{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
复合乳酸菌对全株玉米青贮及有氧暴露后微生物及饲料品质的影响
[万学瑞1  , 豆思远
, 豆思远2 , 李玉3 , 何轶群1 , 王川1 , 张小丽1 , 雷赵民4, * ]
, 豆思远]
|
|


作者简介:万学瑞(1979-),女,甘肃白银人,副教授,博士。 E-mail: 383921499@qq.com
为探讨不同复合乳酸菌对全株玉米青贮及有氧暴露后青贮饲料中微生物数量及其发酵品质的影响,进一步筛选出可提高青贮饲料品质和有氧稳定性的复合乳酸菌接种剂,共设置3个不同组合乳酸菌分别添加全株玉米进行青贮,并设空白对照,测定青贮3、7、15和30 d,开袋有氧暴露1、3、7、15和30 d时微生物数量、发酵品质和营养成分的变化情况。结果表明,分别添加异型乳酸菌组合(Hetero)、同型乳酸菌组合(Homo)和同型+异型乳酸菌组合(Homo+Hetero)的各处理组均能有效地增加青贮过程和有氧暴露后饲料中乳酸菌的数量、乳酸(LA)和乙酸(AA)的含量,减少好氧细菌、酵母菌和霉菌数量,降低pH和氨态氮(NH3-N)含量,减少粗蛋白(CP)损失,显著改善青贮饲料发酵品质,抑制青贮饲料有氧暴露后的二次发酵,其中Homo+Hetero效果最好,Homo处理组好于Hetero处理组。
This research investigated the effect of lactic acid bacteria (LAB) preparations on microbial population counts and silage quality of whole maize silage during fermentation and on aerobic exposure after fermentation. The aim was to test if inoculation with LAB preparations can improve silage quality and aerobic stability. Three different LAB preparations [Homofermentative (Homo), Heterofermentative (Hetero), combined Homo+Hetero] were added to whole maize during ensiling. A blank control treatment was also included. Data on microbial population count, fermentation quality and nutrient contents of silage were evaluated during fermentation at 3, 7, 15, and 30 days after ensiling, and on completion of fermentation, after 1, 3, 7, 15 and 30 days of aerobic exposure. It was found that both during maize silage fermentation and during subsequent aerobic exposure, the LAB population counts, the