



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
不同株型玉米基因型对干旱胁迫的响应分析
[赵小强1, *  , 陆晏天
, 陆晏天1 , 白明兴1 , 徐明霞1 , 彭云玲1, * , 丁永福1 , 庄泽龙1 , 陈奋奇1 , 张大志2 ]
, 陆晏天, 丁永福|
|


作者简介:赵小强(1990-),男,甘肃陇西人,副教授,博士。E-mail: zhaoxq3324@163.com
改良株型结构是挖掘玉米产区光、热、水潜力的重要途径,是玉米理想株型育种的基础。为了深入揭示玉米株型相关耐旱响应机理,研究在花期干旱胁迫下对株型差异较大的3份玉米基因型(廊黄、昌7-2和TS141)穗位叶的株型结构、气孔形态、生理代谢、光合特性及产量进行了研究。结果表明:1)干旱胁迫下,3份不同株型基因型的叶长、叶宽、叶面积及叶夹角均显著减少,叶形系数和叶向值均显著增大,平展株型基因型TS141的6个株型性状较紧凑株型基因型廊黄和昌7-2变化更大;2)干旱导致3份不同株型基因型的上/下表皮气孔个数趋于增多,上/下表皮气孔长度和宽度趋于缩短,TS141的上/下表皮气孔个数均有较大的增多,上/下表皮气孔长度和宽度均有较小的降低;3)干旱胁迫后,TS141的叶绿素a、b、(a+b)含量及叶绿素a/b均明显下降,廊黄和昌7-2的叶绿素a/b均有较小地上升,其余3个性状均有较小地下降;4)干旱条件下,3份不同株型基因型的气孔导度、净光合速率、叶片温度、蒸腾速率和RuBPCase活性均有不同程度地减少,胞间CO2浓度均有不同程度地升高,与TS141相比,廊黄和昌7-2变化的幅度更小;5)干旱处理后,3份不同株型基因型的单株产量、单穗重、穗粒重、百粒重、穗长及出籽率均显著下降,对TS141的影响程度远大于廊黄和昌7-2;6)主成分PCA分析和热图Hierarchical clustering分析表明,干旱胁迫下玉米这些性状间的关系较为复杂,彼此相互关联,这些性状能完全概括不同株型基因型的耐旱性差异。因此,推测玉米株型结构作为重要的干旱调节者,其可与气孔形态、生理代谢及光合特性相互调节共同构成玉米株型相关耐旱响应机制,且由于紧凑株型玉米对干旱胁迫的调节能力更强,其在今后育种中具有更好的抗旱潜能。
, LU Yan-tian, DING Yong-fuImprovement of plant architecture is an important way to fully exploit the potential of environmental resources of light, heat, and water in maize ( Zea mays) production, and it is the basis of breeding targeting ideal plant type. This research studied the response mechanisms of drought tolerance related to plant architecture in maize. The plant architecture traits, stomatal morphologies, physiological metabolisms and photosynthesis characteristics of ear leaf, and yield were recorded for three different maize genotypes (Langhuang, Chang7-2, and TS141) with differing plant architecture under drought stress during the flowering stage. It was found that: 1) Leaf length, leaf width, leaf size, and leaf angle were decreased significantly, and leaf shape value and leaf orientation value of the three maize genotypes were increased significantly under drought stress. Moreover, genotype TS141, characterized as ‘flat’, had larger changes of these six plant architecture traits than genotypes characterized as ‘compact’ (Langhuang and Chang 7-2); 2) Drought stress increased the numbers of upper and lower epidermal stomata of all three maize genotypes, but decreased stomatal length and width. The change in stomatal number of TS141 was greater than for the other cultivars, and the size reduction was smaller; 3) Chlorophyll a, b, and (a+b) contents, and chlorophyll a:b ratio of TS141 were significantly decreased by drought. For the tested genotypes, chlorophyll a:b ratio had a smaller increase, and the other three traits had a smaller decrease in Langhuang and Chang 7-2 after drought stress than in TS141; 4) Stomatal conductance, net photosynthetic rate, the temperature of leaf thermocouples, transpiration rate, and RuBPCase activity decreased to different degrees in different genotypes, and intercellular CO2 concentration increased to different degrees in the three tested maize genotypes under drought stress. However, compared with TS141, Langhuang and Chang 7-2 had smaller changes; 5) Yield per plant, ear weight, grain weight, 1000-seed weight, ear length, and kernel ratio of all three maize genotypes were decreased significantly under drought stress, and TS141 was affected more severely than Langhuang and Chang 7-2; 6) Principal component analysis and a heat map with hierarchical cluster analysis indicated that the relationships among these traits in maize under drought stress were complex and interrelated with each other, and could completely account for the differences in drought tolerance between the different plant architecture genotypes. Therefore, it is deduced from these results that plant architecture may be an important drought regulator that can be co-regulated with stomatal morphologies, physiological metabolisms, and photosynthetic characteristics to form the response mechanisms of drought tolerance. Moreover, because 'compact' maize genotypes displayed stronger drought tolerance, these types have better drought tolerance potential for use in future breeding work.
