



{kind=link}
{kind=link}
{kind=link}
围栏封育对温性草甸草原植物群落构成及生态位特征的影响
[聂莹莹 , 徐丽君
, 徐丽君* , 辛晓平, 陈宝瑞, 张保辉]
, 徐丽君, 辛晓平, 陈宝瑞, 张保辉]
|
|


作者简介:聂莹莹(1990-),女,科研助理,硕士。E-mail: 547105832@qq.com
为明确温性草甸草原群落构成和生态位对围栏封育的响应,运用了Levins生态位指数和Pianka生态位重叠指数以及物种多样性指数分析围栏内外植物种对资源的利用情况和种间竞争情况。结果表明:围栏封育显著促进退化草地植被恢复,植被地上生物量和地下生物量分别较对照提高255.06%,51.06%;物种多样性与丰富度分别增加了10.36%,10.34%。围封样地内生态位宽度排名前2位的植物为囊花鸢尾和羊草,生态位宽度值分别为0.921和0.873;自由放牧样地生态位宽度排在前2的植物为蒲公英和寸草苔,生态位宽度值依次为0.912和0.791。围栏封育可以显著提高草地生产力,并且对物种多样性也有一定的促进作用,是应对退化温性草甸草原恢复的有效措施;与自由放牧相比,围栏封育增加了群落植物种整体生态位重叠值和种间竞争。
This study explored the effects of fence enclosure on community composition and niche characteristics in temperate meadow steppe. The indexes Levins niche breadth, Pianka niche overlap and species diversity were used to evaluate the degree of resource utilization by species and the intensity of interspecific competition inside and outside the enclosure. It was found that: the fencing enclosure significantly promoted vegetation restoration of degraded grassland. The aboveground biomass and belowground biomass in the enclosure treatments were enhanced by 255.06% and 51.06%, respectively. The species diversity and richness were enhanced in fenced enclosures by 10.36% and 10.34%, respectively. The two species with the highest niche breadth in enclosures were Iris ventricosa and Leymus chinensis, with values of 0.921 and 0.873, respectively. In comparison, the two species with the highest niche breadth in the freely grazed grassland were Taraxacum mongolicum and Carex duriuscula, with values of 0.912 and 0.791, respectively. Our results clearly show that enclosure by fencing is an efficient restoration measure that not only promotes species diversity but also improves grassland productivity. Compared with free grazing, fencing enclosure increases the overall niche overlap value and inter-species competition of plant communities and their component plant populations.
