



{kind=link}
{kind=link}
{kind=link}
{kind=link}
植物沙障不同种植模式对川西北沙地的恢复效应
[苟小林1, 2  , 刘文辉
, 刘文辉1 , 陈有军1, 2 , 周蓉3 , 周青平2, * ]
, 刘文辉]
|
|


作者简介:苟小林(1988-),男,四川剑阁人,博士。E-mail: gouxlchina@163.com
生物沙障被广泛地应用于沙化治理中,但是生物沙障不同种植模式下对沙化治理的相关研究较少,尤其是复合草本在高寒地区不同种植模式下的研究较为缺乏。为了明确混合草本沙障不同种植模式对高寒沙地治理效应,选择川西北沙地作为典型高寒半湿润沙地,利用燕麦、垂穗披碱草、中华羊茅作为混合草本沙障,自2014年利用行播、带播、撒播种植模式恢复高寒半湿润沙地4年,于2014年测定一年生燕麦生长状况,调查4年后地表植物群落盖度和多样性特征,并且测定土壤水分、容重、含水量、pH值、有机碳、全氮、全磷等指标,分析复合草本不同种植模式对高寒沙地恢复效应。研究结果表明:复合草本带播和撒播模式对高寒沙地恢复效果明显。一年生燕麦后期从生态系统中消失,多年生草本适应性良好。带播和撒播种植模式显著增加了地上植物盖度、物种多样性、生态优势度。带播和撒播模式也显著减小了土壤容重,增加了土壤含水量、有机碳、全氮、全磷等含量。在高寒沙化复合草本种植恢复过程中,建议将土壤容重、有机质、全氮作为衡量土壤性质变化的指标。综合地表植被和土壤理化性质变化,建议以混合草本带播和撒播种植模式作为高寒沙化治理的主要种植模式。
Establishment of vegetation for sand stabilization is a widely used technique in restoration of desertified land, and little research has been conducted into the effects of different planting patterns on restoration outcomes. In particular, few studies have examined different planting patterns for composite grass-sand barriers for restoration in alpine-cold desert regions. In this study, typical alpine sub-humid sandy land in the northwest of Sichuan was planted in 2014 in three patterns (row planted, band planted, or scatter planted) with a mixture of three grasses: Avena sativa, Elymus nutans and Festuca sinensis. Data were collected in August 2014 on the growth of A. sativa, and again in August, 2017 to check plant community, coverage, biodiversity and collect soil samples. Soil water content, bulk density, pH value, organic carbon, total nitrogen, total phosphate, ammonia nitrogen, nitrate nitrogen and available phosphate were determined by standard laboratory techniques. Results showed that band and scatter planting patterns had significant positive effects on restoration of sub-humid sandy land. The annual species A. sativa had disappeared from the ecosystem by 2017, and the two perennial grasses grown well. The band and scatter planting patterns showed significantly superior plant coverage, biodiversity and ecosystem development, together with decreased soil bulk density, but increased soil water content, organic carbon, total nitrogen, total phosphate and other benefits. For sub-humid alpine sandy land restoration using grass mixtures, we suggest soil bulk density, organic carbon and total nitrogen are useful indexes of soil improvement. Also we recommend use of grass mixtures in band or scatter planting patterns as a suitable technique for restoration of desertified alpine land, to both restore vegetation cover above ground, and improve soil properties below ground.