玉米(Zea mays)作为重要的粮食、饲料及生物质能源, 其在保障国家粮食安全、促进畜牧业发展及提高人民生活水平等方面发挥着十分重要的作用。然而玉米对干旱比较敏感, 干旱缺水不仅抑制玉米正常生长发育、干扰各种生理代谢, 进而严重影响产量和品质[1, 2, 3, 4], 据估计1980-2015年干旱导致全球玉米产量减少了39.3%[5]。因此, 提高玉米的耐旱性并进行耐旱育种已成为解决玉米高产稳产的重要途径。
Jensen[6]在1949年最早描述了株型对植物群体光合性能的影响, 即植物各叶片光合作用效能相等、叶面积总量相同的状况下, 光合作用总量主要取决于各叶片的空间排列。而后, Donald[7]在1968年提出了植物理想株型(ideal-type)的概念, 指出理想株型是集光合作用、生长发育及产量最优的植物构造模式。Duvick[8]在2005年研究发现, 通过育种及遗传改良玉米株型结构, 显著改善了玉米群体的密植性、冠层光合特性、通风透气性及抗倒伏特性, 使美国玉米产量在过去80年里增加了8倍。另外, 近些年来学者已在玉米、小麦(Triticum aestivum)、水稻(Oryza sativa)、大豆(Glycine max)、油菜(Brassica napus)等多种作物中研究指出, 植物的株型结构可以感知水分变化等环境信号, 并能通过调节株型结构的变化有效地缓解植物对水分胁迫的伤害, 最终改善植物的耐旱性[2, 3, 4, 9, 10, 11, 12, 13, 14]。何海军等[15]研究表明, 干旱胁迫时, 平展株型玉米品种豫玉22和中间株型品种沈单16的产量和水分利用效率均显著低于Pioneer公司选育的紧凑株型品种先玉335, 而其耐旱系数在3个品种间则相反依次升高。彭云玲等[16]报道, 玉米具有一定的株型调节能力, 缺水时其通过调节叶夹角的收缩、叶向值的增大等改变株型结构, 以减少植株对太阳辐射的接收和水分的散失, 进而增强玉米植株的耐旱性。Zhao等[3]在8种水旱环境下剖析了2套F2∶ 3群体叶长、叶宽、叶夹角、叶面积、叶形系数及叶向值等株型结构的分子遗传机理, 在干旱环境下检测到了29个稳定表达的sQTLs (stable QTLs), 并通过meta-QTL分析, 在21个meta-QTLs区间内预测到了lg1 (liguleless 1)、Kn1-DL (knotted 1)等调控玉米株型发育的24个候选基因。因此, 遗传改良、选育出株型理想或合理的玉米新品种, 使其在改善株型结构的同时, 又能显著提高品种的耐旱性, 以实现品种高产已成为最重要的玉米育种策略之一。然而, 关于玉米株型结构与耐旱性的内在关系及株型相关耐旱机理的研究目前尚不清楚。因此, 基于以上考虑, 本研究以株型差异较大的3份玉米基因型为试验材料, 花期在干旱胁迫下, 通过系统地研究不同株型基因型玉米穗位叶的株型结构、气孔形态、生理代谢、光合特性及产量变化, 以期揭示玉米株型相关耐旱响应机理, 为玉米株型相关耐旱分子遗传机理剖析、株型相关耐旱种质遗传改良及抗旱高产育种提供理论参考。
试验采用遮雨棚于2018年在甘肃农业大学校内试验田(36.08° N, 103.70° E, 海拔1525 m)进行。试验地为沙壤土, 中等肥力, 前茬种植玉米, 播前施尿素80 kg· hm-2。采用裂区试验设计, 试验地划分为旱区和水区等2个一级小区, 再将每个一级小区又细分为3个二级小区随机排列廊黄、昌7-2和TS141, 每个二级小区3.0 m2(1.5 m× 2.0 m), 并于4月26日播种, 行距50 cm, 株距20 cm, 每行10株, 9月30日收获。旱区干旱胁迫处理参照彭云玲等[1]的方法, 即在玉米开花前10 d开始停止灌水, 18 d后恢复灌水; 水区正常供水处理每隔7~10 d灌水一次, 直到玉米生理成熟。待玉米干旱胁迫处理14 d后取3株长势整齐一致的植株穗位叶, 测定其穗位叶的株型结构、上/下表皮气孔形态、叶绿素含量、光合特性及产量。
