

{kind=link}
{kind=link}
{kind=link}
{kind=link}
4种间作模式下牧草根系特性及其碳、氮代谢特征研究
[蔺芳1, 2  , 刘晓静
, 刘晓静1, * , 童长春1 , 吴勇1 ]
, 刘晓静, 童长春|
|


作者简介:蔺芳(1983-),女,河南新乡人,在读博士。E-mail: fanglin2035@126.com
为探究间作对牧草根系生长特性和碳、氮代谢特征的影响,通过田间试验,以紫花苜蓿、小黑麦、燕麦、玉米、甜高粱5种单作模式为参照,对紫花苜蓿/小黑麦、紫花苜蓿/燕麦、紫花苜蓿/玉米和紫花苜蓿/甜高粱4种间作模式进行了研究,采用目前先进的微根管观测系统并结合计算机成像技术对紫花苜蓿和4种禾本科牧草单作及间作下根系的生长状况进行了原位观测,并且对4种间作模式下碳、氮代谢关键酶及产物进行了研究。结果表明,无论单作还是间作,根系参数与碳、氮代谢各指标均呈正相关关系,其中根长、根表面积均与核酮糖-1,5-二磷酸羧化酶(RuBPCase)和硝酸还原酶(NR)活性呈极显著正相关( P<0.01),与蔗糖磷酸合成酶(SPS)和谷氨酰胺合成酶(GS)活性呈显著正相关( P<0.05),表明根系特性与碳、氮代谢具有一致性;4种间作模式下禾本科牧草的根长、根表面积等根系参数明显优化,而紫花苜蓿根系特性与单作相比表现不一,与玉米和甜高粱间作的紫花苜蓿其根系特性受到了明显地抑制(显著低于单作),表现为竞争弱势,而与小黑麦和燕麦间作的紫花苜蓿则未受影响;4种间作模式下禾本科牧草功能叶RuBPCase和NR活性与单作比均显著性增加,而紫花苜蓿功能叶RuBPCase和NR活性与单作相比趋势不尽相同,与玉米和甜高粱间作的紫花苜蓿其RuBPCase和NR活性显著低于单作,而与小黑麦和燕麦间作的紫花苜蓿则差异不显著。由此可见,供试牧草根系特性和碳、氮代谢特征表现趋势一致;在4种间作模式下根系特性和碳、氮代谢特征均表现为禾本科牧草优于豆科牧草;紫花苜蓿/小黑麦和紫花苜蓿/燕麦这2种间作模式下综合表现最佳。
This study aimed to explore the effects of intercropping on root growth characteristics and carbon and nitrogen metabolism characteristics of some common forage crops. Five species monocultures (alfalfa, triticale, oat, maize and sweet sorghum) were used as reference crops in a field experiment, and four intercropping combinations (alfalfa/triticale, alfalfa/oat, alfalfa/maize and alfalfa/sweet sorghum) were evaluated. The root system growth of alfalfa and the four Poaceae crops under monoculture and intercropping were observed in situ through advanced minirhizotron and computer imaging technology, and the key enzymes and products of carbon and nitrogen metabolism under the four intercropping combinations were studied. It was found that the root parameters were very significantly ( P<0.01) and positively correlated with carbon and nitrogen metabolism whether in monoculture or intercropping. Root length and surface area were significantly ( P<0.05) and positively correlated with ribulose-1,5-bisphosphate carboxylase (RuBPCase) and nitrate reductase (NR) activity, and also with sucrose phosphate synthase (SPS) and glutamine synthetase (GS) activity, indicating that the root characteristics were consistent with carbon and nitrogen metabolism. Root length, root surface area and other root system parameters of the four tested Poaceae forages were significantly enhanced when intercropped with alfalfa, while alfalfa performance differed between monoculture and intercropped treatments. Root systems of alfalfa intercropped with maize and sweet sorghum were significantly inhibited (significantly smaller than in monocropping), and the alfalfa plants showed weak competitive ability. However, root systems of alfalfa intercropped with triticale and oat were unaffected. When intercropped with alfalfa, functional leaf RuBPCase and NR activities of the four Poaceae species were significantly increased, compared with monocropping. For alfalfa functional leaves, RuBPCase and NR activities of alfalfa plants intercropped with maize and sweet sorghum were significantly lower than in monoculture, while leaf enzyme activities of alfalfa intercropped with triticale and oat were not significantly changed, compared to monoculture. In summary, in this study the root system development of the tested forages mirrored the leaf carbon and nitrogen metabolism differences, and the four grasses outcompeted alfalfa in the intercropping treatments. The two intercropping mixtures alfalfa/triticale and alfalfa/oat had the best overall performance.