沙化土地的治理一直是退化生态系统保护和恢复的重要内容, 特别是在我国沙漠化问题严峻的背景下, 沙化土地的防治成为关系国计民生的重大生态问题[1, 2]。在长期的沙化恢复研究中, 诸多学者提出了生物沙障防治、草灌方格防治、自然封育等措施, 并且取得了卓越的成效[3, 4, 5, 6]。如在科尔沁沙地使用了蒿属(Artemisia)植物和麦草秸秆方格进行沙化治理[4], 能够有效改善沙漠生境; 利用黄耆属(Hedysarum)等对低海拔干旱的腾格里沙漠进行治理[3], 对生态系统恢复具有正向的促进作用; 在高纬度的古尔班通古特沙漠利用草方格进行沙化防治, 有效地阻止了沙粒移动, 促进了沙面环境稳定[6]。生物沙障已经被广泛应用于沙化治理中, 但现阶段主要的应用模式是通过草灌方格进行沙化治理, 种植模式的研究较为缺乏[6]; 同时研究区域也主要针对低海拔地区的寒、旱沙漠和退化生态系统, 而高海拔寒冷地区相关研究较少[7, 8]。高海拔寒冷地区生态系统退化严重, 也面临着沙漠化急剧扩张等问题[7, 8, 9]。针对高寒沙化区域的沙漠化防治势在必行, 但现阶段沙化防治手段较为缺乏, 而利用生物沙障必然受到独特的环境条件的影响。如高寒地区海拔较高, 每年植物生长周期较短[10], 生物沙障的物种筛选需要适应该地区的短暂的生长周期, 并且需要考虑沙化防治的效果; 特别是筛选植物的种植模式可能受寒冷环境(漫长的冬季、雪被、频繁地冻融)的影响, 从而对植物的生长和繁殖产生影响[11, 12], 进而影响生态系统内植物更新和演替过程, 从而影响沙化防治。但高海拔寒冷地区生物沙障不同种植模式下沙化防治的研究极为缺乏, 对高寒地区沙化防治工作开展不利, 亟待研究。
青藏高原东南缘的川西北地区是典型的高海拔寒冷地区, 该地区是我国重要的高寒牧区, 同时也是长江和黄河的重要源头区域[13], 但是该区域生态环境退化严重, 部分地区沙漠化急剧扩张, 该区域的环境保护和沙化治理问题已经刻不容缓[14]。川西北沙化地区主要分布于半湿润气候条件下, 形成的沙地主要以固定和半固定的沙丘为主[15]。综合考虑川西北生境和本土植物分布, 发现可采用适生的灌木、中药材、牧草作为生物沙障进行沙化防治, 如该区域适生的披碱草属(Elymus)牧草, 大面积种植的燕麦(Avena sativa)[16]等; 但是利用这些草本作为沙化防治的相关研究极少, 特别是利用复合草本在不同种植模式下对川西北沙化地的适用性和对沙地的防治效果没有明确的结论。为了明确复合草本不同种植模式在川西北地区沙地上种植对沙地的防治效应, 本研究在前期工作的基础上, 以川西北高海拔寒冷沙地为研究基础, 分析燕麦、垂穗披碱草(Elymus nutans)和中华羊茅(Festuca sinensis)为复合草本不同种植模式下沙地地表植被变化情况, 土壤水分、容重、养分等变化情况, 探究复合草本不同种植模式对高寒沙地的防治效应, 为沙化防治提供基础研究。
研究区域位于四川省阿坝州红原县日干乔生态保护区(33.18° N, 102.62° E, 海拔3470 m)。日干乔生态保护区属于青藏高原东南缘, 为半湿润气候[15], 年降水量在750 mm左右, 2017年降水量为725 mm, 该区域处于高海拔寒冷区域, 近5年的年平均气温为2.55 ℃, 雨热集中。生长季节较短, 每年的5-9月为生长季节, 春、秋二季短暂, 冬季较为漫长, 具有分布不一的雪被覆盖, 土壤在经过频繁的冻融条件后进入和结束冬季过程[14, 15]。