呼伦贝尔是我国温带草甸草原分布最集中、最具代表性的地区, 是我国目前原生植被保存最好、景观生态类型和生物多样性最丰富的草原生态系统类型[1]。近些年, 随着生产发展和人口增加, 人们对草地的掠夺逐渐加剧, 导致呼伦贝尔草原由点状退化已经发展成了区域性的整体退化[2, 3], 这不仅阻碍了当地草地畜牧业的发展, 也对草甸草原物种多样性保护和生态环境维持构成严重威胁。因此, 草地生态系统的管理是诸多学者关注的热点。围栏封育作为一种有效、简便易行的促进退化草地生态恢复的策略[4], 不仅能够促进物种多样性[5], 提高退化草地生产力[6], 而且还能改善土壤的理化性质[7]。围栏封育是当前退化草地恢复与重建的重要举措之一, 在一定的恢复时间尺度内可提高物种丰富度, 促进物种对资源利用能力和生态位功能的发挥[8]。
生态位概念自1917年Grinnell提出以来, 受到国内外众多生态学者的关注, 并对该理论进行了广泛研究, 使得该理论在植物种间关系、生物多样性、群落结构和演替以及种群进化等方面得到广泛应用[9]。研究认为植物利用资源的状况反映了植物种群间相互关系[10], 因此资源利用状况是认识群落组成形成机制的主要问题, 而生态位理论可以揭示物种间对可利用资源的量化问题[11]。经典的生态位理论认为, 物种在某种(或多种)环境资源利用上存在差异, 即物种间的生态位分化, 这是物种共存的基本机制之一[12], 也是物种进化、群落结构变化和群落演替的主要动力和群落动态研究的重点[13]。关于生态位理论的研究目前已经较为成熟, 尤其对于生态位宽度与生态位重叠的测度[14]。研究种群生态位, 了解各种群在群落中的地位和作用以及种群间的相互关系, 对植被资源保护、可持续利用和植被的恢复重建等具有重要意义。
近年来, 众多学者在草甸草原上开展了大量的工作, 研究内容主要集中于封育下草甸草原群落结构与物种多样性[15]、功能群结构与生物量[16]、土壤理化性质[17]、土壤种子库[18]、种群空间分布格局[19]和群落稳定性[20]等方面。但针对围栏封育和自由放牧两种干扰方式下的植物群落构成和种群生态位特征的研究尚少见相关报道。因此, 本研究基于经典的生态位理论, 通过调查温性草甸草原植物群落数量特征, 分析在围栏封育和自由放牧两种干扰方式下草原植物群落构成、生态位宽度以及生态位重叠, 探索退化草甸草原植物群落的种群资源利用状况及生态适应能力, 以期为呼伦贝尔温性草甸草原植物多样性维持机制与植被恢复提供参考依据。
研究区位于内蒙古呼伦贝尔草原生态系统观测研究站(北纬49° 19', 东经119° 56'), 温带大陆性季风气候。1月平均气温为-48.5 ℃, 7月平均气温为36.2 ℃, 年平均气温为1.5 ℃。≥ 10 ℃的活动积温在1700~2300 ℃, 无霜期约为110 d。年平均降水量350~400 mm, 主要集中在6-9月。草甸草原建群种为贝加尔针茅(Stipa baicalensis), 优势种为羊草(Leymus chinensis), 常见种有糙隐子草(Cleistogenes squarrosa)、麻花头(Serratula komarovii)、扁蓿豆(Medicago ruthenica)、寸草苔(Carex duriuscula)、裂叶蒿(Artemisia laciniata)等, 并伴以其他杂类草。土壤类型主要为淡栗钙土。
试验地为呼伦贝尔羊草草甸草原围封样地, 围封试验始于2006年, 将2006年作为封育第1年, 围栏外的自由放牧地作为对照样地, 于2006-2019年每年7月初植物生长最旺盛的季节进行取样(本试验采用2013年即封育8年的监测数据), 采用样方法在围栏内外随机进行取样测定植物高度、盖度、密度、地上生物量和地下生物量指标, 每个样方面积为1 m× 1 m, 围栏内取10个, 围栏外取10个, 共计样方20个。在围栏内外分别选取重要值大于0.2的物种进行生态位分析。