根系是土壤的利用者和产量的贡献者, 其起到上接地上部植株下启地境的关键作用[1], 因此对根系特性进行研究具有十分重要的现实意义。越来越多的事实证明, 根系构型是影响植物生长的重要因素, 反映了植物具有的营养遗传性状[2], 所以根系的许多指标, 如根长、根尖数、根表面积、根体积等, 均可以作为衡量植物个体生长发育状况和代谢能力的重要参数[3]。由于作物根系生长过程中的不可见性, 根系形态检测与研究发展缓慢, 传统的根系研究方法主要是利用挖掘法、剖面法和土钻法[4]对根系样本开展研究, 均属于破坏性取样, 不但会对研究对象的正常生长产生干扰, 而且也不能长期对同一根系的生长和分布进行追踪, 操作方法费时费力, 误差较大。实现对根系形态的实时、准确获取是农业生产中提高作物品质和产量的前提和关键[5]。目前, 微根管法是监测根系生长动态的新技术, 可在非破坏条件下真实地反映植物的根系生长情况[6], 是一种成本低、操作方便并更能接近作物田间实际状态的根系形态原位测定方法。
植物的根系特性对地上部植株的生长和碳、氮代谢能力等方面具有重要影响[7]。作物的生长和发育所需的养分主要通过根系的吸收获得, 根系吸收水和养分输送到地上部, 地上部通过碳、氮代谢合成有机物质, 供植物体正常生长与发育[8]。碳代谢和氮代谢作为植物体内两大最基本的代谢过程, 在植物的生长发育过程中起到了至关重要的作用, 二者共同调节碳水化合物和蛋白质的合成, 共同影响且决定着作物的产量和品质[9]。其中, 植物叶片中的核酮糖-1, 5-二磷酸羧化酶(ribulose-1, 5-bisphosphate carboxylase, RuBPCase)和蔗糖磷酸合成酶(sucrose phosphate synthase, SPS)分别是光合固定与糖转化过程中的关键酶, 硝酸还原酶(nitrate reductase, NR)与谷氨酰胺合成酶(glutamine synthetase, GS)分别是植物体内硝态氮转化与铵态氮同化过程中的关键酶, 它们作为碳、氮代谢的关键酶, 在碳、氮代谢过程中发挥着重要作用[10]。
豆科/禾本科间作是目前我国农业生产中应用最为广泛的一种多元种植模式。在豆科/禾本科间作系统中, 豆科固氮不仅可以维持间作系统运转所需的养分, 还可以通过减少氮素的投入降低土壤环境中硝酸盐含量, 因此不论是从资源利用方面还是对环境的贡献方面, 这种组合都是一个可持续的生态系统[11]。在间作种植模式中, 由于两种作物种植在同一块土地上, 这两种作物既可能发生相互促进作用, 同时也会竞争环境资源和生长空间, 包括植株地上部、地下部对资源的竞争。目前普遍认为在豆科/禾本科间作系统中禾本科作物具有更强的竞争优势, 是系统中的优势物种, 并且对系统的总生物量有决定性的影响[12]。目前, 在豆科/禾本科间作的研究中, 有关其根系和碳、氮代谢的研究多集中于粮食作物/牧草间作, 而在牧草/牧草间作上进行的研究较少。本研究通过原位扫描技术对4种豆科/禾本科牧草间作模式[紫花苜蓿(Medicago sativa)/小黑麦(Secale sylvestre)、紫花苜蓿/燕麦(Avena sativa)、紫花苜蓿/玉米(Zea mays)、紫花苜蓿/甜高粱(Sorghum dochna)]下的牧草根系特性及其碳、氮代谢特征进行监测分析, 以便于更好地认识和了解4种豆/禾牧草间作系统地下根系-地上碳、氮代谢连续体的相互关系。
