

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
典型草原建群种长芒草( Stipa bungeana)在中国的潜在分布范围预测及主要影响因子分析
[王百竹1  , 朱媛君
, 朱媛君1 , 刘艳书1 , 马风云2 , 张晓1 , 时忠杰1 , 杨晓晖1, * ]
, 朱媛君]
|
|


作者简介:王百竹(1996-),女,山东济南人,在读硕士。E-mail: wangbaizhueva@126.com
以广泛分布在中国北方典型草原的建群种长芒草为研究对象,利用Maxent模型对长芒草在中国当前及未来气候变化下的潜在分布区进行预测并对主要影响其分布的环境变量进行分析,结果表明,采用受试者工作特征曲线(receiver operating characteristic curve, ROC)对模型精度进行检验所得到的训练数据与测试数据的受试者工作特征曲线面积(area under ROC curve,AUC)分别为0.962和0.950,表明模型预测结果可靠,当前中国长芒草高适宜性分布区主要有5个,分别为黄土高原分布区、泰山-沂蒙山分布区、横断山分布区、藏南谷地分布区及天山分布区。在RCP2.6(representative concentration pathways 2.6)和RCP8.5(representative concentration pathways 8.5)两种气候情景模式下预测得到的2070年长芒草最适宜的潜在分布区有逐渐缩小的趋势。Jackknife检验对主导环境变量的筛选结果显示,影响长芒草分布的主要环境变量有地形粗糙度指数(terrain roughness index, tri)、9月降水量(precipitation 09, prec09)、气候湿度指数(climatic moisture index, topowi)、2月最高温度(maximum temperature 02, tmax02)、12月降水量(precipitation 12, prec12)和12月平均温度(average temperature 12, tavg12)。结果可为气候变化背景下中国典型草原的可持续管理提供科学依据。
, ZHU Yuan-jun
Stipa bungeana is a dominant species of typical grassland in the north of China. In this study, we project its potential distribution patterns under current and future climate scenarios using the Maximum Entropy Model, and we identify the major factors influencing the distribution patterns. Initial testing of the model indicated results were reliable. In current