在每个样方内测量如下指标[4]:盖度:采用针刺法测定盖度。高度:每种植物随机5株的平均值。密度:同一植物出现的个数(或丛数)。地上生物量:将样方内的植物分种齐地面剪下, 装入塑料样品袋中, 编号带回实验室。将每个样方中的地上生物量分为地上绿色生物量和枯落物(包括立枯物和凋落物)两部分, 称取鲜重, 并装入纸袋中在恒温(85 ℃)下烘干12 h, 称其干重。地下生物量:在每个样方内用直径10 cm的根钻, 按照0~10 cm、10~20 cm、20~30 cm、30~40 cm、40~50 cm、50~60 cm的深度钻取土柱, 每个样方重复取6钻。将土柱放在0.5 mm网眼的纱网袋里, 用水冲洗, 将采集到的根系置于85 ℃的烘箱中烘干称重, 计算地下生物量。
1.4.1 重要值计算
IV=(RC+RH+RD+RB)/4× 100
式中:IV表示重要值, RC为相对盖度, RH为相对高度, RD为相对密度, RB为相对地上生物量; 相对盖度为某一种植物盖度占全部植物盖度总和的百分比; 相对高度为某一种植物高度占全部植物高度总和的百分比; 相对密度为某一种植物密度占全部植物密度总和的百分比; 相对地上生物量为某一种植物地上生物量占全部植物地上生物量总和的百分比。
1.4.2 物种多样性计算 选用Shannon-Wiener多样性指数、Simpson多样性指数、Pielou均匀度指数和Margalef丰富度指数进行多样性测度, 计算公式如下:
Shannon-Wiener指数
Simpson指数
Pielou均匀度指数 J=H/lnS
Margalef丰富度指数 M=
式中:Pi为样方内的物种i的相对重要值, Pi=Ni/N(Ni为物种i的绝对重要值, N为物种i所在样方的各个植物种的绝对重要值总和), S为样方内的物种数量, N为样方内的全部植株数量。
1.4.3 生态位宽度 采用以下公式[21]计算:
式中:Bj表示第j个物种的生态位宽度, Pjk表示第j个物种在第k个资源水平下的重要值占该种在所有资源水平上的重要值总和的比例, t表示样方总数。
1.4.4 生态位重叠公式[22] 计算如下:
式中:Oij为物种i和j的生态位重叠值, Pik和Pjk分别表示第i和j个物种在第k个样方的重要值占该种在所有样方中重要值总和的比例, t为样方总数。
样地全部物种间生态位重叠值的总平均值=样地内全部物种间生态位重叠值总数/总种对数
采用Excel 2013软件进行数据整理与作图, 运用SPSS 20.0软件进行数据分析, 围栏内外物种多样性与生物量差异显著性采用单因素分析(One-Way ANOVA), 并用Duncan检验进行平均值之间的多重比较, 显著性水平设为P< 0.05。
围栏封育提高了各土层的植物根系生物量(图1A), 其中, 0~10 cm土层差异显著(P< 0.05), 相较于围栏外增加44.98%, 其他土层差异不显著(P> 0.05); 这说明了自由放牧对表层土壤中根系生物量的影响较大。
 | 图1 围栏内外群落地下生物量和地上生物量 * 表示差异显著(P< 0.05), 无* 表示差异不显著, 下同。Fig.1 The belowground biomass and aboveground biomass of plant community inside and outside the enclosure * indicates a significant difference at 0.05 level, no * indicates the difference is not significant, the same below. |
围栏封育能够显著增加地上生物量(P< 0.05), 与围栏外的自由放牧样地相比, 地上生物量增加了255.07%(图1B)。
物种多样性是群落的重要特征, 围栏封育增加了Shannon-Wiener多样性指数、Simpson多样性指数、Pielou均匀度指数和Margalef丰富度指数值, 与围栏外的自由放牧相比, 分别增加了10.36%、10.19%、13.89%和10.34%(图2); 围栏封育增加草地植物群落物种多样性和丰富度, 但差异均未达到显著性水平(P> 0.05)。