试验地位于新乡市洪门镇原堤村(35° 16'7″ N, 113° 57'31″ E), 地处豫北平原, 该区属暖温带大陆性季风气候, 四季分明, 冬寒夏热, 秋凉春早。年均气温14.2 ℃, 年均降水量573.4 mm, 年均日照时数2400 h, 年均湿度68.0%, 无霜期220 d。降水年际间分配不均, 主要集中在7-8月。供试土壤类别为砂壤土, 供试土壤0~20 cm的理化性质为:全氮1.22 g· kg-1, 全磷0.68 g· kg-1, 全钾15.44 g· kg-1, 碱解氮42.67 mg· kg-1, 速效磷18.35 mg· kg-1, 速效钾89.57 mg· kg-1, 有机质9.56 g· kg-1, pH值7.88。前茬作物为玉米。
试验材料均为饲用型牧草, 其中小黑麦由河北省农业科学院提供, 燕麦和甜高粱由北京克劳沃草业技术开发中心提供, 玉米由河南省农业科学院提供, 紫花苜蓿由北京猛犸种业有限公司提供。供试材料的品种及生育期见表1。
| 表1 供试品种及生育期 Table 1 Tested varieties and growth period |
试验设计为完全随机区组, 设紫花苜蓿、燕麦、小黑麦、玉米、甜高粱5种单作模式以及紫花苜蓿分别与以上4种禾本科牧草间作, 共9个处理, 每处理重复3次, 共27个小区, 小区面积4 m× 5 m=20 m2, 总面积540 m2。为了防止小区间的相互影响, 每个小区之间用70 cm小田埂分开, 每个小区一侧有水渠, 排灌分开。各处理以生产中常见的种植方式进行单作或间作, 其中, 单作紫花苜蓿的播量为15 kg· hm-2, 条播, 行距20 cm; 单作小黑麦的播量为225 kg· hm-2, 条播, 行距20 cm; 单作燕麦的播量为180 kg· hm-2, 条播, 行距20 cm; 单作玉米和甜高粱的播种密度为60606株· hm-2, 穴播, 行距50 cm, 株距33 cm。紫花苜蓿/小黑麦和紫花苜蓿/燕麦间作系统中, 种植比例为3∶ 2, 即3行紫花苜蓿和2行小黑麦(燕麦), 间作下紫花苜蓿与小黑麦(燕麦)的行间距为30 cm, 间作下紫花苜蓿、小黑麦、燕麦的播量和行距同单作; 紫花苜蓿/玉米和紫花苜蓿/甜高粱间作系统中, 种植比例为5∶ 2, 即5行紫花苜蓿和2行玉米(甜高粱), 间作下紫花苜蓿与玉米(甜高粱)的行间距为35 cm, 间作下紫花苜蓿、玉米、甜高粱的播种密度和行距同单作。于2016年10月2日播种紫花苜蓿, 2016年10月6日和2017年10月2日播种小黑麦, 2017年3月23日和2018年3月25日播种燕麦, 2017年和2018年5月20日同时播种玉米和甜高粱。试验期间不施用任何肥料, 田间常规管理。