climate scenarios, there were 5 highly suitable areas for S. bungeana: the Loess Plateau region, the Tai-Yimeng mountain region, the Hengduan mountains, the southern Tibet valley and the Tianshan mountain area. Under climate scenarios RCP 2.6 and RCP 8.5 (RCP denotes 'representative concentration pathway') in 2070, the highly suitable areas for S. bungeana would decreased. The results from a jackknife test showed that terrain roughness index, September precipitation, SAGA-GIS topographic wetness, February maximum temperature, December precipitation and December average temperature of December were major environmental variables affecting S. bungeana distribution patterns. These results provide a theoretical basis for the sustainable management of typical grassland in China under climate change.
物种分布模型(species distribution models, SDMs)利用物种的样本分布信息和对应的环境变量信息, 依据特定的算法估计物种的生态位及其栖息地适宜性并投影到景观中, 以概率的形式反映物种对生境的偏好程度, 并生成预测物种出现或适宜的栖息地分布图[1, 2], 其结果可以解释物种出现的概率、生境适宜度或物种丰富度等[3, 4]。经过近20年的发展, 物种分布模型已经成为基础生态学和生物地理学研究的重要工具, 在环境科学研究、自然资源管理和生物多样性保护等方面具有重要的应用价值, 广泛应用于研究全球变化背景下物种的分布与气候间的关系[5, 6, 7, 8, 9]、区域气候变化对植物群落和功能的影响[10, 11]、生态系统功能群和关键种的监测及预测[12]、生态系统不同尺度多样性的管理和保护[13]、外来物种入侵区域的预测[14]以及面向生态系统恢复的关键物种的潜在分布预测和保护区规划[15]等诸多方面, 可以为区域生态系统的保护及可持续发展提供强有力的理论支撑。Maxent模型是一种源自统计力学的物种分布模型(SDMs), 它是预测物种潜在分布的环境模型[16, 17], 因其具有预测精度稳定可靠、连续数据和分类数据均可作为输入变量、可直接产生空间明确的栖息地适宜性图、内置环境变量重要性评估工具等优点而被广泛使用[18]。