lactic and acetic acid concentrations were significantly increased compared with the blank control treatment, while the numbers of aerobic bacteria, yeasts and molds and crude protein loss were significantly reduced and the concentrations of NH3-N and the pH were obviously reduced by addition of the three LAB preparations. These results indicate that LAB preparations improve maize silage nutritional quality and inhibit secondary fermentation after aerobic exposure. The combination treatment Homo+Hetero performed best, and Homo performed better than Hetero.
青贮添加剂的研发和推广应用已成为目前青贮饲料的研究重点和热点, 在规范的青贮制作技术和科学的管理前提下, 利用青贮添加剂可有效调控青贮发酵进程, 改善青贮饲料品质[1, 2]。其中, 乳酸菌作为环保而有效的青贮促进剂越来越受到人们的关注。研究表明, 同型发酵乳酸菌可在青贮发酵过程中快速、有效的产生乳酸, 迅速降低青贮饲料的pH而抑制有害微生物的活动, 减少干物质和粗蛋白的损失, 进而改善青贮饲料的发酵品质[3]; 异型发酵乳酸菌如布氏杆菌虽然积累乳酸的能力不如同型发酵乳酸菌, 但其除了能产生乳酸, 还能产生挥发性短链脂肪酸如乙酸等, 能有效抑制需氧性微生物如酵母菌和霉菌的生长, 可抑制青贮饲料的二次发酵, 提高有氧稳定性, 减少青贮饲料在饲喂过程中的营养损耗[4, 5, 6]。同时也有研究发现, 单一的依靠同型发酵乳酸菌调节玉米秸秆青贮, 不能有效限制青贮开窖后酵母菌和霉菌等腐败菌的繁殖, 导致青贮饲料的有氧稳定性降低[7]; 而异型发酵乳酸菌与同型发酵乳酸菌相比生长较为缓慢, 需要青贮较长时间才能改善青贮饲料的有氧稳定性[8]。将不同类型的乳酸菌组合构成复合乳酸菌作为青贮接种剂, 旨在发挥不同类型发酵乳酸菌的优势, 互相弥补不足, 更好地提高发酵效率[9, 10, 11]。
为了开发适合甘肃地区全株玉米青贮的乳酸菌接种剂, 本实验室前期将从甘肃各地玉米秸秆青贮饲料中分离获得的肠膜明串珠菌肠膜亚种(B1-7)、植物乳杆菌(B3-1)、屎肠球菌(B5-2)、戊糖片球菌(B2-3)和发酵乳杆菌(E2-3)分别作为接种剂, 青贮全株玉米, 结果表明5株乳酸菌尤其是植物乳杆菌B3-1能显著改善青贮饲料发酵品质[12, 13]。本研究采用以上5株乳酸菌组合成不同的复合乳酸菌群开展全株玉米青贮, 跟踪分析青贮过程不同阶段青贮饲料中微生物的数量、pH、营养成分、有机酸和氨态氮的动态变化, 综合评价接种的复合乳酸菌对全株玉米青贮的调控效果, 进而为开发用于全株玉米青贮的复合乳酸菌接种剂提供参考依据。