1.4.1 测定时期与测定位点 于紫花苜蓿的初花期(10%开花)进行刈割, 共刈割5茬, 分别是2018年5月1日(第1茬)、6月10日(第2茬)、7月15日(第3茬)、8月20日(第4茬)和10月1日(第5茬)。其中, 5月1日同时进行了紫花苜蓿(第1茬)和小黑麦(抽穗期)的各指标测定, 6月10日同时进行了紫花苜蓿(第2茬)和燕麦(抽穗期)的各指标测定, 8月20日同时进行了紫花苜蓿(第4茬)和玉米(乳熟期)、甜高粱(乳熟期)的各指标测定。测定位点分别为单作禾本科带、单作紫花苜蓿带、间作禾本科带以及间作紫花苜蓿带。
1.4.2 指标测定 根系参数:于2017年3月安装CI-600植物根系监测系统(美国CID, Inc), 在每个小区中安装一根直径10 cm、长100 cm的有机透明玻璃管(简称根管), 根管与地面角为45° , 地下埋深约90 cm, 管子露出地面约10 cm, 不测量时盖上盖子, 以防灰尘、水或降水和其他物体落入管中, 同时保持管子内壁干净, 从而保证获得清晰的图片。根系扫描完成后, 用WINRHIZO TRON 2009a软件分析数据, 导出数据后整理并通过函数计算得出最终数据, 数据包括根长、根尖数、根表面积和根体积等。
碳、氮含量和碳、氮代谢关键酶:分别对4种禾本科牧草的功能叶(小黑麦和燕麦为旗叶、玉米和甜高粱为穗位叶)和紫花苜蓿功能叶[13](顶叶向下第3片完全展开的健康完整叶片)进行碳、氮含量和碳、氮代谢关键酶的测定。每小区于晴天上午9: 00-10: 00随机测定5片叶片, 重复3次。其中, 采用Somogyi法[14]测定碳水化合物含量; 采用半微量凯式定氮法[15]测定全氮含量; 采用RuBP ELISA试剂盒(上海杰美科技有限公司)测定核酮糖-1, 5-二磷酸羧化酶(RuBPCase, μ mol· g-1· min-1)活性; 采用分光光度法[14]测定蔗糖磷酸合成酶(SPS, mg· g-1· h-1)活性; 采用离体法[16]测定硝酸还原酶(NR, μ g· g-1· h-1)活性; 参照邹琦[16]的方法测定谷氨酰胺合成酶(GS, μ g· g-1· h-1)活性。
用Excel 2007软件整理试验数据后, 采用SPSS 19.0软件进行单因素方差分析(ANOVE)和Pearson相关性分析(双侧检验)。
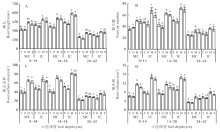
2.1.1 禾本科牧草的根系特性 从根长和根表面积来看(图1A, C), 在0~14 cm和28~42 cm土层中, 只有小黑麦和燕麦间作下显著高于单作(P< 0.05), 在14~28 cm土层中4种禾本科牧草(小黑麦、燕麦、玉米、甜高粱)间作下根长和根表面积均显著高于单作(P< 0.05); 从根尖数和根体积来看(图1B, D), 4种禾本科牧草各土层均显著高于单作(P< 0.05)。此外, 从土壤剖面来看, 根系主要分布在0~14 cm和14~28 cm, 其次是28~42 cm。