长芒草(Stipa bungeana)为禾本科(Gramineae)针茅属(Stipa)的一种多年生、旱生丛生禾草, 是我国针茅属植物分布最广的种类之一, 且是我国暖温型典型草原的建群种, 常见于石质干燥坡地, 主要分布于我国西部, 从东北、华北、西北、西南, 向东延伸到江苏、安徽, 在蒙古、日本也有分布[19, 20, 21]。以长芒草为建群种的草地群落构成了欧亚大陆草原区的最典型地带性植被群落[22]。长芒草叶面产量高, 适口性好, 是我国北方天然草原上主要的野生饲料物种, 同时在保护土壤免受侵蚀和减少水分流失方面也发挥着重要作用[23]。近年来, 在严重的人为干扰和气候变化等多重因素影响下, 我国北方草原天然植被严重退化, 物种多样性锐减, 植被生产力低下, 自然更新能力差, 导致长芒草群落大范围衰退, 且在自然条件下自身的逆转和恢复较为困难, 这对我国北方的畜牧业产生了严重影响[24]。因此, 预测长芒草在中国的最适潜在分布区以及对未来气候变化的响应有着重要的生态学意义, 同时对于我国北方畜牧业生产也具有一定的指导意义。
1.1.1 物种分布数据获取 长芒草分布点的地理位置信息主要通过检索中国数字植物标本馆(http://www.cvh.org.cn)、中国科学院植物研究所植物标本馆(http://pe.ibcas.ac.cn)、中国自然标本馆(http://www.nature-museum.net)等数字标本库获取, 排除掉重复的坐标点和信息不完整的标本, 共获取到345个长芒草分布记录, 这些数据的经纬度由Excel存为CSV格式, 用于Maxent模型分析。
1.1.2 环境变量数据的获取 本研究中所采用的环境变量由生物气候变量和地形变量两部分组成。生物气候变量对于定义一个物种的环境生态位具有非常重要的生物学意义。从世界气候数据网站(http://www.worldclim.oig)下载了84个气候变量(表1), 数据的空间分辨率为30″(1 km× 1 km); 相关的5个地理变量从ENVIREM(http://envirem.github.io/)中获取。IPCC(The Intergovernmental Panel on Climate Change)第5份报告描述了未来的4种气候变暖情景, 即在2100年总辐射强迫(radio frequency, RF)基础上的4种典型浓度路径(representative concentration pathways, RCP), 用于表述在4种不同路径下21世纪温室气体排放及其大气浓度、空气污染物排放和土地利用的情况。RCP中包括一类严格减缓情景(RCP2.6)、两类中度排放情景(RCP4.5和RCP6.0)和一类温室气体高度排放情景(RCP8.5)。本研究选取了2070年RCP2.6和RCP8.5两个情景下的生物气候数据。本研究所有环境变量使用统一的分辨率(30″)、范围和地理坐标系统, 最终获取气候变量84个、地理变量5个, 总计89个环境变量(表1)。