1.3.1 株型结构和产量 参照Zhao等[3]和赵小强等[18]的方法, 测定每一处理下3份基因型植株穗位叶的叶长(leaf length, LL)、叶宽(leaf width, LW)、叶夹角(leaf angle, LA)、叶面积(leaf size, LS; LS=LL× LW× 0.75)、叶形系数(leaf shape value, LSV; LSV=LL/LW)和叶向值[leaf orientation value, LOV; LOV=(90-LA)× (LF/LL)]。式中LF(length from beginning of ligule to flagging point of leaf)表示高点长。待玉米生理成熟后收获并自然风干, 参照石云素等[19]制定的《玉米种质资源描述规范与数据标准》和赵小强等[20]的方法测定其单株产量(yield per plant, YPP)、单穗重(ear weight, EW)、穗粒重(grain weight, GW)、百粒重(100-kernel weight, KW)、穗长(ear length, EL)和出籽率(kernel ratio, KR; KR=GW/EW× 100%)。
1.3.2 上/下表皮气孔 参照Yu等[21]的叶片表面压印法, 制作每一处理下3份基因型植株穗位叶距离叶脉及叶片边缘中间相同、相对部位的上/下表皮切片(叶片大小为1.0 cm× 1.0 cm), 再将切片置于10× 10倍的Leica CTR6000倒置荧光显微镜观察玉米穗位叶上/下表皮气孔个数(No. of upper/lower epidermal stomatal), 利用Motic Images Advanced 3.2 软件测量上/下表皮气孔长度(upper/lower epidermal stomatal length)和上/下表皮气孔宽度(upper/lower epidermal stomatal width)。
1.3.3 叶绿素 参照Lichtenthaler等[22]的方法, 即用5 mL 95%的乙醇避光浸泡每一处理下3份基因型植株穗位叶中部0.1 g至叶片发白为止, 然后稀释10倍, 在665和649 nm下测定吸光值, 再按照公式计算叶绿素a、b(chlorophyll a and b, Chl a and Chl b)的浓度(Chl a=13.95× A665-6.88× A649, Chl b=24.96× A649-7.32× A655)和含量(Chl a and b content, Chl a/Chl b含量=Chl a/Chl b浓度× 提取液体积× 稀释倍数/样品鲜重/1000), 并计算总叶绿素含量[chlorophyll (a+b) content, Chl (a+b)]。类囊体的垛叠程度用叶绿素a/b(chlorophyll a/b, Chl a/b)进行估测[23]。
1.3.4 光合特性 采用美国LI-6400/XT便携式光合仪测定每一处理下3份基因型植株穗位叶中部的气孔导度(stomatal conductance, Gs)、净光合速率(net photosynthetic rate, Pn)、胞间CO2浓度(intercellular CO2 concentration, Ci)、叶片温度(temperature of leaf thermocouple, Tl)和蒸腾速率(transpiration rate, Tr)。具体测定时间选在晴天上午的9:00-11:00进行, 光合有效辐射(photosynthetic active radiation, PAR)设为1000 μ mol· m-2· s-1, CO2浓度设为环境浓度。参照徐芬芬等[23]的方法估测叶肉细胞核酮糖1, 5-二磷酸羧化酶活性(RuBPCase activity, RuBPCase; RuBPCase=Pn/Ci× 100%)。
所有试验数据均采用IBM SPSS 19.0 (SPSS Inc., Chicago, IL, USA)软件统计分析, 试验结果采用mean± SD (standard deviation)表示, 不同干旱处理下上述所测性状采用Duncan法进行方差分析。再将不同干旱处理下所测性状转化成各自的耐旱系数, 采用IBM SPSS 19.0软件进行PCA(principal component analysis)主成分分析, 采用MeV 4.9.0软件进行热图(Hierarchical clustering)分析, 距离度量标准采用Pearson相关系数距离。
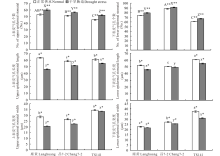