 | 图1 4种间作模式下各土层禾本科牧草的根系特性 MC:单作Monocropping; IC:间作Intercropping; T:小黑麦 Triticale; O:燕麦 Oat; M:玉米 Maize; S:甜高粱 Sweet sorghum。不同小写字母表示同一土层不同种植模式之间差异显著(P< 0.05), 下同。Fig.1 Root system characteristics of gramineae grasses under 4 kinds of intercropping patterns in different soil layers Different lowercase letters indicate significant differences between different planting modes in the same soil layer (P< 0.05), the same below. |
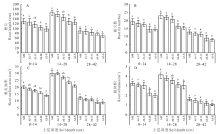
2.1.2 紫花苜蓿的根系特性 从根长、根表面积和根体积来看(图2 A, C, D), 在0~14 cm和14~28 cm土层中, 与小黑麦和燕麦间作下的紫花苜蓿与单作比无显著性差异(P> 0.05), 而与玉米和甜高粱间作下的紫花苜蓿则显著低于单作(P< 0.05), 而在28~42 cm土层中紫花苜蓿所有间作下与单作比无显著性差异(P> 0.05); 从根尖数来看(图2B), 在3个土层中, 与小黑麦和燕麦间作的紫花苜蓿与单作相比无显著性差异(P> 0.05), 而与玉米和甜高粱间作的紫花苜蓿显著低于单作(P< 0.05); 此外, 从土壤的垂直分布来看, 根系主要分布在14~28 cm范围内, 其次是0~14 cm, 最后是28~42 cm。
 | 图2 4种间作模式下各土层紫花苜蓿的根系特性 AM:紫花苜蓿单作; AI-T:与小黑麦间作的紫花苜蓿; AI-O:与燕麦间作的紫花苜蓿; AI-M:与玉米间作的紫花苜蓿; AI-S:与甜高粱间作的紫花苜蓿. 下同Fig.2 Root system characteristics of alfalfa under 4 kinds of intercropping patterns in different soil layers AM: Alfalfa monocropping; AI-T: Alfalfa intercropping with triticale; AI-O: Alfalfa intercropping with oat; AI-M: Alfalfa intercropping with maize; AI-S: Alfalfa intercropping with sweet sorghum. The same below. |