供试乳酸菌:屎肠球菌(Enterococcus faecium)B5-2、肠膜明串珠菌肠膜亚种(Leuconostoc mesenteroides subsp. Mesenteroides)B1-7、发酵乳杆菌(Lactobacillus fermentum)E2-3、植物乳杆菌(Lactobacillus plantarum)B3-1、戊糖片球菌(Pediococcus pentosaceu)B2-3均由本实验室分离鉴定并保存[14]。
2012年9月16日, 自甘肃省临洮县收割青贮用全株玉米, 切成2~4 cm混匀即为本次试验青贮原料, 利用微波炉快速干燥, 以检测其含水量, 当确定青贮原料含水量降至70%左右进行青贮。
试验设4个处理组:同型发酵乳酸菌混合组(Homo):屎肠球菌B5-2、戊糖片球菌B2-3、植物乳杆菌B3-1; 异型发酵乳酸菌混合组(Hetero):发酵乳杆菌E2-3、肠膜明串珠菌肠膜亚种B1-7; 同型+异型发酵乳酸菌混合组(Homo+Hetero):植物乳杆菌B3-1、肠系膜明串珠菌肠膜亚种B1-7、发酵乳杆菌E2-3、屎肠球菌B5-2、戊糖片球菌B2-3; 对照组(CK):MRS液体培养基[12]。各供试乳酸菌分别用MSB液体培养基[12]培养至对数生长期, 调菌浓度为1× 109 CFU· mL-1, 等比例混合制备各处理组接种所用乳酸菌添加剂。按5 mL· kg-1青贮原料添加, 即5× 106 CFU· g-1。
用灭菌喷壶将菌液均匀喷洒于切碎的全株玉米秸秆上, 混合均匀。按每袋500 g装入聚乙烯袋中压实, 真空封口机抽气封口, 每个处理组和对照组30个重复。将密封好的青贮袋放置在恒温环境(20 ℃)中发酵, 分别在青贮的第3、7、15、30天取样, 每次每个处理随机取3袋; 并于青贮第30天时将剩余青贮袋打开暴露于空气中, 分别在开袋后的第1、3、7、15、30 天取样, 每次每处理随机取3袋, 进行青贮饲料的微生物计数及pH测定。
采用普通琼脂平板进行细菌计数, 将接种好的培养基平板放置于37 ℃恒温培养箱中培养24 h计数菌落数; 采用MRS平板进行乳酸菌计数[14], 将接种好的培养基平板放置于37 ℃恒温培养箱中培养3 d后计数菌落数; 采用孟加拉红琼脂平板进行酵母菌计数, 采用葡萄糖麦芽浸膏琼脂平板进行霉菌计数, 接种好后放置于25 ℃恒温培养箱中培养3 d后计数菌落数[15]。采用酸度计法测定全株玉米青贮饲料pH[12]。
将采集的青贮饲料样品干燥、粉碎过0.425 mm筛[12], 测定其可溶性碳水化合物(water soluble carbohydrates, WSC)、中性洗涤纤维(neutral detergent fiber, NDF)、粗蛋白(crude protein, CP)和酸性洗涤纤维(acid detergent fiber, ADF)含量[15]。
采用高效液相色谱法[16]分析青贮饲料的丙酸(propionic acid, PA)、乳酸(lactic acid, LA)、乙酸(acetic acid, AA)和丁酸(butyric acid, BA)含量; 采用苯酚-次氯酸钠比色法[16]分析氨态氮(ammonia nitrogen, NH3-N)含量。
采用SPSS 18.0软件进行方差分析, 并用Duncan法对平均值进行多重比较。
复合乳酸菌对全株玉米青贮期及有氧暴露期微生物数量的影响见表1, 青贮初期各组乳酸菌数量快速增加, 到第7天时达到最大, 之后逐渐减少, 有氧暴露后继续减少, 于第30天时各处理组乳酸菌数量减少至105数量级、对照组为103数量级。在整个青贮期及有氧暴露期, 各处理组乳酸菌数量始终高于对照组且差异显著(P< 0.05), 其中青贮期初期Hetero处理组的乳酸菌数量最高, 7 d后Homo+Hetero和Homo处理组较Hetero处理组高。