正常供水下, 花期廊黄、昌7-2及TS141等3份不同株型基因型穗位叶的6个株型性状间差异显著(P< 0.01或P< 0.05); 除叶向值外, 平展株型基因型TS141的叶长、叶宽、叶面积、叶形系数、叶夹角均显著高于紧凑株型基因型廊黄和昌7-2(表1)。表明, 这6个株型性状受玉米本身遗传调控较大, 不同株型基因型在株型表现上差异明显。花期干旱胁迫下, 3份不同株型基因型穗位叶的叶长、叶宽、叶面积及叶夹角均显著降低(P< 0.01或P< 0.05), 而叶形系数和叶向值则显著升高(P< 0.01或P< 0.05); 干旱胁迫下这6个株型性状在不同基因型间的变化程度存在显著差异(P< 0.01或P< 0.05)(表1)。与TS141相比, 干旱胁迫后廊黄和昌7-2的这6个株型性状的变化幅度较低, 其分别平均降低了20.27%和8.49%、25.90%和14.56%、31.87%和19.75%、-7.73%和-12.18%、26.08%和17.33%、-39.12%和-9.06%。说明, 干旱胁迫能显著改变玉米叶片的株型构造, 以适应胁迫对不同株型玉米的影响, 干旱对平展株型基因型叶片株型的影响远大于紧凑株型基因型。因此, 紧凑株型玉米的耐旱性明显强于平展株型玉米。
| 表1 干旱胁迫对3份不同株型玉米基因型穗位叶6个株型性状的影响 Table 1 Effects of drought stress on six plant architecture traits of ear leaf in three different plant architecture maize gene types |
花期正常浇水下, 3份不同株型基因型穗位叶的上/下表皮气孔个数、长度及宽度间差异显著(P< 0.01或P< 0.05), 紧凑株型基因型廊黄、昌7-2穗位叶的上/下表皮气孔个数显著多于(平均多8/20个)平展株型基因型TS141, 而上/下表皮气孔长度和宽度均显著短于(平均短0.17/9.26 μ m和6.92/12.39 μ m)TS141(图1和2)。暗示玉米的上/下表皮气孔个数、长度及宽度在不同株型基因型间不同, 与紧凑株型玉米相比, 相同单位面积平展株型玉米的上下表皮气孔个数较少, 但由于平展株型玉米叶片较大, 就其相对叶片而言, 平展株型玉米的上下表皮气孔个数较多, 且相同单位面积平展株型玉米的上/下表皮气孔长度和宽度均较长, 意味着平展株型玉米叶片与外界环境进行气体和水分交换的能力大于紧凑株型玉米。花期干旱胁迫下, 3份不同株型基因型穗位叶的上/下表皮气孔个数显著升高外(P< 0.01), 其上/下表皮气孔长度和宽度均显著降低(P< 0.05), 且干旱胁迫下上/下表皮气孔个数、长度及宽度在不同株型玉米间的变化程度不同(P< 0.01或P< 0.05)(图1和2)。说明, 水分亏缺会显著影响不同株型玉米的上下表皮气孔的排列和大小, 以影响叶片的蒸腾、呼吸及光合性能。另外, 缺水胁迫后, TS141和廊黄/昌7-2的上/下表皮气孔个数、长度及宽度分别平均降低了-15.56%和-9.43%、9.63%和19.24%、3.21%和21.85%、-9.84%和-5.08%、9.47%和7.43%、16.96%和5.65%。证实与平展株型玉米相比, 紧凑株型玉米上/下表皮气孔的长度和宽度降低更大, 上/下表皮气孔个数升高更小。因此, 紧凑株型玉米受干旱的影响均较平展株型玉米更小。
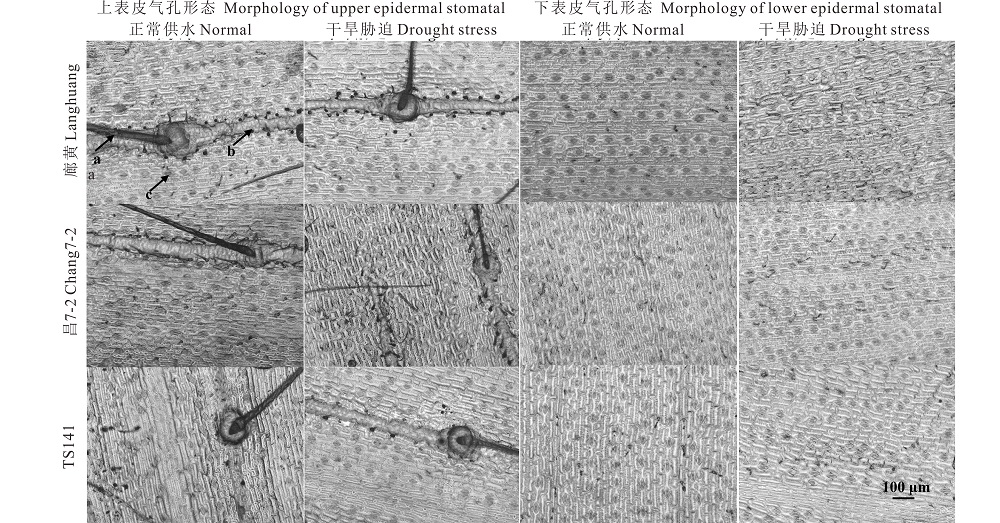 | 图1 干旱胁迫下3份不同株型玉米基因型穗位叶上/下表皮气孔的显微结构 a、b、c分别为绒毛、叶脉和气孔。Fig.1 Microstructure of upper/lower epidermal stomatal of ear leaf in three different plant architecture maize genotypes under drought stress a, b, and c indicate the tomentum, vein, stomatal, respectively. |