土壤表层主要为高寒草甸土和沼泽泥炭土, 沙化地区以风沙土为主[17]。该区域主要受到人为干扰(如过度放牧)的影响, 大量沼泽和泥炭地干涸, 土壤退化。该地区退化形成的沙地夏季温差极大, 太阳直射沙地表面温度可达50 ℃以上, 夜间温度可低至5 ℃左右(图1), 而冬季受地形因素影响, 具有大小不等的雪被覆盖, 冻融情况尤为明显。沙地周边地表植物主要以适应高寒环境的中国沙棘(Hippophae rhamnoides subsp. sinensis)、西藏沙棘(Hippophae thibetana)、奇花柳(Salix atopantha)、沙蒿(Artemisia desertorum)、鹅绒委陵菜(Potentilla anserina)、高山蒿草(Kobresia pygmaea)、四川蒿草(Kobresia setschwanensis)等为优势植物。
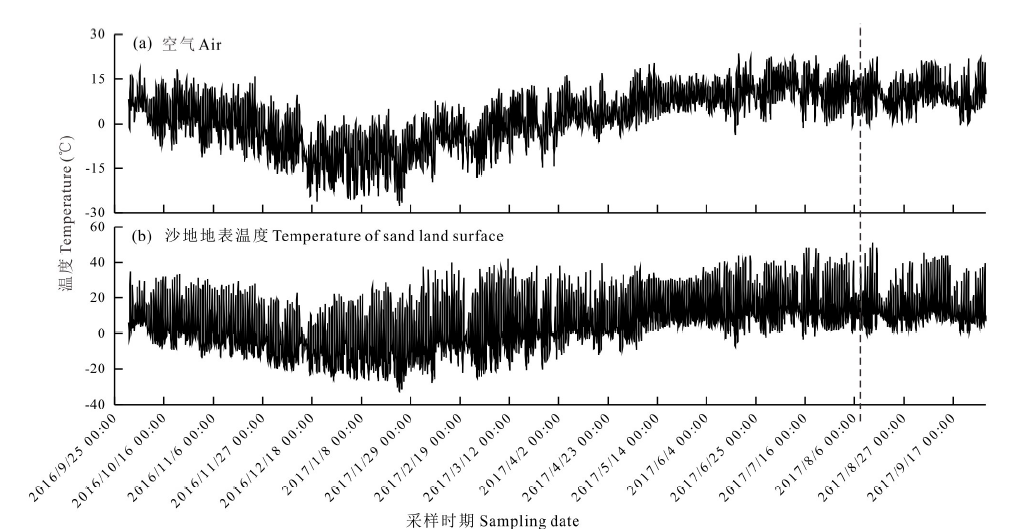 | 图1 研究区空气和沙地地表每2小时温度Fig.1 Temperature of air and sand land surface in every two hour |
于2014年4月末在研究区域选择地形相似的封育沙地, 按照不同的种植模式进行混合草种的种植, 并以相邻的空白沙地作为对照组。混合草种选用一年生的燕麦、多年生的垂穗披碱草和中华羊茅, 将选择的种子按照重量标准, 以燕麦:垂穗披碱草:中华羊茅=10:3:3均匀混合, 混合后的种子用于播种。为保障一年生草本的发芽和成活率, 播种量以燕麦大田播种量为基准[18], 保障种植面积内燕麦达到450万株· hm-2, 所以混合种子的播种量为160 kg· hm-2。行播:利用开沟器开10 cm宽的浅沟, 在浅沟内种植混合草种, 覆土5 cm, 10 cm宽的浅沟交错为50 cm× 50 cm 的草方格, 种植区域为0.5 km× 2 km。带播:在沙地上划分宽为1 m的条带, 条带之间间隔为1 m, 在条带内均匀撒播混合种子, 利用沙土覆盖, 覆土5 cm, 种植区域为1 km× 2 km。撒播:在1 km× 2 km的范围内进行混合种子的撒播, 撒播种子人工平耙, 使混合种子覆土在3~5 cm。根据沙化地实际分布情况, 每个处理有平坦的两块沙地(坡度2° ~4° )和一块缓丘(坡度11° ), 共计3个重复, 重复之间有柳树灌丛阻隔, 阻隔带在50~100 m之间。
于2014年8月在植物的生长旺季, 调查一年生燕麦的成活率和生长情况。在3种种植模式的重复样地中, 随机选择3块10 m× 10 m的样方, 每块样方之间相互水平间隔100 m以上, 调查燕麦的生长情况, 在每块10 m× 10 m 的样方中, 随机选择3块1 m× 1 m 的小样方, 在小样方中按照燕麦生长株数, 计算燕麦的成活率, 并随机选择10株燕麦调查燕麦的株高、抽穗情况。3块小样方的数据平均计算为一个重复, 每个种植模式3个重复。为了不破坏沙地的治理效果和后期的持续恢复, 所以不采集一年生的燕麦进行生物量、生物养分、土壤养分的分析。在治理4年后, 于2017年8月的生长旺季, 调查混合草本种植后沙地植物的分布情况, 调查种植后土壤理化性质和养分变化情况。