| 表1 复合乳酸菌对全株玉米青贮期及有氧暴露期微生物数量的影响 Table 1 Effect of compound lactic acid bacteria on microorganism quantity in whole corn silage and aerobic process (CFU • g_1) |
各处理组和对照组的好氧细菌数量从青贮第3天开始逐渐减少, 至青贮第30天时最少, 有氧暴露后逐渐增加, Homo+Hetero处理组于第3天时最多, 其余各组于第7天时最多, 达到107数量级, 之后又缓慢减少。在整个青贮及有氧暴露期, 各处理组好氧细菌数均显著少于对照组(P< 0.05), 其中Homo+Hetero处理组最少(表1)。
各处理组和对照组的酵母菌数量从青贮第3天开始逐渐减少, 至青贮第30天均小于1× 102 CFU· g-1, 有氧暴露后均逐渐增加, Homo和Hetero处理组于第15天后减少, Homo+Hetero处理组于第7天后减少, 而对照组持续增加于第30天时最大, 达到106数量级, 而Homo和Homo+Hetero处理组只有105数量级。从青贮第7天开始至有氧暴露第30天, 各处理组酵母菌数量均显著少于对照组(P< 0.05), 青贮期各处理组间无显著差异, 但有氧暴露期, Homo和Homo+Hetero处理组间虽无显著差异, 却显著低于Hetero处理组(P< 0.05)(表1)。
各处理组和对照组的霉菌数量随着青贮发酵的进行逐渐减少, 从青贮第7天开始至有氧暴露后最初7 d, 各组的霉菌数量均小于1× 102 CFU· g-1, 之后快速增加, 至有氧暴露第30天时对照组霉菌数量达到2.48× 108 CFU· g-1, 显著高于各处理组(P< 0.05), Homo+Hetero处理组显著低于其他处理组(P< 0.05)(表1)。
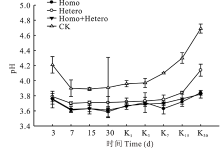
由图1可知, 随着青贮的进行各处理组和对照组的pH均下降, 青贮第3天时除对照组的pH为4.21以外, 各处理组均降至3.8以下; 对照组于第7天时降至3.9, 各处理组和对照组pH自此至青贮第30天一直保持相对稳定, 在整个青贮发酵过程中, 各处理组pH均低于对照组且差异显著(P< 0.05), 但各处理组间无显著差异。有氧暴露后各组pH均有所升高, Homo和Homo+Hetero处理组pH上升较为缓慢, 第30天时分别升至3.84和3.82, Hetero处理组为4.14, 而对照上升最快, 达到4.69, 在整个有氧暴露期间, 各处理组pH均显著低于对照组(P< 0.05), Homo和Homo+Hetero处理组间无显著差异, 均低于Hetero处理组。
 | 图1 复合乳酸菌对全株玉米青贮及有氧暴露期pH的影响 3、7、15、30表示青贮发酵的第3、7、15、30天; K1、K3、K7、K15、K30表示开袋有氧暴露第1、3、7、15、30天, 下同。Fig.1 Effect of compound lactic acid bacteria on pH in whole corn silage and aerobic process 3, 7, 15 and 30 mean the 3rd, 7th, 15th and 30th day of silage fermentation; K1, K3, K7, K15 and K30 mean the 1st, 3rd, 7th, 15th and 30th day of aerobic process, the same below. |