| 表1 全部环境变量 Table 1 List of environmental variables |
1.2.1 环境变量的筛选 由于相同数据来源的生物气候变量之间具有互相关性, 本研究使用R语言中的“ Maxent Variable Selection” 包对89个环境变量进行筛选, 最终获得出13个互相关性系数< ± 0.8且贡献率> 5的气候变量, 加上5个地理变量一共18个环境变量(表2)用于长芒草的物种分布模型构建。
| 表2 用于长芒草SDMs构建的环境变量 Table 2 The selected variables for SDMs of S. bungeana |
1.2.2 最大熵模型的构建和检验 本研究使用Maxent 3.3.3[25]软件包及ArcGIS 10.2进行建模。Maxent是一种从不完全信息中做出预测或推断的通用方法, 其通过寻找最大熵的概率分布来估计一个物种的分布, 可最大程度不约束物种未知分布信息, 更多地保留物种现有分布的环境变量数据, 在物种部分信息缺失的情况下也能较好地对可能分布区做出预测[17]。
模型的训练数据为样本数据随机选择的75%, 测试数据为样本数据剩余的25%。Maxent设定模型运算次数1000次, 进行10次重复运算, 计算了每个变量的ROC(receiver operating characteristic curve)曲线, 并利用软件内嵌的Jackknife检验, 对每个变量的作用进行计算。Jackknife检验可以用来判断各生态变量对分布预测的贡献度, 帮助筛选主导性的环境变量[26]。Maxent软件包使用AUC(area under roc curve)值来评估模型的性能, AUC值为ROC曲线与坐标轴之间的面积, 其值越大, 模型可靠性就越好。一般来说, AUC的值处于0.5到1之间, 当0.5< AUC≤ 0.6时, 模型拟合失败; 0.6< AUC≤ 0.7时, 模型可靠性弱; 0.7< AUC≤ 0.8时, 模型可靠性一般; 0.8< AUC≤ 0.9时, 模型可靠性良好; 0.9< AUC≤ 1时, 模型可靠性优秀[27, 28]。将Maxent模型生成的长芒草潜在分布图导入ArcGIS转换为栅格图, 并采用标准分级中的自然间断点分级法(Jenks’ natural breaks)对长芒草潜在适宜分布区进行适宜性等级划分[29], 将适宜性大于0.51的分布区确定为最适分布区。
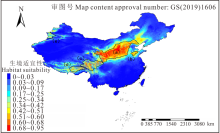
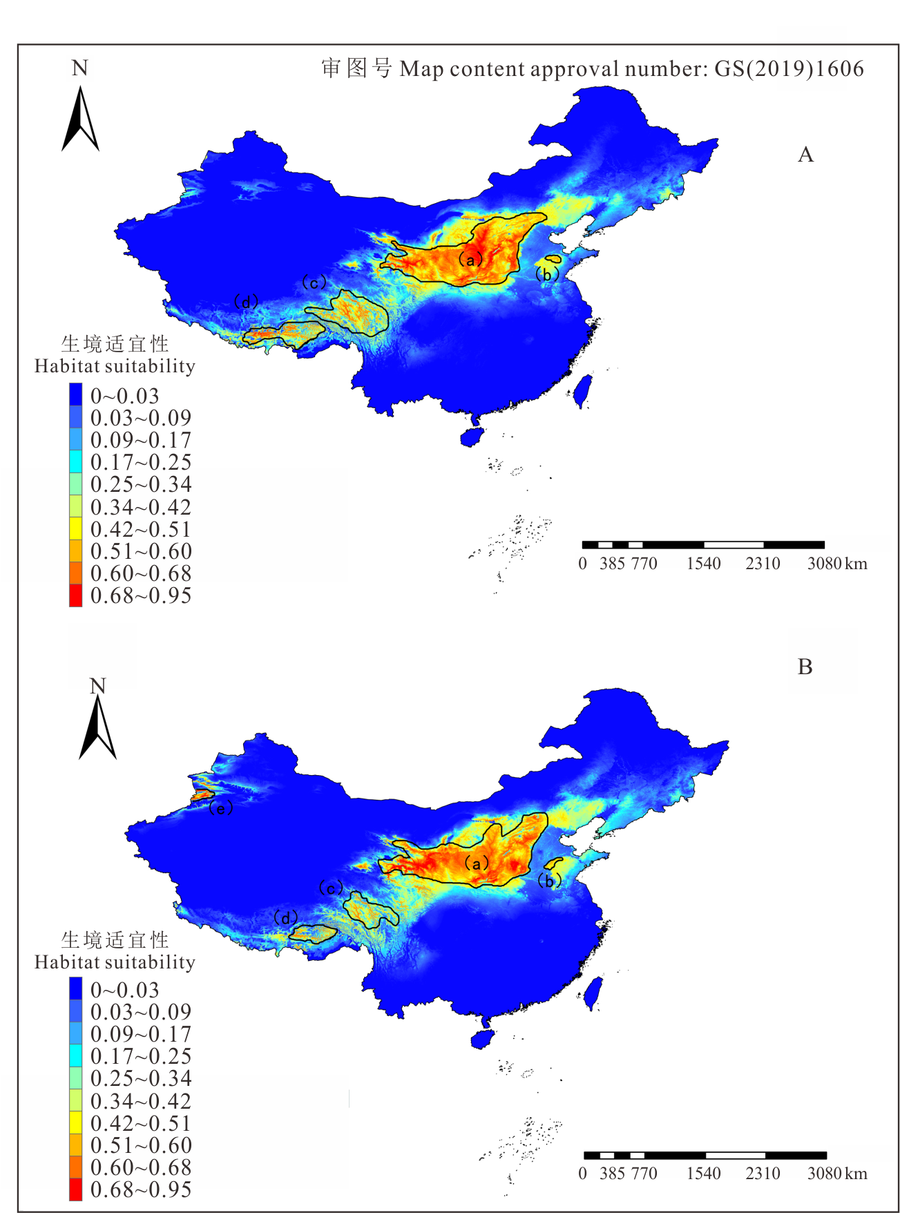
图1是中国长芒草生境适宜性分布图, 如图所示, 长芒草分布范围较广, 东起华北平原东部, 西到新疆天山山脉, 北至黄土丘陵北部及阴山山脉, 南至横断山区南端, 其中, 晋、陕、甘、宁及内蒙古南部, 即暖温带的黄土高原地区, 是长芒草集中分布的区域。本研究按生境适宜性在0.51以上的范围, 划分了5个最适分布区:a区是以黄土高原为主的分布区; b区是以山东省泰山-沂蒙山区为主的分布区; c区以横断山脉为主, 向东到青藏高原东部, 南至香格里拉, 北至青海省南部; d区是以喜马拉雅山为界的藏南谷地地区; e区则位于新疆天山山脉。由此分布区划分可以看出, 长芒草适合生长在石质山坡、黄土丘陵、河谷阶地等位置。