在空白样地和3种种植模式的样地中, 分别随机选择3块10 m× 10 m 的样方, 在每个样方中随机选择3个2 m× 2 m 的小样方, 用于调查植物种类、株数、盖度、株高, 将3个小样方内的调查结果平均为一个重复; 由于一年生和多年生草本在4年的治理下, 植物根系和地表凋落物对土壤表层影响较大, 为了研究表层和相对较深的沙土性质变化, 在3个小样方内利用直径为5 cm 的土钻分别取种植地下0~10 cm和10~20 cm的沙土, 将3个小样方中的土壤混合为一个重复, 将土壤保存于自封袋中, 放入装有冰袋的泡沫盒; 并利用内径为5 cm, 容积为100 cm3的环刀, 分别取0~10 cm土壤剖面0~5 cm、3~8 cm、5~10 cm和10~20 cm土壤坡面10~15 cm、13~18 cm、15~20 cm的原状土壤用于土壤容重测定, 将所有样品带回实验室进行分析; 每个模式3个重复。
利用恢复4年后样方内调查的植物物种和分布情况, 对物种的丰富度指数(Species richness, R)、生态优势度指数(Simpson diversity index, D)、物种多样性指数(Shannon-Wiener index, H')、均匀度指数(Pielou index, E)进行分析, 比较不同恢复模式下植物多样性的变化情况。
群落物种丰富度指数[19]:R=S
式中:S为群落中的总物种数。
生态优势度指数(Simpson指数)[20]:
式中:N为群落中全部种的总个体数, Ni为第i种的个体数, S为群落中的总物种数。
物种多样性指数(Shannon-Wiener指数)[21]:
式中:Pi =Ni/N, Pi为群落中第i种个体数占群落中全部种总个体数的比例, S为群落中的总物种数。
均匀度指数(Pielou指数)[20]:E=H'/ln S
式中:H'为Shannon-Wiener指数, S为群落中的总物种数。
将采集回的土壤样品在室温下过孔径为1 mm筛, 去除沙土中的石子和草根。取部分土壤风干, 进行土壤样品pH、有机碳、全氮、全磷及有效磷含量测定, 新鲜土壤进行土壤水分、铵态氮、硝态氮的测定, 环刀土样测定土壤容重[22]。土壤水分利用105 ℃烘干至恒重进行测定[22]; 土壤硝态氮利用稀盐酸在220和275 nm的差值进行测定[22]; 铵态氮利用靛酚蓝方法进行测定[22]。土壤pH利用水电位法进行测定(PHS-3E), 土水比为1:1[22]; 土壤有机碳利用酸性K2Cr2O7加热消煮, 亚铁盐滴定测定[22]; 利用凯氏半微量法测定土壤全氮含量[22]; 利用HClO4-H2SO4在高温下消解风干土壤样品, 然后利用钼锑抗比色法进行测定土壤全磷含量[22]; 利用NH4F-HCl浸提风干土壤样品, 然后利用钼锑抗比色法进行测定土壤有效磷含量[22]。土壤容重利用环刀(内径5 cm, 容积100 cm3)在105 ℃下加热进行测定[22]。
采用单因素方差分析(One-way ANOVA)方法和最小显著差异法(LSD)分析一年生燕麦不同种植模式之间成活率、株高之间的差异, 分析不同种植模式之间植被盖度、丰富度、生态优势度、物种多样性、均匀度的差异, 分析每个土壤层不同种植模式之间土壤水分、pH、容重、有机碳、全氮、铵态氮、硝态氮、全磷、有效磷之间的差异, 显著水平为0.05。并利用Pearson相关分析不同种植模式下物种多样性和每个土壤层土壤性质之间的关系。利用主成分(PCA)分析方法分析各土壤层土壤指标的主成分效应。所有数据均采用SPSS 19.0分析, 利用OriginPro 9.0作图。
在不同的种植模式下, 复合草本中一年生燕麦的成活率并没有显著差异, 但带播和撒播的燕麦生长情况较好(表1)。行播、带播、撒播3种模式下的燕麦成活率都超过了50%, 并且没有显著差异, 而3种模式下带播和撒播种植的燕麦株高都超过了30 cm, 叶长势也优于行播。