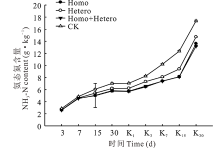
由图2所示, 在整个青贮期和有氧暴露期, 各处理组和对照组的NH3-N含量均逐渐增加, 尤其有氧暴露第15天以后急剧增加, 且对照组始终最高、增幅最大, 与各处理组差异显著(P< 0.05), Homo和Homo+Hetero处理组间无显著差异, 但均显著低于Hetero处理组(P< 0.05)。
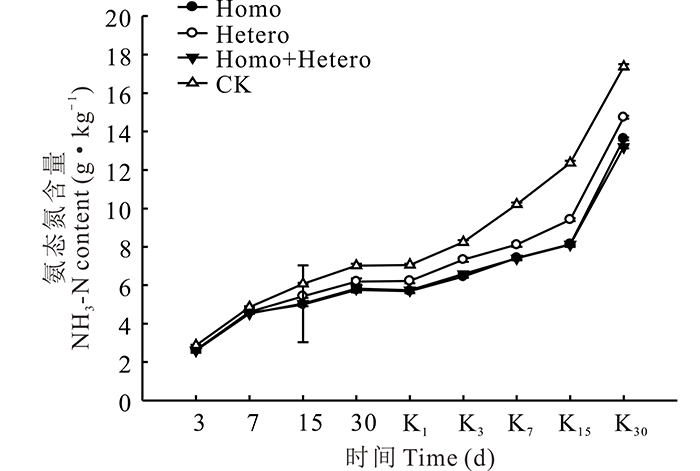 | 图2 复合乳酸菌对全株玉米青贮及有氧暴露期NH3-N含量的影响Fig.2 Effect of compound lactic acid bacteria on NH3-N content in whole corn silage and aerobic process |
如表2可见, 在青贮发酵和有氧暴露后的第30天, LA和AA含量为Homo+Hetero> Homo> Hetero> 对照, Homo+Hetero和Homo处理组间无显著差异, 但与Hetero处理组及对照组差异显著(P< 0.05)。PA和BA在各处理组和对照组中均未检测到。
| 表2 复合乳酸菌对全株玉米青贮期饲料有机酸含量的影响 Table 2 Effect of compound lactic acid bacteria on organic acid content in whole corn silage (mg· mL-1) |
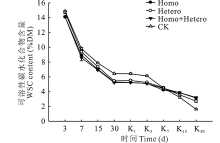
青贮期各处理组和对照组的WSC含量均逐渐降低, 前7 d下降较快, 之后下降速度减缓, 且对照组的WSC含量始终高于各处理组(图3), 青贮第30天时, 对照组与各处理组差异显著(P< 0.05), 但处理组间差异不显著。有氧暴露后, 各处理组和对照组的WSC含量继续保持下降趋势, 对照组于第3天以后快速下降, 并从第15天开始低于各处理组, 第30天时, Homo+Hetero处理组的WSC含量最高, 与Homo处理组无明显差异, 但与Hetero处理组和对照组差异显著(P< 0.05)。
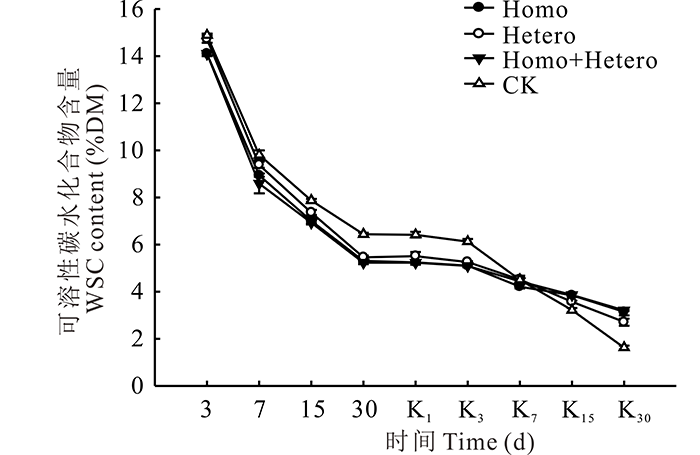 | 图3 复合乳酸菌对全株玉米青贮及有氧暴露期WSC含量的影响Fig.3 Effect of compound lactic acid bacteria on WSC content in whole corn silage and aerobic process |
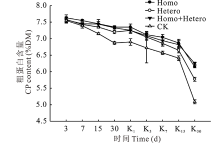
整个青贮期及有氧暴露期, 各处理组和对照组的CP含量均逐渐降低, 且对照组始终低于各处理组(图4), 从青贮第30天开始, 对照组与各处理组差异显著(P< 0.05)。青贮第30天时对照组的CP含量与原料相比损失最多为11.1%, 而Homo和Homo+Hetero处理组损失较少, 分别为4.91%和5.17%。有氧暴露后, 各处理组和对照组的CP含量下降幅度增大, 尤其对照组, 第30天时的CP含量与原料相比损失了36.2%, 与各处理组差异显著(P< 0.05), Homo和Homo+Hetero处理组分别为20.4%和19.2%, CP含量损失较少。