 | 图1 长芒草生境适宜性分布Fig.1 Habitat suitability of S. bungeana |
图2a显示了测试数据和训练数据的遗漏曲线与预测区域遗漏曲线随累积阈值的变化情况, 实测遗漏率最大程度接近于预期的遗漏率时, 代表模型拟合效果最佳。在本研究中, 实测遗漏曲线与预测遗漏曲线几乎重合, 表示模型的可靠性极佳。为了进一步测试Maxent的预测精度, 本模型使用受试者工作特征(ROC)曲线进行检验模型精度, 由图2b可知, 训练数据的AUC值为0.962, 测试数据的AUC值为0.950, 表明Maxent模型预测结果可靠性极佳, 其对长芒草的潜在分布的预测是非常准确的。
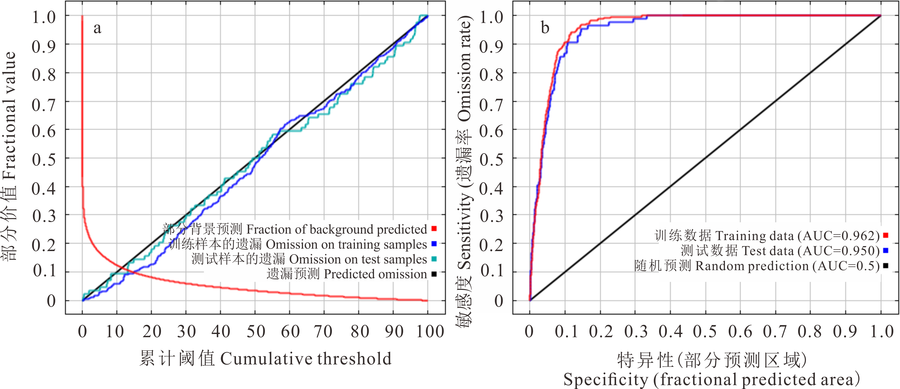 | 图2 长芒草模型精度检验Fig.2 Model accuracy test of S. bungeana a:遗漏调试分析曲线Analysis curve of omission/commission; b:ROC曲线Receiver operating characteristic curve. |
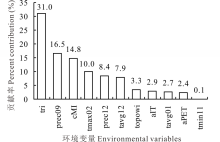
对Maxent模型中各环境变量贡献率的统计结果表明(图3), 共有6个环境变量对模型的影响贡献率大于5%, 由高到低依次为地形粗糙度指数(tri, 31.0%)、9月降水量(prec09, 16.5%)、气候湿度指数(cMI, 14.8%)、2月最高温度(tmax02, 10.0%)、12月降水量(prec12, 8.4%)和12月平均温度(tavg12, 7.9%), 其累积贡献率达88.6%, 是模拟长芒草适生区主要的环境变量, 能反映其最适生境分布的大部分信息; 同时, 所有筛选出的变量, 在模型中都有不同的贡献率, 贡献最小的变量为11月, 最低气温也在0.1%左右, 说明没有无关变量参与Maxent模型分析, 可见该筛选方法对构建Maxent物种分布模型环境过程中的变量筛选是十分有效的。
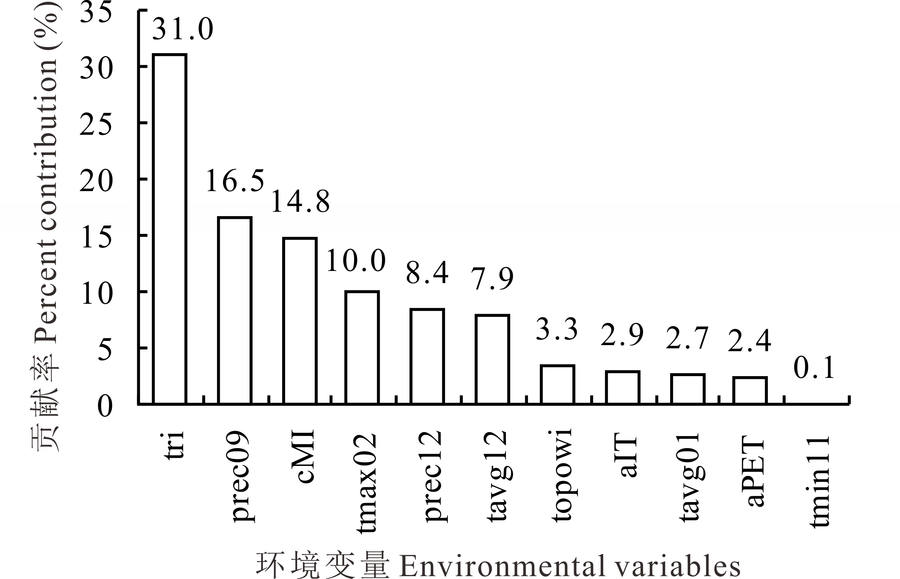 | 图3 环境变量贡献率Fig.3 Analysis of enviromental variables contributions |
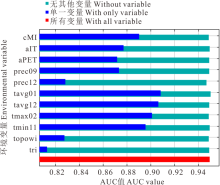
环境变量对长芒草潜在适宜分布区预测模型单一变量贡献率大小的Jackknife 检验结果(图4)显示, 5 个环境变量(cMI、tavg01、tavg12、tmax02、tmin11)的 AUC值均> 0.88, 说明它们是预测长芒草潜在适宜分布区的主要影响变量; 其中tavg01、tavg12和tmax02的单一变量AUC值均超过了0.90, 表明1和12月的平均气温及2月的最高气温是影响长芒草物种适宜分布区最主要的变量, 显示长芒草分布主要受其种子开始萌发的前3个月的气温影响最大。而tri的单一变量AUC值最低, 说明长芒草对地表粗糙度的适生性很强, 因此其在黄土丘陵区, 到石质山坡, 再到河谷阶地均有分布。