 | 图2 围栏内外草地植物群落多样性指数、均匀度指数和物种丰富度指数Fig.2 The diversity, evenness and richness indexes of plant species inside and outside the enclosure |
2.3.1 重要值 围栏内外主要植物种的重要值有明显的差异(表1)。围栏封育样地的羊草、裂叶蒿、麻花头、日荫菅(Carex pediformis)、囊花鸢尾(Iris ventricosa)、叉枝鸦葱(Scorzonera divaricata)、狭叶青蒿(Artemisia dracunculus)、二裂委陵菜(Potentilla bifurca)和沙参(Adenophora stricta)的重要值高于围栏外的自由放牧样地, 分别高出213.53%、313.78%、138.59%、412.52%、161.07%、22.20%、66.93%、62.47%和2181.00%。围栏外自由放牧样地内的贝加尔针茅、糙隐子草、冷蒿(Artemisia frigida)、寸草苔、羽茅(Achnatherum sibiricum)和披针叶黄华(Thermopsis lanceolata)的重要值比围栏内高出101.23%、1759.75%、209.51%、696.40%、40.09%和29.75%。
| 表1 围栏内外主要植物种生态位宽度与重要值 Table 1 Niche breadth and importance value of main species inside and outside the enclosure |
2.3.2 生态位宽度 生态位宽度反映了植物种群对资源的利用及对环境的适应能力, 表征了种群在群落中的竞争地位, 种群生态位越宽, 表明在群落中竞争能力越强, 种群生态位越窄, 表明在资源环境的竞争中处于劣势地位[23]。封育样地内生态位宽度较大的物种有囊花鸢尾、羊草、日荫菅、沙参、麻花头和展枝唐松草(Thalictrum squarrosum), 其中生态位宽度值排在前3位的植物为囊花鸢尾、羊草和日荫菅; 此3种物种出现在调查的每一个样方中, 其中囊花鸢尾属于多年生密丛型草本植物, 且耐盐碱, 耐寒, 占据了最多的资源, 生态位宽度值最大, 为0.921, 羊草和日荫菅次之, 分别为 0.873和 0.852(表1)。而生态位宽度排在后3位的植物是阿尔泰狗娃花(Heteropappus altaicuc)(0.200), 羽茅(0.199)和披针叶黄华(0.163)。3种物种在所有调查的样方中出现的频率极低, 为偶见种, 说明此3种植物具有较低的资源利用谱, 对资源的利用处于劣势地位, 对环境的适应能力相对较差。
围栏外的自由放牧样地生态位宽度较大的物种有蒲公英(Taraxacum mongolicum)、寸草苔、斜茎黄芪(Astragalus adsurgens)、独行菜(Lepidium apetalum)、麻花头、羊草和贝加尔针茅; 生态位宽度排名前3位的物种是蒲公英、寸草苔和斜茎黄芪, 其值分别为0.912、0.791和0.624(表1)。生态位宽度排在后3位的物种为披针叶黄华、冷蒿和日荫菅, 值分别为0.149、0.100和0.100, 在调查的10个样方中, 出现的频率低, 生态位宽度较窄, 说明其对空间的侵占能力较差, 是此群落中易于退化的种群。
综合两种干扰方式下的样地, 其中封育样地的羊草、针茅、麻花头和囊花鸢尾等11种植物的生态位宽度高于围栏外的自由放牧样地, 其中, 群落中的优势种羊草的生态位宽度值比围栏外高出63.48%。围栏外的羽茅、糙隐子草、二裂委陵菜和菊叶委陵菜(Potentilla tanacetiflolia)等6种植物的生态位宽度值高于围栏内(表1)。导致这种现象出现的原因可能是自由放牧区的牲畜的选择性采食, 践踏等使适口性良好、不耐践踏的植物种群数量下降, 生态位宽度降低。而封育却促使这部分植物种群数量增加, 生态位宽度随之增加。