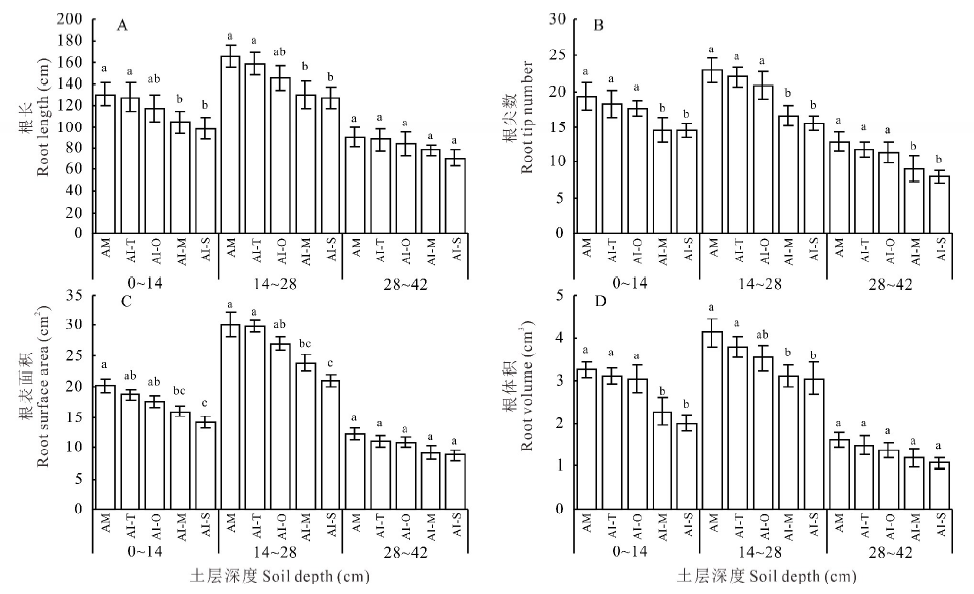
小黑麦、燕麦、玉米和甜高粱间作下RuBPCase活性与相应单作比分别增加了21.42%、20.89%、13.71%和11.79%, 均呈显著性差异(P< 0.05), 其中小黑麦和燕麦间作下RuBPCase活性的增幅最大(> 20%)(图3A)。4种禾本科牧草间作下SPS活性和碳水化合物含量与单作比均无显著性差异(P> 0.05)(图3B, C); 与小黑麦和燕麦间作下的紫花苜蓿其RuBPCase活性与单作比无显著性差异(P> 0.05), 而与玉米和甜高粱间作下分别比单作减少21.70%和24.38%, 呈显著性差异(P< 0.05)(图3D); 紫花苜蓿4种间作模式下SPS活性和碳水化合物含量均略低于单作, 但并无显著性差异(P> 0.05)(图3E, F)。
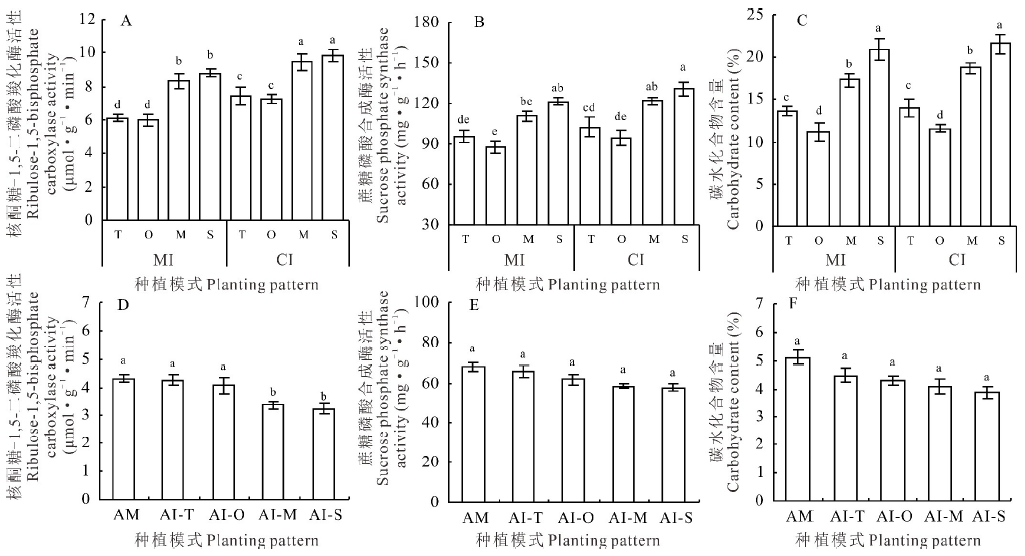 | 图3 4种间作模式下禾本科牧草和紫花苜蓿的碳代谢特征Fig.3 Carbon metabolism characteristics of gramineae grasses and alfalfa under 4 kinds of intercropping patterns |
小黑麦、燕麦、玉米和甜高粱间作下NR活性与相应单作比分别增加了24.54%、21.87%、12.87%和11.29%, 均显著高于单作(P< 0.05), 其中小黑麦和燕麦间作下NR活性的增幅最大(> 20%)(图4A); 4种禾本科牧草间作下GS活性和全氮含量均高于单作, 但无显著性差异(P> 0.05)(图4B, C)。紫花苜蓿间作下NR活性、GS活性和全氮含量均低于单作(图4D~F)。其中, 与小黑麦、燕麦间作下的紫花苜蓿与单作相比NR活性无显著性差异(P> 0.05), 而与玉米、甜高粱间作下与单作比分别降低了22.49%和25.85%, 均显著低于单作(P< 0.05); 4种间作下的紫花苜蓿GS活性和全氮含量与单作相比均无显著性差异(P> 0.05)。
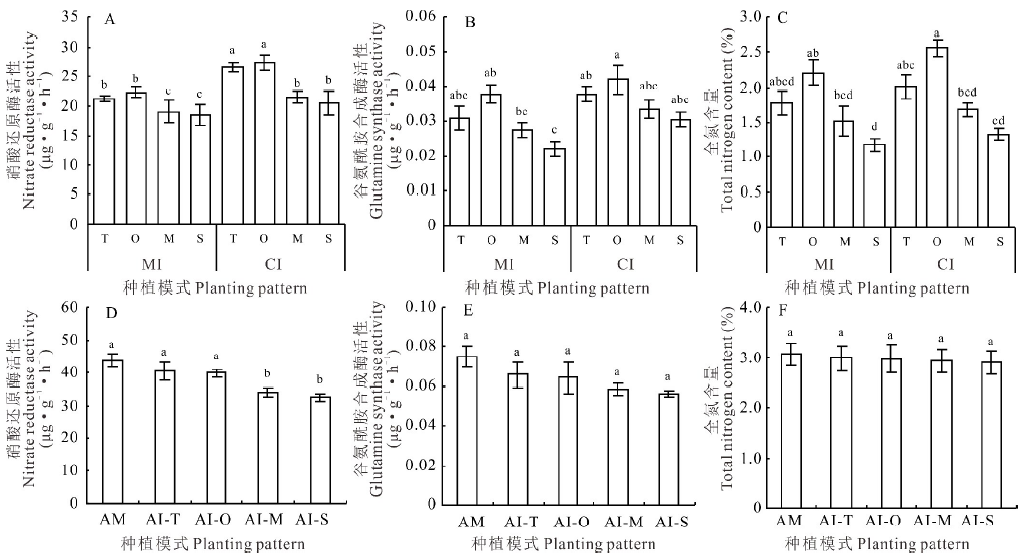 | 图4 4种间作模式下禾本科牧草和紫花苜蓿的氮代谢特征Fig.4 Nitrogen metabolism characteristics of gramineae grasses and alfalfa under 4 kinds of intercropping patterns |
根系主要参数(根长、根尖数、根表面积、根体积)与碳、氮代谢主要指标(碳水化合物、RuBPCase、SPS、全氮、NR、GS)均呈正相关关系(表2)。其中, 根长与RuBPCase和NR之间呈极显著正相关关系(P< 0.01), 相关系数分别达0.801和0.863, 与SPS和GS之间呈显著正相关关系(P< 0.05), 相关系数分别是0.707和0.730。根表面积与RuBPCase和NR之间也呈极显著正相关关系(P< 0.01), 相关系数分别达0.864和0.894, 与SPS和GS之间呈显著正相关关系(P< 0.05), 相关系数分别是0.716和0.763。