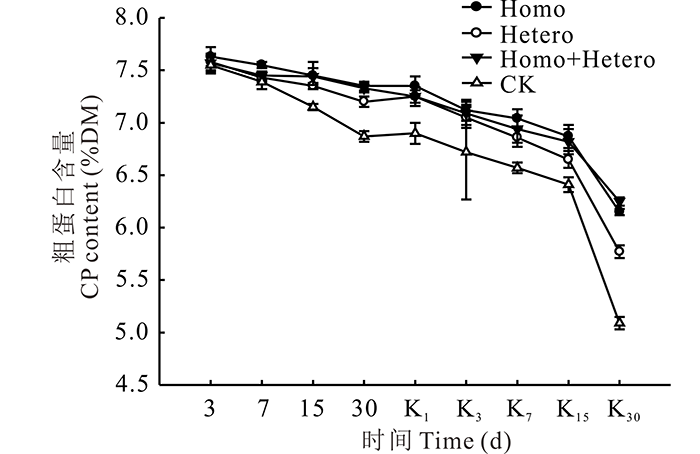 | 图4 复合乳酸菌对全株玉米青贮及有氧暴露期CP含量的影响Fig.4 Effect of compound lactic acid bacteria on CP content in whole corn silage and aerobic process |
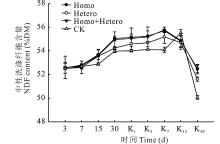
由图5可见, 在青贮过程中, 随着发酵的进行各处理组和对照组的NDF含量均逐渐增加, 有氧暴露后, 继续缓慢增加, 直至第7天达到最高, 之后又大幅下降。有氧暴露15 d后, 青贮饲料中霉菌的数量急剧增加, NDF含量下降可能与霉菌的大量繁殖及其产生的纤维素酶有关。在整个青贮及有氧暴露期间, Homo和Homo+Hetero处理组的NDF含量始终较高, 对照组最低。
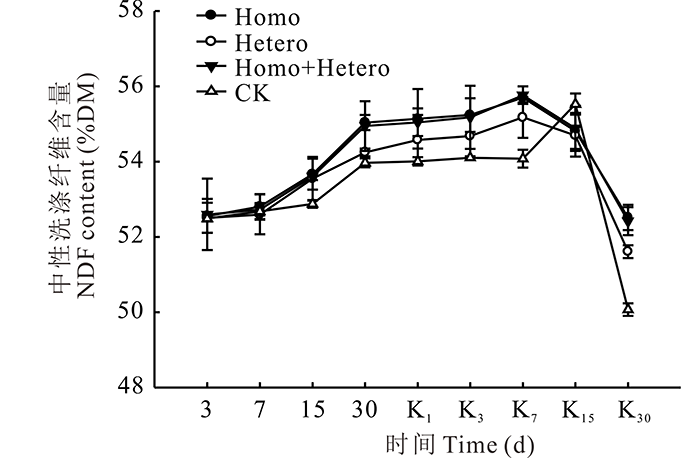 | 图5 复合乳酸菌对全株玉米青贮及有氧暴露期NDF含量的影响Fig.5 Effect of compound lactic acid bacteria on NDF content in whole corn silage and aerobic process |
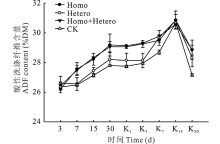
在青贮和有氧暴露期间, 各处理组和对照组的ADF含量变化趋势与NDF含量相似(图6)。
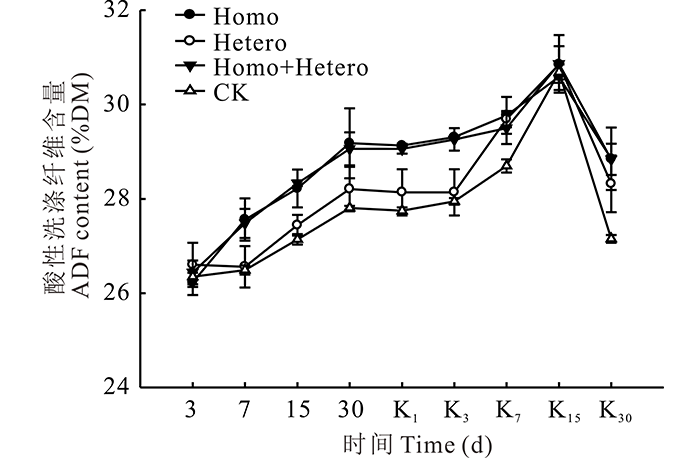 | 图6 复合乳酸菌对全株玉米青贮及有氧暴露期ADF含量的影响Fig.6 Effect of compound lactic acid bacteria on ADF content in whole corn silage and aerobic process |
全株玉米中糖的含量较高, 比较容易青贮, 但开窖后有氧稳定性较差, 容易发生二次发酵, 加入乳酸菌进行青贮可有效提高青贮饲料品质并抑制二次发酵[17]。在青贮过程中, 添加复合乳酸菌的各处理组乳酸菌总数均显著高于对照组, 而好氧细菌、酵母菌和霉菌数量均显著低于对照组, pH亦低于对照组, 表明添加的各复合乳酸菌均能很好的定植和繁殖, 能有效地调控青贮的发酵进程, 显著抑制了有害微生物的繁殖。其中Hetero处理组(肠膜明串珠菌B1-7+屎肠球菌B5-2)在青贮初期乳酸菌总数最多, 这与Stoked等[18]和Kung等[19]的结果一致, 肠系膜明串珠菌和屎肠球菌在青贮发酵早期迅速生长繁殖, 从而为乳酸杆菌的生长创造适宜的条件[18, 19]。与单一乳酸菌添加相比[12], 青贮发酵7 d后乳酸菌数量为Homo+Hetero> Homo> B3-1> Hetero> 对照组, 可见青贮发酵中后期, 单一添加植物乳杆菌B3-1处理组乳酸菌总数高于其他供试乳酸菌, 而3种同型乳酸菌组合Homo处理组高于单一植物乳杆菌B3-1, 5种同型和异型乳酸菌组合Homo+Hetero处理组高于Homo处理组; 相应的, 好氧细菌、酵母菌和霉菌数量大小顺序为Homo+Hetero< Homo< Hetero< 对照组, pH亦然, 青贮第30天的有机酸LA和AA含量也如此, Homo处理组的LA和AA含量比Hetero处理组高, 与一些研究结果不同[11], 但Homo+Hetero处理组LA和AA含量均比Homo处理组更高, 这表明在青贮过程中相同发酵类型的乳酸菌组合添加优于其各组成的单一乳酸菌, 同型和异型乳酸菌组合优于相同发酵类型的乳酸菌组合, 更有利于乳酸菌的增殖发酵, 可能是因为复合乳酸菌能满足青贮发酵不同阶段对乳酸菌的不同需求, 充分发挥协同效应, 促进青贮进程。