通过对调查样地植物群落中各植物的生态位宽度与重要值之间的回归分析, 围栏封育下的各植物种生态位宽度与重要值之间的相关关系的方程为:y=0.0259x+0.3724, R2=0.4203, P< 0.01; 围栏外的自由放牧样地各植物种生态位宽度与重要值之间建立的关系为:y=0.0152x+0.3298, R2=0.2678, P< 0.01(图3)。二者均呈现正相关关系, 且解释变量R2值大小为围栏内> 围栏外, 说明围栏内生态位宽度与重要值的相关关系大于围栏外。一般来说, 重要值越大的物种, 其生态位宽度往往也越大, 对资源的利用和对环境的适应能力也越强。但也有研究认为, 重要值大小并非是生态位宽度大小唯一的因素, 也就是说重要值最大的物种其生态位宽度也并非最大[13]。本研究中也存在个别植物种重要值较大, 但是生态位宽度值并不是最大的。这可能与植物种群自身特性制约, 或是土壤湿度与不同干扰因子影响息息相关。
 | 图3 围栏内外生态位宽度与重要值的相关关系Fig.3 Correlation between niche breadth and important value inside and outside the enclosure |
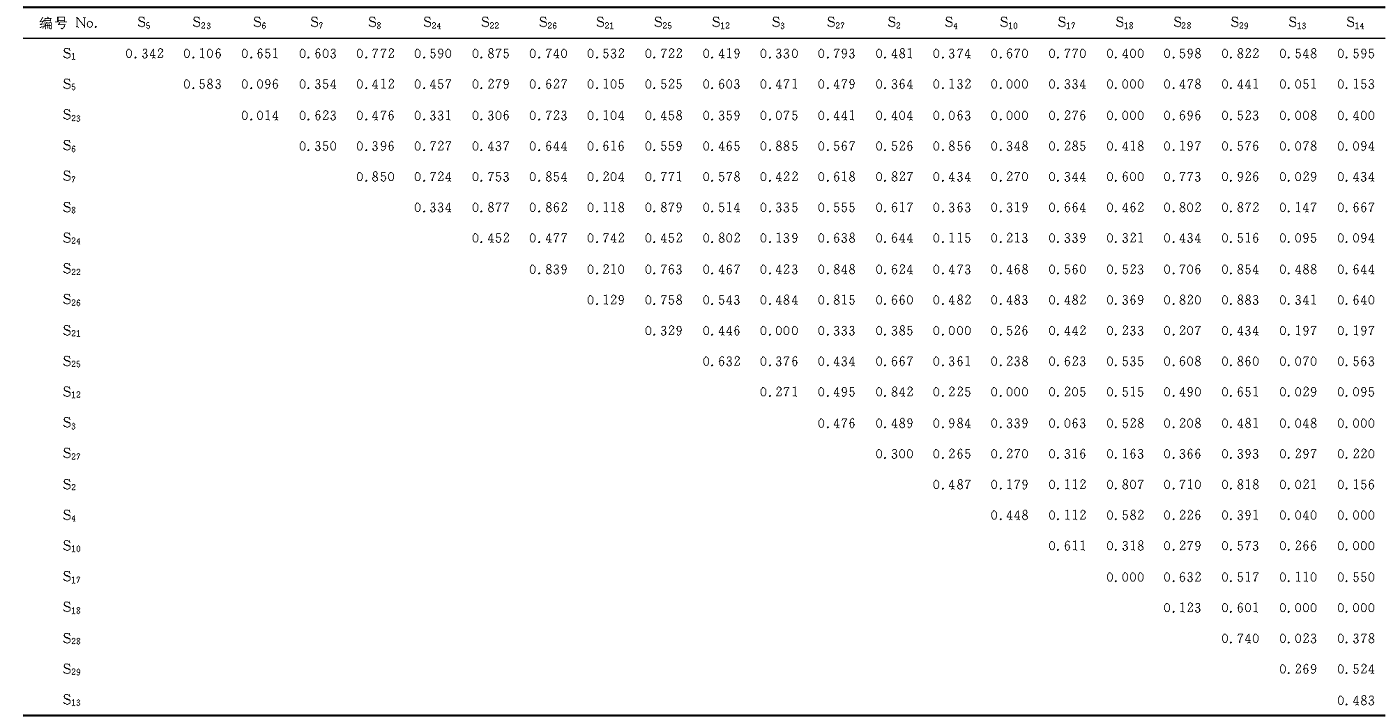
生态位重叠体现了种群之间对于相同等级资源利用的相似程度和空间上的配置关系, 较高的生态位重叠代表了种群之间对环境资源具有相似的生态学要求, 因此可能会存在激烈的竞争[24]。 从表2可以看出, 围栏内生态位重叠值为0.000的种对有13种, 占总种对数的5.14%; 生态位重叠值在0.000~0.300的种对有61对, 占总种对数的24.11%。生态位重叠值在0.300~0.850的有165种对, 占总种对数的65.22%; 生态位重叠值大于0.850的有14对种对, 占总种对数的5.53%, 分别为羊草和日荫菅(0.875)、冷蒿和羽茅(0.885)、冷蒿和糙隐子草(0.856)、裂叶蒿和麻花头(0.850)、沙参和裂叶蒿(0.854)、囊花鸢尾和裂叶蒿(0.926)、麻花头和日荫菅(0.877)、 沙参和麻花头(0.862)、 细叶白头翁和麻花头(0.879)、 囊花鸢尾和麻花头(0.872)、日荫菅和囊花鸢尾(0.854)、沙参和囊花鸢尾(0.883), 细叶白头翁和囊花鸢尾(0.860)、羽茅和糙隐子草(0.984)。生态位宽度最大的囊花鸢尾(0.921)与其他物种之间的重叠值均较高。生态位宽度均较小的羽茅(0.199)和糙隐子草(0.224)有着最高的重叠, 但与其他大多数物种的重叠值较低。生态位重叠值大, 说明两物种对资源的利用方式相似或对生境的要求基本一致。
| 表2 围栏内主要植物种生态位重叠 Table 2 Niche overlap of main plant species inside of enclosure |
由表3可知, 围栏外的自由放牧样地内有24对种对的生态位重叠值为0.000, 占总种对数8.00%; 生态位重叠值在0.000~0.300共有123对种对, 占总种对数的41.00%; 生态位重叠值在0.300~0.850共有143对种对, 占总种对数的47.67%; 生态位重叠值大于0.850的有10种对, 占总种对数的3.33%, 分别为羊草和羽茅(0.873)、裂叶蒿和羽茅(0.900)、狭叶青蒿和轮叶委陵菜(0.918)、麻花头和二裂委陵菜(0.980)、狭叶青蒿和针茅(0.861)、针茅和糙隐子草(0.861)、叉枝鸦葱和囊花鸢尾(0.899)、二裂委陵菜和菊叶委陵菜(0.978)、扁蓿豆和车前(0.942)。生态位重叠最大值出现在麻花头和二裂委陵菜之间, 说明两个物种对环境资源具有相似的生态学要求。
| 表3 围栏外主要植物种生态位重叠 Table 3 Niche overlap of main plant species outside of encl osure |