| 表2 根系特性与碳、氮代谢的相关性分析 Table 2 Correlation analysis of root system characteristics and carbon and nitrogen metabolism |
植物根系作为提供养分和水分的“ 源” , 其在介质中的延伸显著影响着地上部分干物质的积累, 地上部分代谢产物的形成和转运也同时反馈到根系的生长特性上。Tjoelker等[17]、Withington等[18]和Freschet等[19]研究表明, 在获取和分配有限的环境资源方面, 植物地上和地下部分之间存在较强的相关性。本研究中, 根长、根表面积与RuBPCase活性呈极显著正相关(P< 0.01), 与SPS活性呈显著正相关(P< 0.05), 说明5种供试牧草的根系发育状况直接影响光合碳代谢, Hou等[20]、汪顺义等[21]和文莲莲等[22]在对甘薯(Dioscorea esculenta)和番茄(Lycopersicon esculencum)的研究中也发现随着根长的增加, RuBPCase和SPS活性均显著提高。同时, 本研究中, 根长、根表面积与NR活性呈极显著正相关(P< 0.01), 与GS活性呈显著正相关(P< 0.05), 说明根长和根表面积对氮代谢也起到了积极的促进作用, 王佳等[23]和宋淑英等[24]在对小麦(Triticum aestivum)根系的研究中也证实存在此现象。由此可见, 无论是禾本科牧草还是豆科牧草, 根长和根表面积与碳、氮代谢有着直接关系, 根系的发育状况直接决定了碳、氮代谢的活跃程度。
本研究采用了微根管观测法对不同禾本科/豆科牧草间作模式下根系进行了研究, 这种方法是由Bates[25]于1937年在对黑麦草根系进行研究时最早采用的, 微根管法具有原位、定点、连续观测、结果准确等优点, 是获取根系构型和生长状态的一种有效手段, 可获取植物根系分布及生长发育状况、根长、根尖数等参数信息。近年来, 这种方法得到了根系研究者的青睐, 该技术逐渐在林地[26, 27]、农田[28, 29]和草地[30]等研究领域使用。并且在应用中不断改进, 不仅可以通过改进根管的埋设方式, 利用长、短根管相结合的埋设方式加大根管的监测范围, 还可通过调整根管的长度和直径, 使其适合大型深根植物的研究[29], 国外已经有利用超长根管(5~10 m)进行深层土壤养分和水分等的研究[31]。目前利用微根管法通过结合电子摄像技术或计算机断层成像技术等其他技术手段, 对根系的研究仍在进一步深化, 尤其在对深层土壤养分、水分及其与植物、土壤微生物等的互作领域研究中。在本研究中应用的微根管法采用1 m的根管并结合计算机成像技术进行根系原位观测和WINRHIZO TRON 2009a软件进行后期图像分析, 最终得到根系特性相关数据, 与传统的破坏性根系研究法相比, 本研究所用微根管法是在不影响植物正常生长的前提下对根系生长进行原位、无损、动态和连续的观测, 数据更为真实可靠, 是目前根系原位研究最先进的方法。