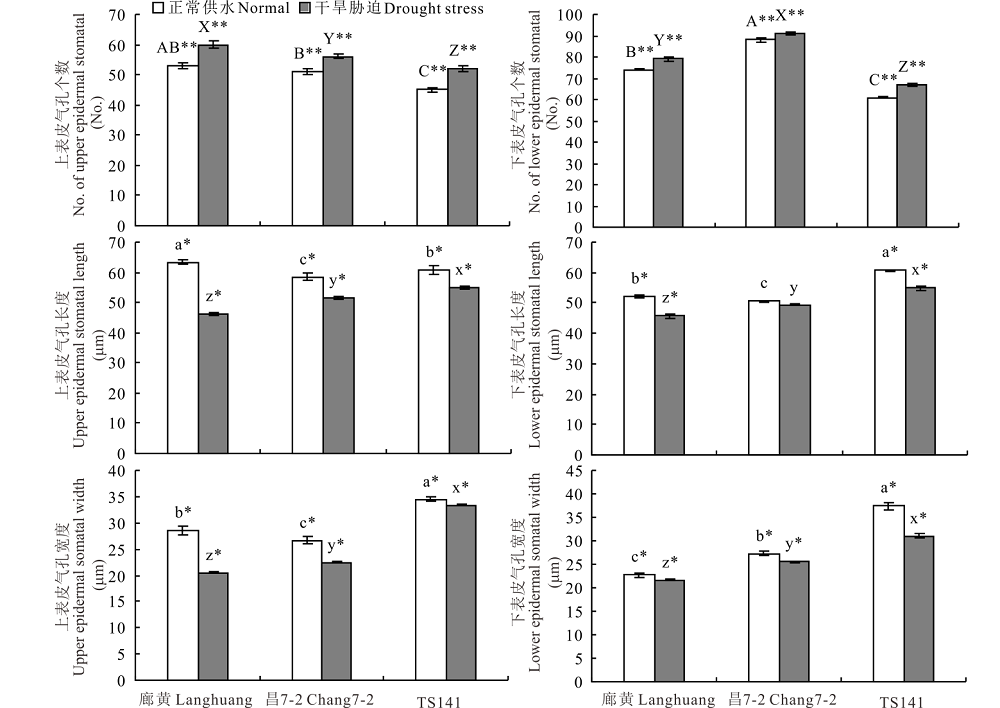 | 图2 干旱胁迫对3份不同株型玉米基因型穗位叶上/下表皮气孔的影响 同一玉米基因型单个性状在不同水分处理标有“ * * 或* ” 分别表示P=0.01或P=0.05水平下差异显著; 不同玉米基因型单个性状在正常供水/干旱胁迫处理下标有“ A、B、C” 或“ a、b、c” /“ X、Y、Z” 或“ x、y、z” 分别表示P=0.01或P=0.05水平下差异显著。下同。Fig.2 Effects of drought stress on upper/lower epidermal stomatal of ear leaf in three different plant architecture maize gene types The “ * or * * ” with single traits of one maize genotype under different watering conditions indicate significant difference at P=0.01 or P=0.05 level, respectively. The “ A, B , C” or “ a, b, c” /“ X, Y, Z” or “ x, y, z” with single traits of different maize genotypes under well-watered/water-stressed conditions indicated significant difference at P=0.01 or P=0.05 level, respectively. The same below. |
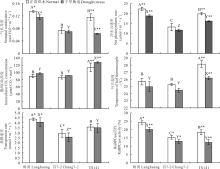
正常供水处理, 3份不同株型基因型花期穗位叶的叶绿素a、b、a+b含量及叶绿素a/b间差异显著(P< 0.01或P< 0.05), 这4个性状均表现为廊黄> TS141> 昌7-2(图3)。说明, 不同株型基因型的叶绿素含量间差异明显, 其可能是不同株型玉米叶绿素合成酶活性及叶绿素合成速率不同引起的。花期缺水处理, 紧凑株型廊黄、昌7-2穗位叶的叶绿素a、b、(a+b)含量均降低(P> 0.05), 而平展株型TS141的这3个性状均显著降低(P< 0.01或P< 0.05)(图3)。此外, 花期廊黄和昌7-2在缺水处理下穗位叶的叶绿素a/b均升高(P> 0.05), 而TS141的叶绿素a/b显著降低(P< 0.05)(图3)。说明, 缺水能抑制不同株型玉米叶绿素合成酶活性及叶绿素合成速率, 或能加速叶绿素降解酶活性及叶绿素降解速率。与平展株型玉米相比, 干旱胁迫对紧凑株型玉米的这4个性状的影响程度更小。
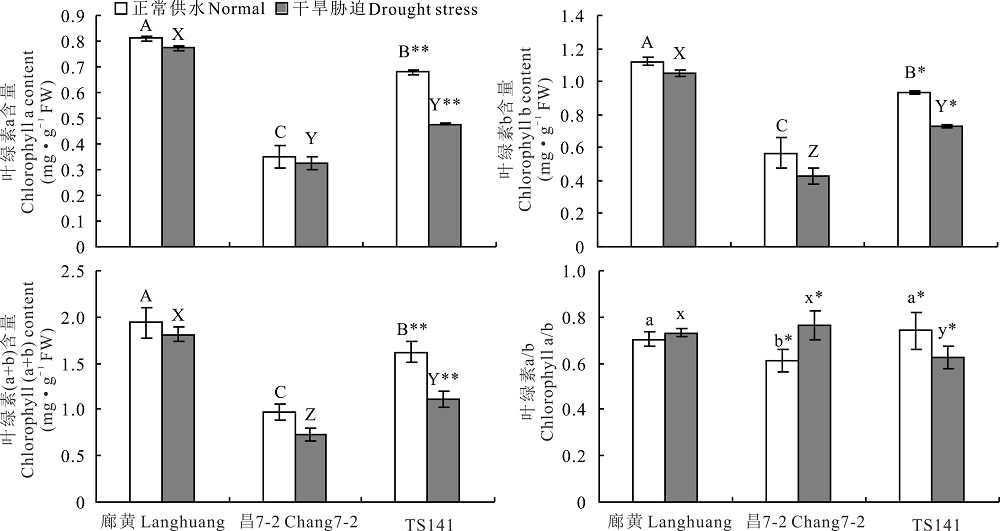 | 图3 干旱胁迫对3份不同株型玉米基因型穗位叶4个叶绿素相关性状的影响Fig.3 Effects of drought stress on four chlorophyll related traits of ear leaf in three different plant architecture maize gene types |
花期3份不同株型基因型正常供水下, 其穗位叶的气孔导度、净光合速率、胞间CO2浓度、叶片温度、蒸腾速率及RuBPCase活性在不同基因型间差异显著(P< 0.01), 且平展株型基因型(TS141)穗位叶的这6个性状分别是紧凑株型基因型(廊黄和昌7-2)的1.12、1.12、1.29、1.11、0.98和0.94倍(图4)。揭示由于受基因型本身遗传特性的调控, 导致不同株型玉米的气孔开度、光合性能、植株吸水与散热状况及类囊体的垛叠程度也不尽相同。此阶段干旱胁迫后, 3份不同株型基因型穗位叶的胞间CO2浓度则显著升高(P< 0.01), 其余5个性状均明显降低(P< 0.01或P< 0.05), 且紧凑株型廊黄和昌7-2的这6个性状在胁迫下的变化幅度远小于平展株型TS141, 其分别平均降低了10.14%和45.69%、14.64%和18.49%、-6.79%和-12.64%、7.07%和3.00%、1.99%和9.36%、32.74%和16.32%(图4)。说明, 干旱胁迫能显著抑制不同株型玉米的气孔开度、光合性能、植株吸水与散热状况及类囊体的垛叠程度, 紧凑株型玉米的这种抑制程度明显低于平展株型玉米, 耐旱能力更强。
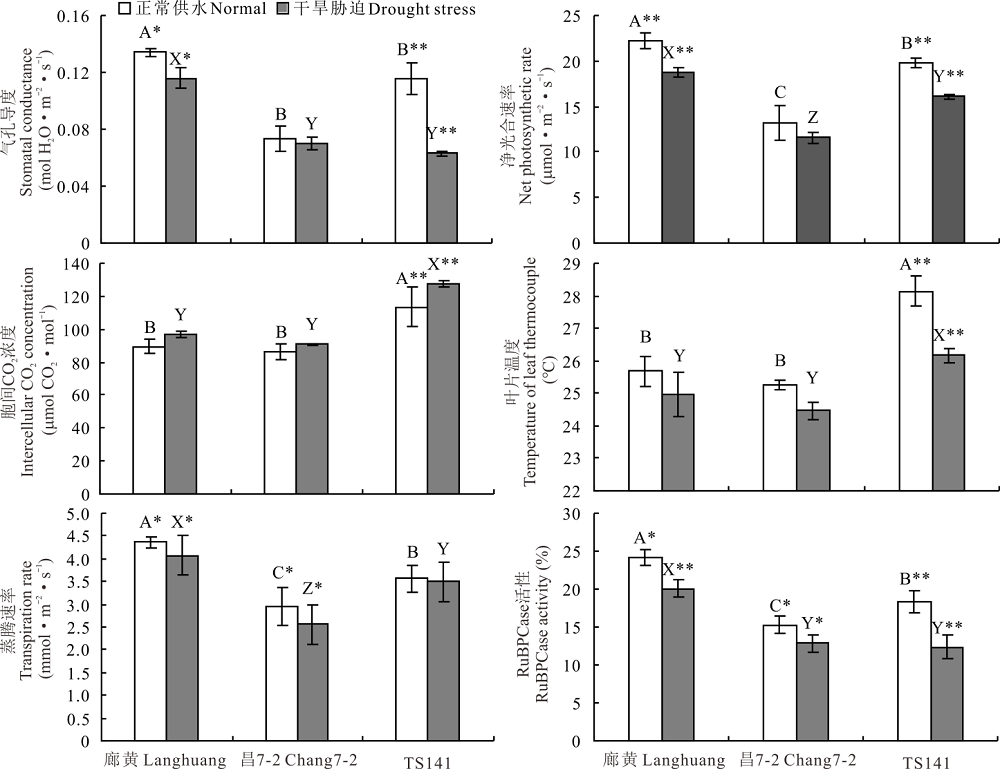 | 图4 干旱胁迫对3份不同株型玉米基因型穗位叶6个光合特性的影响Fig.4 Effects of drought stress on six photosynthetic characteristics of ear leaf in three different plant architecture maize gene type |