| 表1 不同模式下燕麦和植物群落的变化特征 Table 1 Changing of oats and plant community under different models |
不同种植模式下植物物种多样性和盖度增加, 均匀度减少(表1, 图2)。3种播种模式下, 种植4年后植物群落盖度显著增加, 并且撒播模式盖度显著高于对照和其他种植模式, 达到84%。复合草本种植后的植物物种丰富度显著高于对照组, 并且撒播的物种丰富度最高, 达到9种; 生态优势度指数在3种播种模式之间没有显著差异, 但显著高于对照组; 物种多样性指数3种播种模式显著高于对照组, 并且撒播模式下显著高于其他模式; 而复合草种播种降低了植物群落均匀度, 并且带播和撒播显著低于对照组。播种4年后一年生燕麦已经退出生态系统, 原生赖草一直存在于生态系统中, 人工种植的垂穗披碱草和中华羊茅也在生态系统中生长良好。一年生的灰绿藜、菥蓂、播娘蒿等物种入侵了恢复的生态系统, 其他多年生的草本如早熟禾、以礼草等分布不一。
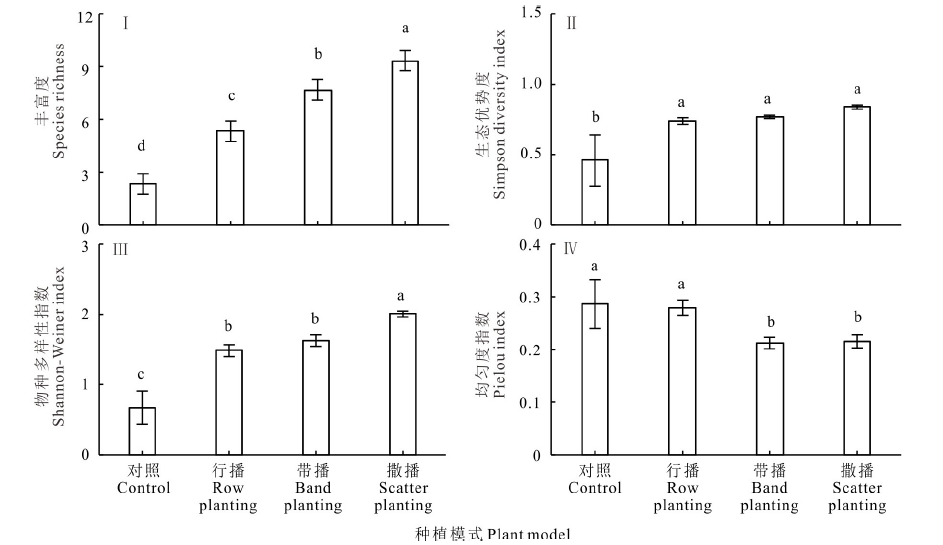 | 图2 不同种植模式下植物多样性指数 不同小写字母表示同一指标在不同种植模式下差异显著(P< 0.05)。Different lowercase indicate that the differences in the different plant models are significant at level 0.05Fig.2 Comparisons of plant diversity indexes under different plant models |
4年复合草本种植增加了土壤水分, 减少了土壤容重, 增加了土壤中养分含量(图3)。3种种植模式下, 行播明显减少了10~20 cm层的土壤水分, 撒播却显著增加了两个土层的土壤水分, 带播显著增加了10~20 cm层的土壤水分含量(图3Ⅰ )。4年的带播和撒播都明显减少了两个土层的土壤容重(图3Ⅱ )。行播和带播明显减小了土壤pH值(图3Ⅲ )。带播和撒播明显增加了两个土层中的有机碳含量, 行播却减少了土壤0~10 cm的有机碳含量(图3Ⅳ )。3种播种模式都增加了土壤中全氮含量, 带播和撒播增加明显, 撒播明显增加了土壤中硝态氮含量, 撒播和行播显著增加了土壤中铵态氮含量(图3Ⅴ , Ⅵ , Ⅶ )。带播和撒播明显增加了土壤中全磷含量, 而3种种植模式都显著增加了土壤中有效磷含量(图3Ⅷ , Ⅸ )。
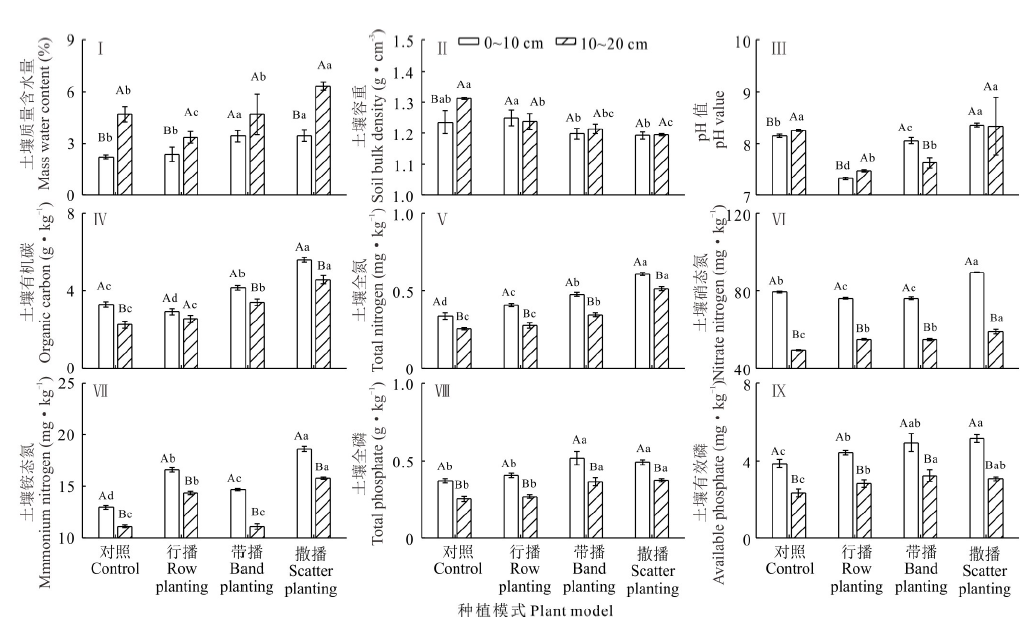 | 图3 不同种植模式下土壤理化性质和养分的比较 不同小写字母表示各指标在同一土层不同种植模式下差异显著(P< 0.05), 不同大写字母表示每种处理下各指标在不同土层之间差异显著(P< 0.05)。 Different lowercase indicate that the differences of the same soil layer in the different plant models are significant at level 0.05, different capital letters indicate that the differences are significant in different soil layers of every plant model at level 0.05 respectively.Fig.3 Comparisons of soil physical, chemical and nutrition under different plant models |