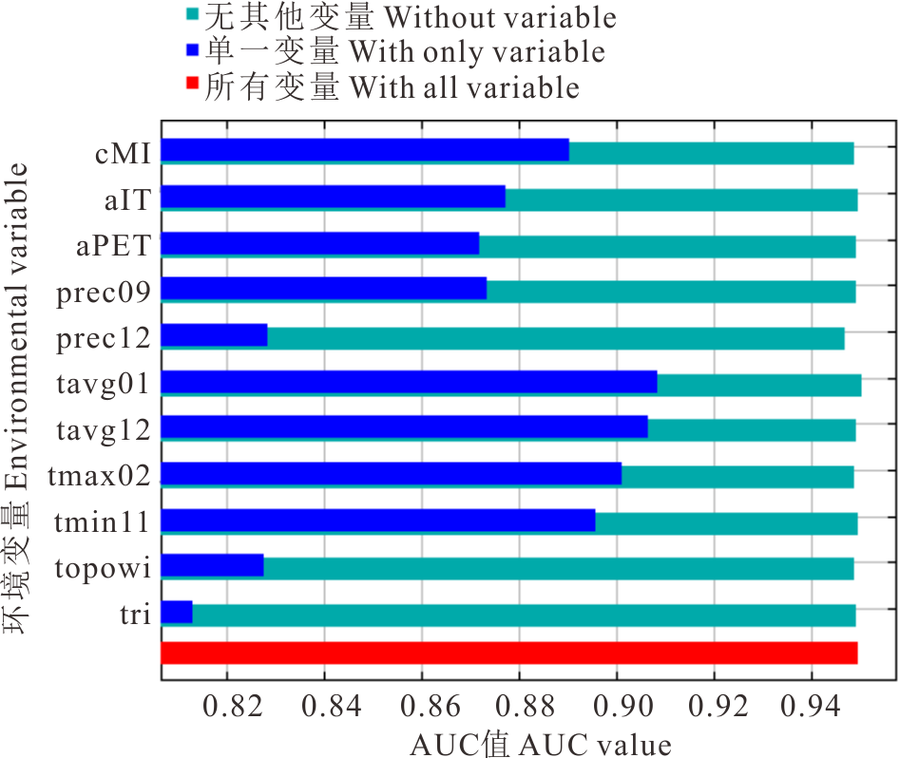 | 图4 长芒草SDMs Jackknife检验Fig.4 Jackknife test of S. bungeana SDMs |
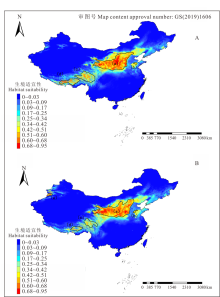
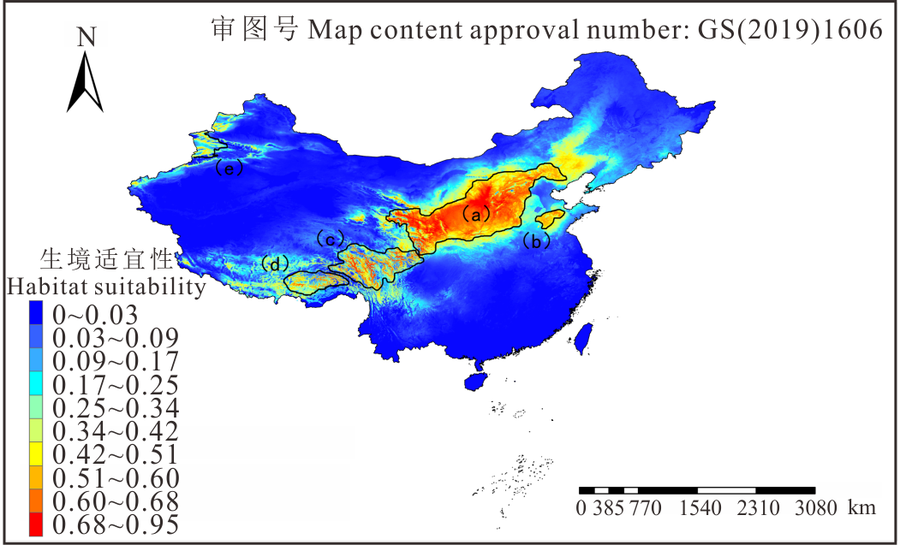
图5表示2070年RCP2.6和RCP8.5两种气候情景下的生境适宜度分布。图5A是RCP2.6气候情景模式下长芒草生境适宜度分布图, 整体上来看a区的面积有所减少, 但0.68以上的最适宜分布区的面积更加集中, 高适分布区中心位于黄土高原中部的甘肃省和山西省部分地区, 越往外适宜度越低, 同时b、c、d区面积减少, 图1中长芒草分布的e区, 即天山山脉分布区在该情景模式下则完全消失, 而山东、河北、辽宁三省的大部分地区都成了长芒草的中适分区。
| 图5 长芒草在两种气候变暖情形下的生境适宜性分布Fig.5 Suitable habitat distribution of S. bungeana at two globe warming situations A: RCP2.6, Representative concentration pathways 2.6; B: RCP8.5, Representative concentration pathways 8.5. 下同 The same below. |
图5B是2070年RCP8.5气候情景模式下长芒草生境适宜度分布图, 很明显与RCP2.6的相比, a区最适宜分布面积减少, 且更为分散, 由黄土高原中部往东西两边扩散, 主要集中在山西省南部以及青藏高原东北部。b区面积减少且发生西移, c、d两区的面积进一步减少, 且生境适宜性0.68以上的区域与当前气候下和RCP2.6情景模式相比, 分布范围缩小, 青藏高原整体的中适分布区域增加, 一直延伸至青藏高原南部, 但低适区域减少, e区是位于新疆北部天山山脉的分布区域, 相比当前气候下分布区面积整体减少, 但是在伊犁河谷附近出现了一部分0.68以上的最适分布区。山东、河北、辽宁三省的大部分地区都成了长芒草的低适分区, 整个东北地区及内蒙古北部的低适生区域大面积减少。
环境变量对长芒草最适分布区预测贡献的Jackknife 检验结果显示(图6), 在RCP2.6情景模式下, cMI、bio11、bio12、bio09、prec09、tmax11、tmax01在Jacknife 检验中AUC> 0.75, 说明这些变量在模型构建过程起到了主要作用; 而其他环境变量如topowi、tri等的AUC较低, 表明这些变量对于模型构建的贡献较低; 在RCP8.5情景模式下, cMI、bio11、bio09、prec04、tmin03在Jacknife 检验中AUC> 0.75, 说明这些变量在该情景下的模型构建起到了主要作用。