花期正常供水下, 由于3份不同株型基因型所携带有丰富的遗传背景, 因此, 其在单株产量、单穗重、穗粒重、百粒重、穗长及出籽率间的差异显著(P< 0.01), 且平展株型TS141除了出籽率显著低于紧凑株型廊黄和昌7-2外, 其余产量性状均显著高于紧凑株型玉米(表2)。3份不同株型基因型在花期干旱胁迫后6个产量性状均显著降低(P< 0.01或P< 0.05), 且干旱胁迫下廊黄和昌7-2的6个产量性状的降低幅度远小于TS141, 其分别平均降低了13.77%和22.68%、12.48%和21.19%、13.95%和15.25%、24.04%和38.98%、11.34%和24.43%、3.71%和7.71%(表2)。表明, 花期缺水能显著影响玉米产量, 其单株产量、单穗重、穗粒重、百粒重、穗长及出籽率均显著降低。干旱胁迫下紧凑株型玉米比平展株型玉米的产量降低更少, 耐旱性更强。
| 表2 干旱处理对3份不同株型玉米基因型6个产量性状的影响 Table 2 Effects of different watering conditions on six yield traits in three different plant architecture maize gene types |
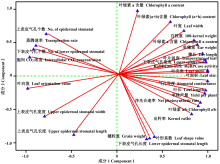
花期3份不同株型基因型在2种水旱处理下对其穗位叶的6个株型性状、6个上/下表皮气孔性状、4个生理性状、6个光合特性及6个产量性状进行分析处理, 计算各性状的耐旱系数。对这些性状的耐旱系数进行PCA主成分分析表明, 从主成分分析的初始化特征值和因子提取后的特征值可知, 28个主成分中前2个主成分的特征值均大于1, 且累计贡献率达到了100%(表3)。说明, 前两个主成分能代表28个性状的全部信息。因此, 可以用这28个性状对3份不同株型玉米的耐旱性进行概括。其中决定第一主成分的主要是叶长、叶面积、叶夹角、下表皮气孔宽度、叶绿素a含量、叶绿素a/b、气孔导度、净光合速率、叶片温度、RuBPCase活性、单株产量、单穗重、百粒重、穗长、出籽率, 可以概括为株型因子、气孔因子、生理代谢因子、光合因子及产量因子, 解释70.53%的贡献率; 决定第二主成分的主要包括叶绿素b及叶绿素(a+b)含量, 可以概括为生理代谢因子, 解释29.47%的贡献率(图5和图6)。
| 表3 所测28个相关性状的耐旱系数主成分分析 Table 3 Principal analysis of drought resistance coefficient with 28 measured traits |
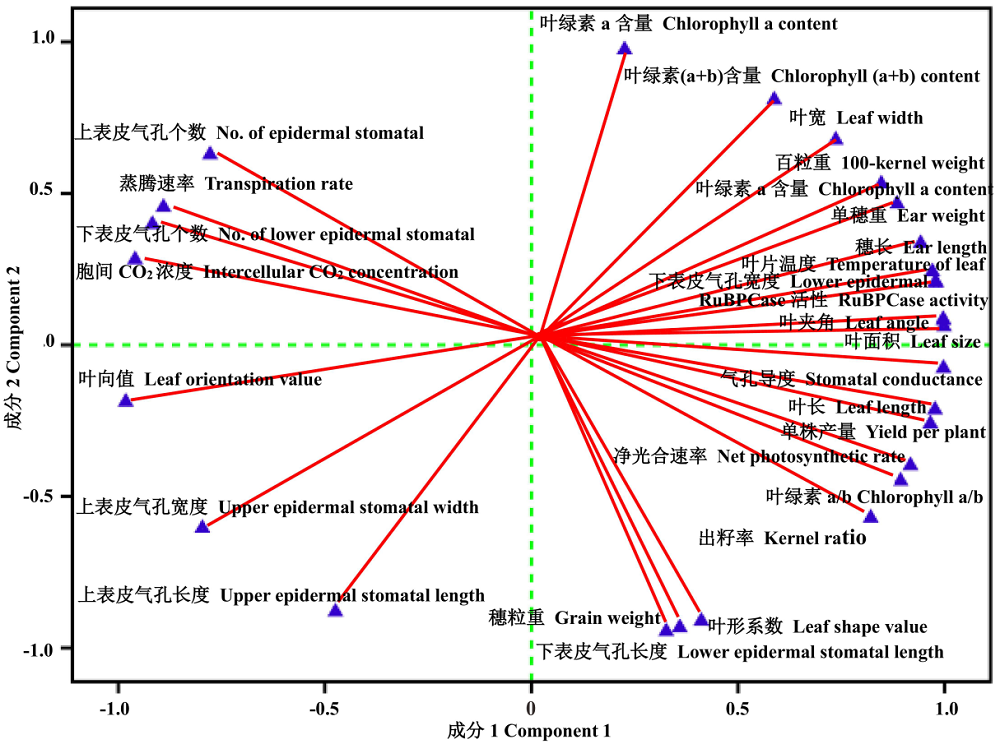 | 图5 所测28个相关性状的耐旱系数主成分载荷图Fig.5 Biplot of drought resistance coefficient with 28 measured traits |
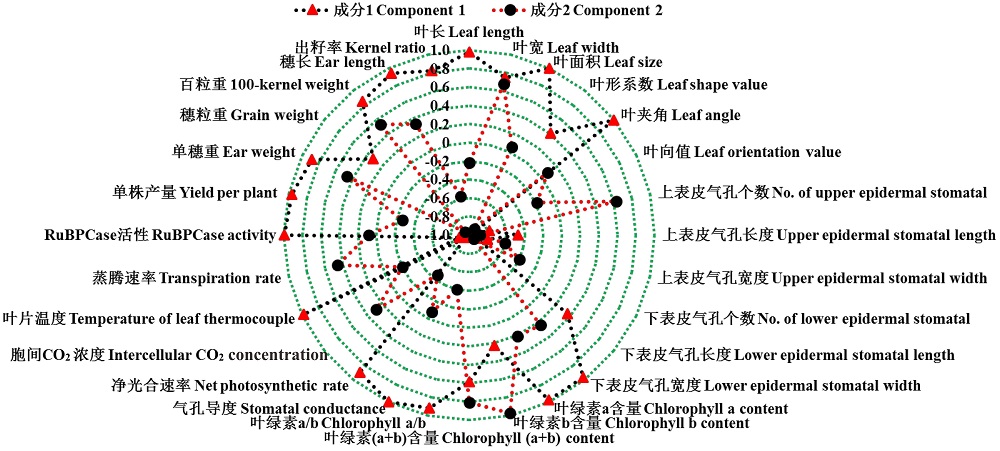 | 图6 所测28个相关性状的耐旱系数主成分特征向量Fig.6 Eigenvectors of drought resistance coefficient with 28 measured traits |
根据各性状的耐旱系数进行热图Hierarchical clustering分析表明, 利用这28个性状的耐旱系数在Pearson相关系数距离为0.63处可以将3份不同株型基因型划分为耐旱玉米廊黄和昌7-2及旱敏感玉米TS141(图7)。此外, 这28个性状的耐旱系数在Pearson相关系数距离0.26处被划分成了3类, 即第一类包括7个性状为叶向值、上表皮气孔长度、宽度、个数、胞间CO2浓度、下表皮气孔个数及蒸腾速率; 第二类涵盖了9个性状为叶绿素a、b、(a+b)含量、叶宽、叶片温度、下表皮气孔宽度、单穗重、百粒重及穗长; 第三类主要为叶形系数、下表皮气孔长度、气孔导度、叶面积、叶夹角、叶长、RuBPCase活性、叶绿素a/b、净光合速率、穗粒重、单株产量及出籽率等12个性状(图7)。表明玉米的这6个株型性状、6个上/下表皮气孔性状、4个生理性状、6个光合特性及6个产量性状彼此之间紧密关联, 相互作用共同决定玉米耐旱能力的强弱。进一步分析还发现, 叶长、叶面积、叶夹角、下表皮气孔宽度、叶绿素a、b及(a+b)含量、叶绿素a/b、气孔导度、净光合速率、叶片温度、RuBPCase活性、单株产量、单穗重、百粒重、穗长、出籽率的颜色区分较明显, 能够较好地区分3份不同株型玉米基因型的耐旱性差异(图7), 这与 PCA主成分分析的结果一致, 说明这些性状可用于区分不同株型玉米的耐旱能力。