不同种植模式下, 0~10 cm土壤层的土壤水分、有效磷、全氮、全磷、铵态氮和植物群落盖度、丰富度、生态优势度、物种多样性、均匀度密切相关; 10~20 cm土壤层的容重、有效磷、有机碳、全氮、全磷、硝态氮和植物群落盖度、丰富度、生态优势度、物种均匀度密切相关(表2)。土壤中养分含量和物种丰富度均呈正相关关系, 土壤水分和物种丰度显著正相关, 土壤容重和丰度呈显著负相关, pH值和丰富度关系不显著。土壤pH值和容重都与物种生态优势度呈负相关关系, 但是只有10~20 cm土层显著, 其余都呈正相关关系。整个土壤层只有全氮、全磷、有效磷和物种多样性指数及群落盖度显著相关。整个土壤层的有机碳、全氮、全磷、有效磷、铵态氮和植物群落的物种多样性呈显著正相关。整个土壤层的有机碳、全氮、全磷、有效磷和群落均匀度显著负相关, 并且表层土壤水分和植物群落均匀度也呈极显著负相关。整个土壤层的水分含量、有机碳、全氮、全磷、有效磷、硝态氮和植物群落盖度显著正相关, 而整个土壤层的土壤容重和植物群落盖度显著负相关。
| 表2 土壤性质和植物群落之间的相关关系 Table 2 Pearson’ s correlations between soil properties and plant community |
不同种植模式下土壤各指标主成分分析后得分前3的为有机碳> 全氮> pH, 容重和其他土壤指标在第一主成分下变异较大(图4)。0~10 cm土壤层土壤性质各指标第一主成分达66.48%, 第二主成分达14.29%, 综合达 80.77%; 10~20 cm土壤层土壤性质各指标第一主成分为63.32%, 第二主成分为20.49%, 综合达83.81%。两个土壤层除容重外, 其余指标在第一主成分上的投影皆为正值, 其中前3的得分排序为有机碳> 全氮> pH。在两个土壤层中, 容重和其余指标在第一主成分上分布情况完全相反, 其他指标在第二成分分布差异较大, 特别是pH值在第二成分上的分布完全相反。对照组土壤各指标在第一主成分上的分布和3个处理组也明显不同。
 | 图4 不同种植模式下两个土壤层土壤成分的主成分分析Fig.4 Principal components analysis of soil properties in two soil layers under different plant models |
以一年生(燕麦)和多年生(垂穗披碱草、中华羊茅)混合草本对川西北高寒半湿润沙地进行沙化治理, 不同的种植模式并没有对燕麦的成活率产生明显的影响, 但是燕麦在当年生长后, 行播的燕麦长势(株高、叶等)劣于带播和撒播的燕麦(表1)。川西北地处高海拔, 生长季节主要为水、热集中的5-9月, 该时期日照时间较长, 并且太阳辐射较大[23]。在高寒沙地上生长的燕麦由于地表植被较少, 采用行播方式形成的燕麦区域单位面积密度小于带播和撒播的燕麦, 在受到生长季节强日照后, 地表温度极高, 而夜间温度较低(图1), 极大的昼夜温差对当年的燕麦生长造成了影响[24, 25], 而带播和撒播的燕麦单位面积内生长密度较大, 植株之间可以相互保护, 避免地表高温[26], 因此带播和撒播的燕麦长势要优于行播的燕麦。