通过对当前气候及2种未来气候变化的情景下长芒草的SDM分析, 发现对长芒草Maxent模型起主要作用的环境变量发生了变化, 当前气候条件下对长芒草最适分布区起主要作用的环境变量主要是气温, 而随着未来气候变暖, 温度逐渐升高, 主要发挥作用的环境变量由原来的气温逐渐转为降水, 9月降水量和气候湿度指数都成为未来气候条件下影响长芒草最适分布区的重要环境变量。
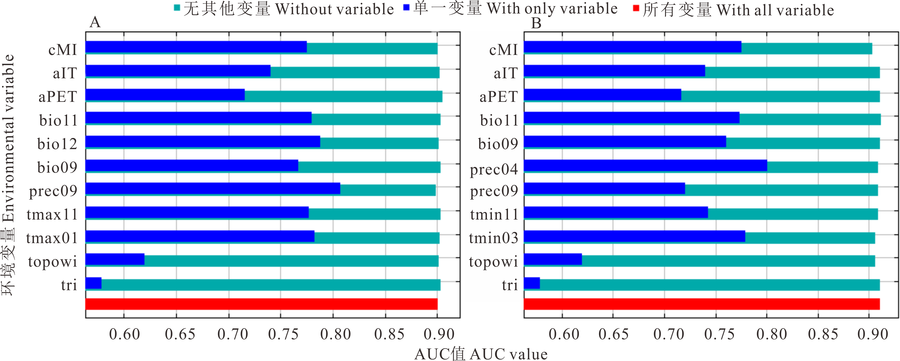 | 图6 长芒草在两种气候变暖情形下的AUC值Jackknife检验Fig.6 Jackknife of AUC of S. bungeana at two globe warming situations |
胡忠俊等[30]基于Maxent模型和GIS的青藏高原紫花针茅(S. purpurea)分布格局模拟研究中, 紫花针茅分布与生长季降水、年均降水量和最湿季度降水量有很强的相关性, 意味着紫花针茅的分布区主要处于青藏高原年降水量特别是生长季降水量较多的区域; 而最干季度降水量及最冷季度降水量的贡献率很小, 表明紫花针茅对于极端环境及温度的忍耐程度较高。陈俊俊等[28]在应用Maxent 模型对短花针茅(S. breviflora)在中国的潜在分布区研究及预估研究时, 利用Jackknife检验分析了环境因子对短花针茅潜在分布的影响, 得出排在前2位的因子是最冷季度的降水量和年平均气温, 最冷季度的降水是指12月-次年2月的降水, 主要是因为短花针茅的返青期大约在每年的4月, 这时短花针茅能够使用的土壤中的水主要是冬季储存在土壤中的水分, 所以最冷季度的降水为返青提供了非常好的便利条件; 第二个主要环境因子是年平均气温, 该因素主要控制着短花针茅的种子繁殖。本研究中Maxent模型结果表明, 影响长芒草适宜性分布的主要环境因子有地形粗糙度指数、9月降水量、气候湿度指数、2月最高温度、12月降水量和12月平均温度, 这6个环境变量的总贡献率达到了88.6%, 结合Jackknife检验结果显示, 温度对于长芒草的分布影响最大, 根据结果显示的环境因子有12月平均温度、1月平均温度、2月最高温度, AUC值均超过了0.90, 即最冷季的温度是影响长芒草分布的主要环境因子。与长芒草相比, 紫花针茅为高寒草原的建群种, 因其主要分布在青藏高原, 物种本身对于低温的忍耐程度高, 所以影响其分布的主要环境变量不是温度, 而是降水; 而短花针茅是荒漠草原的建群种, 荒漠草原区物种的发育主要受降水的制约[29], 同时最冷季的降水直接关系到春季短花针茅种子的萌发, 因此影响短花针茅分布的主要环境变量为最冷季降水。