本研究中, 4种间作模式下禾本科牧草和豆科牧草的根系形态发生了改变, 禾本科牧草的根长、根表面积等根系参数明显优化, 而紫花苜蓿根系特性与单作相比表现不一, 与玉米和甜高粱间作的紫花苜蓿其根系特性受到了明显地抑制(显著低于单作), 表现为竞争弱势, 而与小黑麦和燕麦间作的紫花苜蓿则未受影响, 分析原因, 可能是因为豆科/禾本科间作下, 紫花苜蓿通过根瘤固氮及氮素转移等作用促进禾本科氮素吸收, 从而有利于禾本科牧草根系生长; 另一方面, 根的生长方式受基因控制, 具有信号识别能力, 单作下禾本科作物的根系会分布在原有区域, 当发现异己植株时, 禾本科植物为了占据更多的空间和吸收更多的养分, 便生长出更多的根系, 且根系有向豆科植物根系分布区域延伸和扩展的趋势[12], Hauggaard-Nielsen等[32]和李秋祝等[33]在豌豆(Pisum sativnm)/大麦(Hordeurn vulgare)和蚕豆(Vioia faba)/玉米间作研究中也有相似的发现, 说明间作体系中在营养吸收方面禾本科牧草较豆科牧草具有竞争优势, 4种间作模式中紫花苜蓿/小黑麦和紫花苜蓿/燕麦2种间作模式在根系特性方面表现更佳。同时, 本研究的4种间作模式下, 禾本科牧草功能叶RuBPCase和NR活性与单作比均显著性增加, 而紫花苜蓿功能叶RuBPCase和NR活性与单作相比趋势不尽相同, 其中, 与玉米和甜高粱间作的紫花苜蓿其RuBPCase和NR活性显著低于单作, 而与小黑麦和燕麦间作的紫花苜蓿则差异不显著, 唐秀梅等[34, 35]在研究中也发现木薯(Manihot esculenta)/花生(Arachis hypogaeo)间作系统中, 木薯叶片碳、氮代谢关键酶均提高, 而花生叶片则降低, 这可能与高位作物(禾本科牧草)和低位作物(紫花苜蓿)对资源的占有和利用能力不同有关[36], Keddy等[37]的研究表明, 间作下不同的作物在获取环境资源时存在竞争作用, 通常叶面积较大及叶片位置较高的植物在竞争环境资源时处于竞争优势地位, 因而具有竞争优势的作物代谢更为活跃, 由此可见, 间作体系中禾本科牧草的碳、氮代谢更加活跃与旺盛, 并且紫花苜蓿/小黑麦和紫花苜蓿/燕麦这2种间作模式在碳、氮代谢方面表现更佳。此外, 通过相关性分析发现, 在本研究的单作和间作模式下, 根系参数与碳、氮代谢各指标均呈正相关关系, 表明根系特性与碳、氮代谢具有一致性。
本研究中, 供试牧草根系特性和碳、氮代谢特征表现趋势一致, 在4种间作模式下均表现为禾本科牧草优于豆科牧草; 紫花苜蓿/小黑麦和紫花苜蓿/燕麦这2种间作模式下综合表现最佳。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|