种植4年后, 一年生的燕麦退出了生态系统, 在不同种植模式中均未调查到燕麦, 燕麦在该生态系统中不能完成繁殖过程, 从该生态系统中消失。这可能是由于该区域的生长季节短暂, 同时燕麦需要忍受较大的环境变化(较大的温差、干旱、贫瘠等), 因此选用的燕麦品种在该区域没有完成繁殖过程, 不适应此生态环境, 逐渐退出了该生态系统, 人工选择的燕麦在生态系统植被演替中是一种前期物种[27]。但是燕麦在生长过程中, 特别是在种植初期, 当年的生长可能为多年生草本提供了保护效果, 如遮阴、地表水分保持等[28, 29], 并且死亡后地表形成的枯落物和残茬也可能对多年生草本幼苗安全度过冬季起到保护作用[30], 但是其机理仍然需要进行深入研究。
不同种植模式下, 经过4年的沙化治理, 植物群落盖度显著增加, 并且撒播种植模式下的植物群落盖度增加量最多。撒播模式下植物群落物种多样性和物种丰度最高, 并且增加的物种涵盖了菊科、黎科、十字花科、景天科等, 其中对水分要求较高的川西景天(景天科)只分布于撒播模式下, 因此撒播模式下水分条件可能优于其他模式。带播和撒播模式下物种均匀度分布明显小于对照和行播模式, 带播和撒播模式下入侵物种数目增加, 而增加了的各物种在短时期内可能还没有形成较多的个体分布, 导致了均匀度减少[31, 32], 因此带播和撒播种植模式下物种恢复情况要优于未治理和行播模式。未治理沙地上的优势物种赖草, 在不同种植模式下的生态系统中均生长良好, 并没有退出生态系统, 赖草对该区域沙化环境适应性良好。而选用的垂穗披碱草和中华羊茅都在3种模式中生长良好, 因此选用的多年生草本也适应该生态系统。一年生较为耐旱的短命植物菥蓂、灰绿藜等入侵恢复样地, 并且在3种模式下都广泛分布, 耐旱的短命植物对该生态系统有较好的适应能力, 这可能是由于短命植物在较短的生长周期内能够完成生命史, 并且在该时期内充分利用较为集中的降水[33, 34, 35]。
由于川西北高寒沙地土壤养分含量较低, 土壤贫瘠, 而利用一年生燕麦和多年生草本进行生态修复, 集中的浅层根系和地表凋落物主要影响表层土壤, 特别是在0~10 cm表土层。因此在分析过程中, 选择了0~10 cm和10~20 cm土壤层作为本研究对比分析对象, 能够基本反映土壤性质变化情况。在实际试验中, 不同种植模式4年的治理后, 带播和撒播显著增加了土壤层0~10 cm的含水量, 带播和撒播模式明显改善了土壤水分条件, 一方面, 川西北地区降水集中于生长季节, 该时期较高的地表植被盖度缓解了受到太阳直射引起大量水分蒸发[36, 37]; 另一方面, 地下密集的根系和死亡后的植物残体可能改善了土壤条件(如容重减小、有机质含量增加, 图3Ⅱ , Ⅳ ), 从而增加了土壤持水能力[38, 39, 40]; 并且川西景天在撒播模式下生长, 也侧面反映了撒播模式下较好的水分环境。经过4年的种植, 土壤容重减小, 土壤有机碳含量增加; 并且带播和撒播模式下效果显著, 在这2种模式下, 地表植被可能有较多的有机物反馈至土壤中, 增加的土壤有机质环境导致土壤容重减小, 土壤有机碳含量增加[41], 这些有机物主要来源于前期种植的燕麦死亡后地表凋落物归还, 地下根系分解等, 这可能得益于带播和撒播模式下较好的燕麦长势, 因此带播和撒播模式下对土壤有机碳的反馈能力高于行播模式。3种模式4年的治理下均显著增加了土壤中全氮含量, 而撒播模式下土壤铵态氮和硝态氮含量增加量最为显著, 因此撒播模式下对土壤氮含量的反馈能力最好。撒播和带播模式下均显著增加了土壤中全磷含量, 因此带播和撒播模式对土壤中磷的反馈能力也较好。由于土壤中的氮和磷含量相对有机碳含量较低, 并且较为稳定, 而在川西北地区沙地土壤中氮和磷含量远远低于未沙化的草地生态系统[14], 虽然带播和撒播模式明显改善了土壤中的氮和磷含量, 但是具体的机理还需进一步的深入研究。