与这两种针茅不同, 长芒草为典型草原建群种, 在年降水392~376 mm 的地区均可较好发育[21], 且长芒草本喜暖耐旱, 在降水较多的华北森林草原区可以很好生长, 在干旱的天山山脉也有分布, 因此对于水分的忍耐程度较大[31]; 而有研究表明, 恒温条件下, 随温度的升高, 长芒草种子发芽率呈现先升后降的趋势, 20 ℃条件下发芽率达到最高, 在5~20 ℃之间时, 发芽指数随温度的增加而增加, 20 ℃以后, 发芽指数随温度的增加而下降[32]; 从3月末到4月初, 长芒草开始萌发, 大多数种子在6月下旬成熟[33], 种子成熟后会有一些处于休眠状态, 只有经过一段时期的低温处理, 再经过温度升高气温变暖, 才能解除休眠状态, 进入萌芽期, 在一定程度上, 长芒草种子萌发的前3个月的环境温度对于其种子萌发率起到了重要影响[32], 而植物种子通过风媒、虫媒、动物迁徙携带等途径进行传播, 进而影响物种分布, 那么种子传播到新的环境后, 能否成功萌发是直接影响该物种分布的主要因素, 因此, 温度成了制约长芒草分布的主要环境变量。孙建华等[34]对退化草地土壤种子库的研究发现, 土壤种子库密度随草地退化程度(放牧强度)的增加而显著下降, 长芒草主要分布在水土流失严重的黄土丘陵区, 干旱、半干旱气候的特点就是降水少、蒸发量大, 相比地下, 地表处于相对湿润状态的持续时间较短, 因而, 降水可满足种子萌发对水分的需求, 但地表快速的干旱极易引起幼苗夭折, 从而造成种子库的萌发损耗, 最终导致种子萌发率均随水分胁迫的增加而降低[35, 36, 37], 在未来气候变暖的情况下, 随着草地退化程度的加深, 降水因素将替代温度, 成为影响长芒草生长分布的关键环境变量。
长芒草栖息地适宜性的Maxent结果表明, 在RCP2.6和RCP8.5两种气候情景模式下, 随着气候变暖, 预测得到的2070年长芒草最适宜的潜在分布区有逐渐缩小的趋势, 最适潜在分布区整体明显减少, 大部分的低适宜分布区消失, 且长芒草高适宜度的栖息地向中适宜度转变。长芒草是我国北方典型草原的建群种和优势种, 且为草原上最主要的野生饲料物种之一, 因此保护长芒草群落对于维持草原植被的稳定性及促进我国北方畜牧业的发展尤为重要。同时, 由于近年来的过度放牧及黄土高原水土流失加剧, 长芒草的生境不断缩减和退化[38], 为有效保护长芒草草原生境, 应建立合理的草原生态补偿机制, 制定有效的放牧管理制度, 实施科学的草畜平衡、围栏放牧[39]。
本研究利用基于最大熵理论的Maxent模型并结合地理信息系统, 综合分析了长芒草在当前及两种未来气候情景下的潜在适宜性分布及影响潜在适宜分布区的主导环境变量, 得出以下结论:
1)利用最大熵模型预测长芒草的潜在分布区, 模型训练数据与测试数据的精度分别为0.962和0.950, 根据AUC指数的评价结果, 该模型结果是精确可靠的。
2)在模型构建时选择的11个环境变量中, 长芒草的分布主要受6个变量的影响:地形粗糙度指数、9月降水量、气候湿度指数、2月最高温度、12月降水量和12月平均温度。其中, 影响当前气候条件下的长芒草最适分布区的环境变量主要为温度, 而影响未来气候情景下长芒草最适分布区环境变量逐渐转变为降水和地表湿度。
3)长芒草栖息地适宜性的Maxent结果表明, 在气候变化的背景下, 长芒草的最适潜在分布区整体明显减少, 大部分的低适宜分布区消失, 且长芒草高适宜度栖息地向中适宜度转变。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|