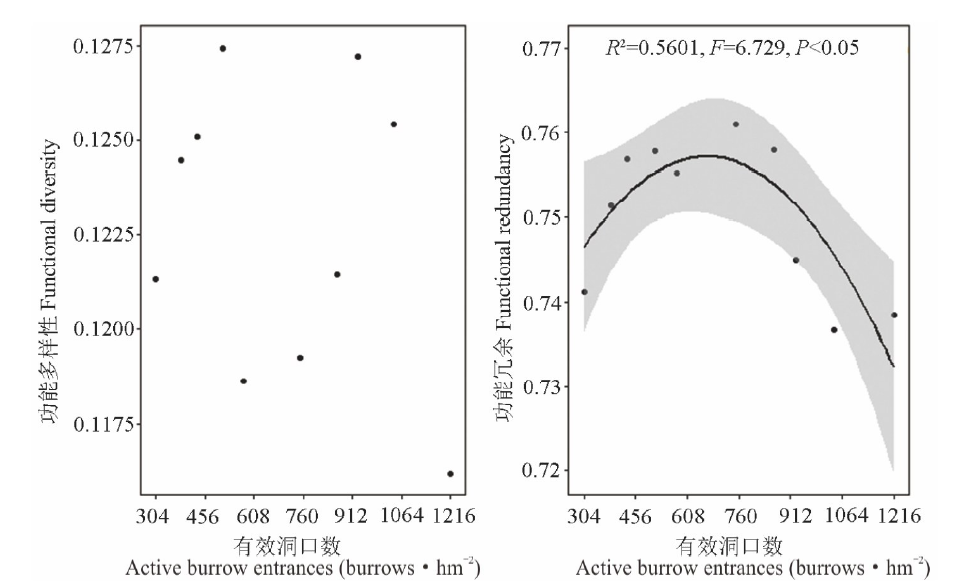 | 图4 高原鼠兔干扰区不同干扰强度下功能多样性和功能冗余的变化Fig.4 The functional diversity and functional redundancy under different disturbance intensities of plateau pika |
Godron稳定性计算结果表明:高原鼠兔干扰显著影响了植物群落的稳定性(F=3.393, df=18, P=0.046), 表现为干扰样地距离稳定点的距离较远, 说明群落不稳定。随着干扰强度增加, 各样地距离稳定点的距离表现为先降低后增加(图5), 即中度干扰时植物群落稳定性距离稳定点的距离最近, 稳定性最好。
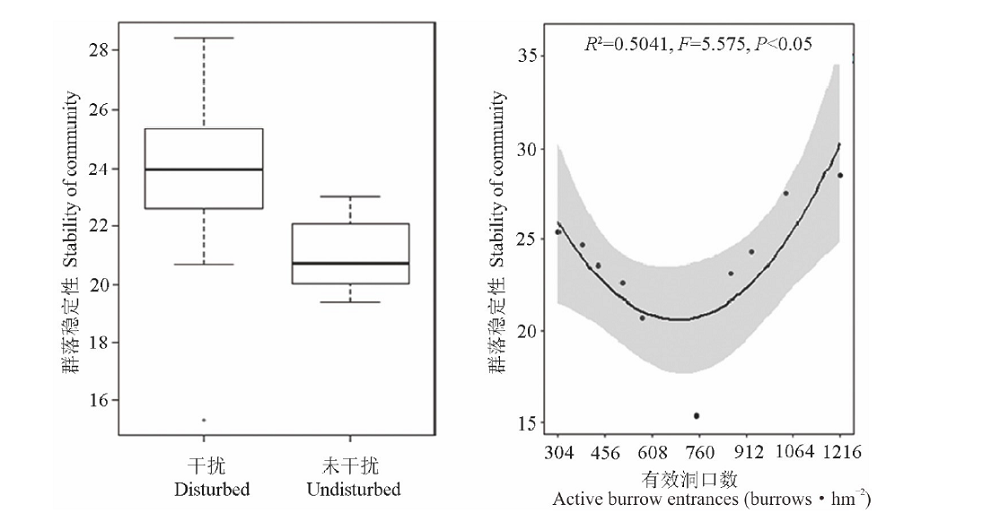 | 图5 高原鼠兔干扰及干扰区内不同干扰强度下植物群落稳定性的变化Fig.5 The effect of plateau pika disturbance on community stability and its variation under different disturbance intensities |
高原鼠兔干扰下, 植物群落稳定性与物种丰富度指数和功能冗余呈显著正相关(P< 0.05)(表2), 即植物群落稳定性随着物种丰富度指数和功能冗余的增大而增大。植物群落物种多样性指数、均匀度指数和功能多样性与植物群落稳定性之间无明显的相关关系。从物种多样性内部分析, 功能冗余与多样性指数和丰富度指数呈现显著正相关(P< 0.05), 丰富度指数与多样性指数之间呈显著的正相关(P< 0.01)。
| 表2 高原鼠兔干扰下植物群落多样性与稳定性的相关关系 Table 2 Correlation coefficients of species diversity and community stability under plateau pika disturbance |
高原鼠兔作为青藏高原高寒草甸生态系统内特有的群居性小型哺乳类动物[15, 16], 通过掘洞、采食、排泄和覆土等行为直接或间接影响植物群落结构和功能[31], 且影响效应随高原鼠兔干扰强度不同而存在差异[15, 26]。本研究表明, 高原鼠兔干扰增加了高寒草甸植物群落的物种丰富度指数, 但降低了物种多样性指数, 这既与阿根廷荒漠草原上栉鼠(Ctenomys mendocinus)干扰对植物多样性和丰富度的影响趋同[32], 又与高原鼠兔对青海治多县和甘肃碌曲高寒草甸植物多样性的影响类似[26], 主要原因是高原鼠兔干扰增加了高山嵩草草甸生境的异质性, 为不同物种共存提供了条件, 从而增加了植物丰富度指数[33]; 其次, 高原鼠兔通过携带行为和粪便将植物种子分散到大量裸斑, 这些种子当草毡层存在时, 发芽和定居较难, 而其散落于裸斑, 则增加了土壤种子库[34, 35], 且容易定居, 从而增加了植物丰富度指数。然而高原鼠兔的干扰降低了优势种重要值, 削弱了其在植物群落内的地位和作用[21]。物种均匀度指数是多样性指数的另一个基本组分, 反映了植物群落或生境中各物种个体分配的均匀程度[36]。本研究发现高原鼠兔干扰后物种均匀度指数下降, 这与放牧对美国落基山脉地区灌木草原群落均匀度影响一致[37], 主要是高寒草甸被干扰后, 土壤含水量和养分的变化使部分物种的生长状态更加弱势, 而部分物种生长潜势得到强化, 从而引起均匀度指数的降低。在高原鼠兔干扰区不同干扰强度下, 物种多样性指数和丰富度指数均随着干扰强度的增加呈现先增高后降低的趋势, 这与Bagchi等[38]在高山跨喜马拉雅地区干旱牧场上的Voles(Alticola stoliczkanus)和高原鼠兔干扰的研究结果基本一致。Biswa等[39]对加拿大Ontario西北部森林人为干扰下的河岸生境植物群落中的研究, 也证实了在中度干扰下植物群落的多样性指数和丰富度指数最大。说明小型草原哺乳动物在中等干扰梯度时能够增加植物物种丰富度指数和多样性指数。