在围栏内外生态位重叠值为0的植物种对, 其对资源的利用方式具有较大差异, 或者分布具有局限性, 造成明显的生态位分化。在调查的10个样方中, 这些物种的分布区域有限, 而且出现频率极低, 仅在1~2个样方中出现, 导致与其他植物共存的概率低, 所以出现生态位重叠值为0的情况。由表4可以得到, 围栏封育增加了群落整体生态位重叠值, 与对照相比, 生态位重叠指数平均值增加了25.57%。说明围栏封育为植物提供了更好的生存空间, 使种群间利用资源接近, 进而加剧了植物种间的竞争。围栏外的自由放牧区, 牲畜的啃食与践踏, 促使生态位分化, 降低了资源的利用和物种间的竞争。
| 表4 围栏内外所有物种生态位重叠指数平均值 Table 4 Average niche overlap index of all the species inside and outside the enclosure |
物种多样性是维持草地生态系统稳定和生产力的基础[25]; 生物量则是生态系统生产力的重要体现, 更是整个生态系统运行的能量基础和营养物质来源[26]。前人的研究中关于植物多样性对封育时间的响应并未得到一致性的结论。有研究指出, 围栏封育增加了物种多样性[27], 也有研究表明围栏封育使物种多样性降低[28], 还有研究指出封育对物种多样性没有显著性影响[29]。导致这些结论不同的主要因素包括封育前草地退化程度、草地类型或生境、封育时间以及放牧强度等差异[30]。本研究中围栏封育样地与围栏外自由放牧样地相比, 围栏封育显著增加了群落地上生物量和地下生物量; 同时也增加了物种多样性和物种丰富度, 与左万庆等[31]围栏封育措施对退化羊草草原植物群落特征影响的结果一致。围栏封育是恢复退化草地的有效措施, 可以改善生境, 为植物群落中减少或消退的物种再次入侵创造了条件。
生态位宽度和生态位重叠是影响物种多样性及群落结构的决定因素, 都在一定程度上反映了植物自身的特性和对环境的适应性, 二者既有联系, 又有区别[32]。生态位宽度是衡量植物种对环境资源利用状况的一个指标, 反映某一种群对环境的适应性, 其值大小表示占有资源的强弱[33]; 生态位重叠表示种群间对资源的共享能力及环境资源的利用状况, 反映种群间的竞争关系和分布地段的交错程度, 生态位重叠值的大小表示植物利用资源方式的相似性[34]。相关研究表明, 生态位宽度越大, 对环境的适应能力越强, 同时伴随着较高的生态位重叠, 生态位宽度较小的物种则因对资源的利用能力较弱, 分布较窄而与其他物种间有较低的生态位重叠, 但由于环境资源存在高度的空间异质性或者不同演替阶段的竞争特点存在差异, 较高的生态位重叠值也会出现在生态位宽度较小的物种间[35]。
种群生态位除受自身特性制约外, 群落海拔、土壤湿度和人为干扰等主要环境因子对种群生态位也有重要影响[36]。目前, 围栏封育和放牧是人类在草地生态系统管理利用实践中的主要干扰类型, 对生态系统过程产生重要影响。本研究结果显示, 围栏封育样地内生态位排在前2位的植物为囊花鸢尾和羊草, 其中囊花鸢尾是密丛型草本植物, 对资源有着较强的利用能力, 羊草为群落中的优势种, 优势种在创建植物群落内部独特的环境和决定群落种类组成方面有着重要作用[24]。放牧由于牲畜的选择性采食和践踏改变草地植物种群分布, 从而影响着草地种群生态位特征[8]。自由放牧样地内的一些适口性好的高大型牧草生态位降低, 从而为牲畜不喜食或是耐牧强、耐践踏的低矮型植物种的生存拓宽了空间, 增强了对资源的利用力; 本研究中的蒲公英, 因其极强的生命力、耐贫瘠等特性, 在自由放牧样地内表现出最大的生态位宽度值; 寸草苔为下繁草, 耐践踏, 繁殖能力强, 其生态位宽度值次之。研究还发现围栏封育样地内的植物种生态位重叠值大于0.850和处于0.300~0.850的种对分别比围栏外的自由放牧样地增加1.80%和17.88%。由此可见, 围栏封育避免外界干扰, 为生境提供了一个休养生息的机会, 使植物种群间利用资源接近, 进而加剧了植物种间竞争。
1)围栏封育相较于自由放牧可以显著提高草地生产力, 对物种多样性也有一定的促进作用, 是应对退化温性草甸草原恢复的有效措施。
2)与自由放牧比, 围栏封育显著增加了草地优势种羊草的生态位宽度(增幅为63.39%)。
3)围栏封育相较于自由放牧增加了植物群落内物种整体生态位重叠值和植物种间竞争。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|