 | 图7 3份不同株型玉米基因型所测28个相关性状的耐旱系数热图Hierarchical clustering分析Fig.7 Heat map with hierarchical clustering analysis of drought resistance coefficient with 28 measured traits in three different plant architecture maize gene types |
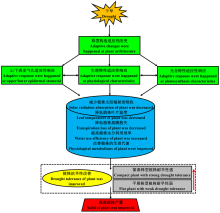
本研究进一步推测了玉米株型相关耐旱响应机制(图8), 即干旱等环境信号作用下, 不同株型玉米的株型结构均会对干旱胁迫产生适应性的改变, 株型结构的变化会进一步调节气孔形态、生理代谢及光合特性, 或不同株型玉米的气孔形态、生理代谢及光合特性间也会相互影响, 以减少不同株型玉米植株对太阳辐射的接收、降低植株的叶片温度及蒸腾散失、提高植株的水分利用效率、改善植株的生理代谢, 最终达到改善玉米耐旱性的目的, 并提高玉米的产量。此外, 本研究也证实了紧凑株型玉米廊黄和昌7-2较平展株型玉米TS141更耐旱, 可能与紧凑株型玉米具有更合理的株型结构有关。
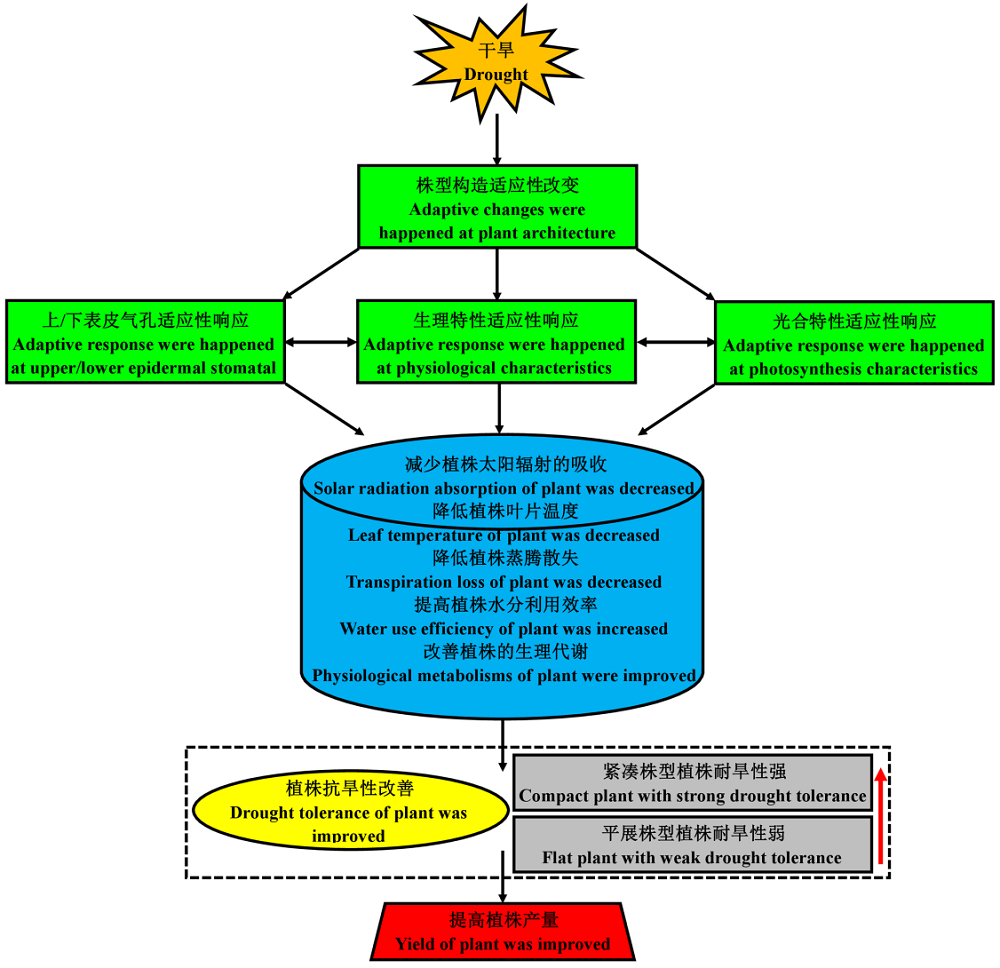 | 图8 不同株型玉米基因型可能的耐旱响应机制Fig.8 Possible response mechanism of drought tolerance in different plant architecture maize gene types |
叶片作为玉米进行光合、蒸腾及抗逆的主要器官, 是玉米生命活动的基础, 其叶长、叶宽、叶形系数、叶面积、叶夹角及叶向值等协同作用共同塑造了玉米株型结构, 最终影响玉米的产量及品质。目前学者[4, 18, 24, 25, 26, 27]对玉米株型结构的研究主要集中在株型结构与密植性、耐遮荫性及抗倒伏性的关系方面, 认为理想株型玉米在实现密植的同时, 还可以实现个体间竞争最小化及光合作用最大化, 以改善玉米生长状况, 最终显著提高玉米产量。而关于株型结构与耐旱性的关系研究较少。Zhao等[2, 3, 4]和赵小强[13]利用3份株型及耐旱性差异较大的玉米自交系及其组配的2个F1杂交种、2套F2及2套F2∶ 3群体在9种水旱环境下剖析了玉米株型结构与耐旱性间的关系, 分析表明大喇叭口期及花期干旱胁迫时, 除叶向值显著升高外, 株高、穗位高、穗位比、叶长、叶宽、叶面积、叶夹角及雄穗分枝数等株型性状均显著降低, 这些株型性状与玉米耐旱性状ASI(anthesis-silking interval)及产量紧密相关, 紧凑株型玉米比平展株型玉米具有更强的耐旱性。Ribaut等[28]和Jeff[29]报告发现玉米耐旱株型应具有相对矮小的植株、较粗的茎秆、较小的雄穗和穗上叶片、上举的叶片、较好的持绿性及发达的根系。在2种水旱环境下董红芬等[30]以20份玉米杂交种为试验材料研究发现玉米的耐旱性状与株型性状间的关联度较为复杂, 其中ASI与雄穗分枝数、叶片卷曲度及穗秃尖与叶片数的关联度最大。本研究结果表明, 3份不同株型玉米花期在干旱胁迫时, 其穗位叶的叶长、叶宽、叶面积及叶夹角均显著降低, 叶形系数和叶向值则显著升高, 这与干旱胁迫下彭云玲等[16]、Zhao等[3]对玉米的研究、Yang等[14]对小麦的研究及王晋[31]对水稻的研究结果相一致。本研究进一步分析发现, 花期干旱胁迫下紧凑株型廊黄和昌7-2的单株产量、单穗重、穗粒重、百粒重、穗长及出籽率比平展株型TS141降低更少, 这与赵小强等[32]、董红芬等[30]和何海军等[15]的研究结果相一致。说明玉米的株型结构可以感知外界水分信号的变化, 并能通过调节其株型结构来适应玉米植株对干旱胁迫的损害, 以提高玉米的耐旱性。因此, 由于存在株型效应的影响, 紧凑株型玉米可以通过自身株型优势, 较平展株型玉米具有更强的耐旱能力。