功能多样性是联系物种多样性和生态系统功能的关键性因素[40], 其对生态系统功能变化具有决定性作用[41], Cadotte等[42]研究发现植物功能性状多样性对扰动和环境胁迫更敏感, 比物种多样性更能揭示生态系统功能的变化, 本研究发现, 高原鼠兔干扰增加了植物功能多样性, 原因可能是高原鼠兔干扰下植物群落中的生态位空间分化度增加, 因此虽然被占据, 但并没有被完全利用[21]。功能冗余是生物群落抗御性的关键属性, 对生态系统过程的维持具有保险作用[8]。高原鼠兔干扰增加物种功能冗余, 说明植物群落在响应高原鼠兔干扰过程中, 物种间类似或相同功能的程度增加[43], 其综合反映的结果就是维持植物群落受到高原鼠兔干扰后的稳定性[44]。高寒草甸群落的稳定性不仅与生物多样性有关, 还与干扰类型和干扰程度存在着紧密关系[45], 本研究表明高原鼠兔的干扰影响高山嵩草草甸植物群落的稳定性, 可能是由于高原鼠兔的干扰改变了植物群落的物种组分[23], 也可能促进或抑制了某些植物的生长, 改变了物种在原有植物群落中的地位和作用, 导致植物群落稳定性降低。然而, 当高原鼠兔干扰程度从轻度增加到中度时, 物种丰富度和功能冗余增加, 植物群落稳定性逐渐增加, 但干扰梯度从中度增加至重度时, 植物群落稳定性降低, 这是因为高干扰强度下植物群落内的物种多数为杂类草[21], 而杂草较优势种高山嵩草更容易退化, 整体导致植物群落稳定性降低。因此高原鼠兔干扰对植物群落稳定性的影响不能仅仅采用干扰与否评价, 更应该考虑干扰强度的影响, 才能全面揭示高原鼠兔干扰对植物群落稳定性的影响。
植物多样性与群落稳定性关系的问题在科学界存在着长期的争论。许多研究指出物种多样性与物种的功能多样性和功能冗余正相关[29], 对植物群落的稳定性具有显著正效应[8, 46, 47]。本研究得出, 高原鼠兔干扰下群落的稳定性与物种丰富度指数和功能冗余呈现正相关, Pillar等[8]在研究巴西南部的放牧草地中也发现了类似的结果, 其研究表明了草地植物群落稳定性的维持仅依赖于功能冗余的保险效应, 而不受功能多样性的影响。功能冗余与物种多样性指数和丰富度指数显著正相关, 这一定程度上支持了Naeem[44]的研究结果, 即物种丰富度越高, 功能冗余就越大。Pillar等[8]对巴西南部放牧草地的研究还表明, 群落稳定性仅有功能冗余决定, 本研究发现群落稳定性与物种丰富度指数也呈现正相关, 这主要取决于物种丰富度和功能冗余二者间的关系。Pillar等[8]是基于一个长期放牧草地的研究, 长期的放牧压力可能使得草地植物产生内在冗余, 进而使功能冗余成为维持植物群落稳定性的主要因子。因此, 利用物种丰富度指数和功能冗余说明高原鼠兔干扰下高山嵩草草甸稳定性的变化过程更为科学。
综上所述, 高原鼠兔干扰能够增加高山嵩草草甸植物群落的丰富度指数、功能多样性指数和功能冗余, 降低了群落的稳定性。但这种有益作用又与高原鼠兔干扰梯度密切相关, 只有中等干扰程度下才能增加群落稳定性, 重度干扰下植物群落稳定性则会降低。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|