不同种植模式4年的治理后, 土壤理化性质和地表群落特征密切相关, 地表植被群落恢复过程引导了地下土壤性质的相应变化, 植物群落盖度和多样性的变化影响着土壤水分, 容重和养分的变化过程[42, 43], 如地表植物盖度的增加, 使地表温度、水分等环境条件改善[36], 从而促进了地下养分循环和积累过程, 而地表植物年间的死亡和生长过程增加了土壤中养分的来源, 并促进了物质循环[44, 45], 因此土壤性质的变化也能够良好的反映生态系统恢复过程。在已测定的土壤性质中, 土壤容重和其他指标在第一主成分上象限分布不同, 不同种植模式下对土壤容重的作用和其他土壤性质的作用相反, 而其他指标得分前3的是有机碳、全氮、pH值。但是pH值和地表植物群落的相关性并不显著, 特别是在第二象限上, 两个土壤层中pH 值的分布差异极大, 而其他指标如有机碳、有效磷、全磷、含水量分布差异也很大, 这些分布差异的原因一方面可能是因为第二主成分的解释率太低, 在0~10 cm和10~20 cm 土层分别只有14.29%和20.49%, 所以各土壤指标在两个土壤层下的分布差异较大; 另一方面1~10 cm和10~20 cm 土层在人工种植草本治理情况下, 生长过程受地表(0~10 cm)夏季高温、冬季低温、夏季集中降水、冬季更为恶劣的冻融条件影响(图1), 形成了和10~20 cm极大的环境差异, 而地表更多的集中了凋落物归还, 浅层根系分解, 所以环境的和种植反馈过程的多因素作用差异也可能是造成两个土壤层各土壤指标差异较大的原因[36, 44, 45]。因此在进行指标的综合考虑过程中, 容重可以作为衡量不同种植模式下的指标之一, 而综合得分较高的有机碳、全氮、pH值3种土壤指标中, pH值由于和地表植物群落相关性不高, 因此需要单独评价, 不适宜在评价指标中作为主要参考因素。因此建议将有机碳、全氮和容重作为不同种植模式下土壤性质变化基本的评价指标。
以一年生和多年生复合草本对川西北高寒半湿润沙地进行治理, 采用行播、带播、撒播3种模式进行沙化恢复。综合地上生物特征和地下土壤特征, 发现4年的带播和撒播种植模式对川西北高寒半湿润沙地恢复效果良好。在选择的恢复草本物种中, 一年生的燕麦无法完成自我繁殖过程, 多年后从生态系统中消退, 而多年生草本可以完全适应川西北高寒半湿润生境。带播和撒播模式明显增加了地表植物群落盖度, 植物物种多样性和生态优势度, 明显减小了地表植物群落均匀度指数; 其中效果最佳的撒播模式盖度增加至80%, 植物物种数增加至9种。带播和撒播显著减小了土壤容重, 增加了土壤含水量、有机碳、全氮、全磷、硝态氮、有效磷含量。不同种植模式地表植物群落特征和地下土壤性质具有密切关系, 而地表植物群落变化对土壤各指标的影响机理需要进一步深入研究。通过主成分分析后发现, 降维后的土壤指标中, 土壤容重、有机碳、全氮可以作为土壤性质变化的主要衡量因子, 而pH值受限于不同土层极大的环境差异、物质反馈等, 暂不建议作为川西北高寒半湿润沙地草本恢复土壤的评价指标。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|