同型乳酸菌发酵能提高青贮发酵的品质, 但是在青贮开窖后, 同型乳酸菌发酵不能有效限制酵母菌和霉菌等腐败菌的繁殖, 因此青贮饲料的有氧稳定性降低[20]。本研究发现有氧暴露后, 各处理组和对照组乳酸菌总数随暴露时间延长逐渐减少, 相比来说Homo> Homo+Hetero> Hetero> 对照组; 好氧细菌、酵母菌和霉菌数量逐渐增多, pH逐渐升高, Homo+Hetero< Homo< Hetero< 对照组。表明添加的各复合乳酸菌能不同程度抑制有害微生物的繁殖, 提高全株玉米青贮饲料有氧稳定性, 同型和异型乳酸菌组合要优于同种发酵类型乳酸菌组合。研究结果还表明Homo处理组的好氧细菌、酵母菌和霉菌数量均少于Hetero处理组, 这可能是因为Homo组合中的植物乳杆菌B3-1的添加不仅可增加青贮饲料中的LA含量, 还可提高AA含量[12], 另外植物乳杆菌B3-1对供试的金黄色葡萄球菌、蜡状芽孢杆菌、沙门菌、大肠杆菌和酵母菌都具有较强的抑菌活性[14], 在有氧环境下, 通过抑菌物质抑制有害微生物的活动, 减少了二次发酵, 这与Tabacco等[17]的结果一致, 玉米秸秆青贮饲料的有氧稳定性与酵母菌数量呈负相关; 也可能与供试的异型发酵乳酸菌种类有关, 肠膜明串珠菌和屎肠球菌一般在青贮发酵初期增殖较快, 耐酸性较差, 开窖后相对较低的pH限制了其增殖发酵, 产生的LA和AA较少(表2), 而报道中能提高开窖后有氧稳定性的乳酸菌多是布氏乳杆菌[6, 21]和希氏乳杆菌[22]。
从青贮开始到有氧暴露第7天, 各添加复合乳酸菌的处理组的WSC的含量均低于对照组, 各处理组间无显著差异, 是因为添加的乳酸菌的大量繁殖消耗所致; 而有氧暴露第7天之后, 对照组WSC含量的逐渐低于各处理组, Hetero也逐渐低于Homo和Homo+Hetero处理组, 主要是因为有氧暴露第7天之后, 对照组以及Hetero处理组因产生的LA和AA较少, 不能有效抑制好氧微生物的繁殖, 引起WSC含量的损耗高于乳酸菌繁殖的消耗。在青贮发酵及有氧暴露期, 各添加复合乳酸菌的处理组的CP、NDF和ADF含量变化呈相似的变化趋势, 在同一取样时间点, 均是Homo+Hetero> Homo> Hetero> 对照组, Homo+Hetero与Homo处理组无明显差异, NH3-N含量则相反。以上结果表明, 添加复合乳酸菌可显著改善全株玉米青贮饲料营养品质, 抑制二次发酵, 延缓青贮饲料在有氧暴露阶段的腐败, 其中Homo+Hetero处理组效果最优。
添加复合乳酸菌能有效地增加青贮过程和有氧暴露后饲料中乳酸菌的数量、LA和AA的含量, 减少好氧细菌、酵母菌和霉菌数量, 降低pH和NH3-N含量, 减少CP含量的损失, 显著改善青贮饲料发酵品质和营养品质, 抑制青贮饲料有氧暴露后的二次发酵, 其中同型和异型乳酸菌组合成的Homo+Hetero处理效果最优。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|