干旱环境压力下气孔调节是植物适应干旱环境、抵御干旱的重要机制之一, 而环境水分条件也会影响气孔的发育及开度[33]。本研究发现3份不同株型玉米花期在水分胁迫后, 其穗位叶的上/下表皮气孔个数显著增多, 而上/下表皮气孔长度和宽度及气孔导度均显著降低。这与于海秋等[34]和李真真等[35]的研究结果类似。其可能是缺水时叶片生长受到抑制, 叶面积变小, 进而影响了单位面积上气孔的排列、大小及开度有一定的关系。另外, 本研究还发现紧凑株型廊黄、昌7-2穗位叶的上/下表皮气孔长度和宽度及气孔导度的降低幅度远大于平展株型TS141, 而廊黄、昌7-2穗位叶的上/下表皮气孔个数上升幅度远小于TS141, 阐明紧凑株型玉米气孔受干旱胁迫的影响较平展株型玉米更小。
玉米遭受干旱等逆境胁迫时其各种生理代谢反应发生紊乱, 进而影响玉米的正常生长发育。叶绿素a和b是光合作用中光能的主要载体, 其含量会引起一系列的生理代谢变化, 可在一定程度上反映植株的光合性能[36]。本研究表明花期3份不同株型玉米在干旱胁迫下穗位叶叶绿素a、b及(a+b)含量均降低。这与杨碧云等[37]对干旱胁迫下紫色小白菜和Rajasekar等[38]对玉米所做的研究结果相一致。其可能原因是干旱胁迫导致原卟啉IX氧化酶和原叶绿素酸酯氧化还原酶等叶绿素合成酶活性及叶绿素合成速率降低, 或叶绿素b还原酶、脱镁螯合酶、脱镁叶绿酸a氧化酶等叶绿素降解酶活性及叶绿素降解速率加剧有关。叶绿素a/b反映类囊体的垛叠程度, 类囊体的垛叠程度越小, 光抑制越强[21]。本研究发现干旱胁迫下平展株型TS141的叶绿素a/b显著降低, 而另2份紧凑株型玉米的叶绿素a/b显著升高, 意味着干旱胁迫下紧凑株型玉米较平展株型玉米其类囊体的垛叠程度增大, 受光抑制程度减弱。
光合作用是植物最基本的生命活动对干旱胁迫响应极为敏感, 当植物遭受缺水时光合速率会不同程度的降低。徐芬芬等[23]研究表明紫色小白菜随着NaCl胁迫的增大, 净光合速率、胞间CO2浓度、RuBPCase活性及光系统PSⅡ 活性均显著降低。Hazem等[39]研究表明叙利亚大麦(Hordeum vulgare)的光合速率、光系统PSⅡ 活性及最大光化学效率(Fv/Fm)与其生长状态呈显著正相关, 当叙利亚大麦处于120 mmol· L-1 NaCl逆境压力时其光系统PSⅡ 明显受到破坏, 光合速率和Fv/Fm降低。本研究表明花期干旱胁迫下3份不同株型玉米穗位叶的净光合速率、蒸腾速率及RuBPCase活性均降低, 而其胞间CO2浓度均升高, 且紧凑株型廊黄和昌7-2穗位叶的净光合速率、胞间CO2浓度、蒸腾速率及RuBPCase活性的变化幅度较平展株型玉米TS141更小, 这与何海军等[15]的研究结果类似, 何海军等[15]指出干旱胁迫条件下, 平展株型玉米品种豫玉22、中间株型品种沈单16及紧凑株型品种先玉335的净光合速率、蒸腾速率、胞间CO2浓度均随干旱胁迫程度加大而显著减少, 且这3个光合特性的大小均表现为先玉335> 沈单16> 豫玉22。其原因可能与干旱胁迫下不同株型玉米叶片叶绿素合成、光系统活性、气孔发育和开度及叶肉细胞活性受到抑制等因素有关。另外, Toshiyuki等[40]研究报道叶片温度的变化可以作为干旱胁迫下衡量作物生长发育状况的一个重要指标。本研究分析也表明, 3份不同株型玉米穗位叶的叶片温度在干旱胁迫下降低, 且平展株型TS141的叶片温度降低程度大于紧凑株型廊黄和昌7-2, 这可能与干旱胁迫下平展株型玉米的株型结构变化较大有关, 也可能与其气孔大小、开度及蒸腾速率等有关, 进而影响到不同株型玉米植株的水分状况, 还需有待进一步研究。
本研究表明, 由于紧凑株型玉米具有更合理的株型结构, 进而紧凑株型玉米能够更少的接收太阳辐射、维持更低的叶片温度和蒸腾散失、能更高效地提高水分利用效率及能更好地改善植株生理代谢, 因此, 紧凑株型玉米较平展株型玉米更耐旱。但是当干旱程度超过紧凑株型玉米植株自身的调节能力时, 紧凑株型玉米的生长和代谢就会受到显著抑制并影响到玉米的产量。基于以上考虑, 在一定的干旱环境下, 今后可建立玉米株型相关耐旱体系, 通过育种手段来遗传改良玉米株型结构, 使其成为植株气孔和生理代谢的调节者, 同时又能让玉米保持较高的光合速率及水分利用效率, 培育出适宜旱区推广的优良新品种。此外, 在优良新品种培育中要兼顾各株型性状所起的作用, 尽可能地使玉米株型结构合理, 进而最大限度地发挥品种自身的增产潜力。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|